Protective Role of α2HS-Glycoprotein in HBV-Associated Liver Failure

Abstract
:1. Introduction
2. Results
2.1. Subjects
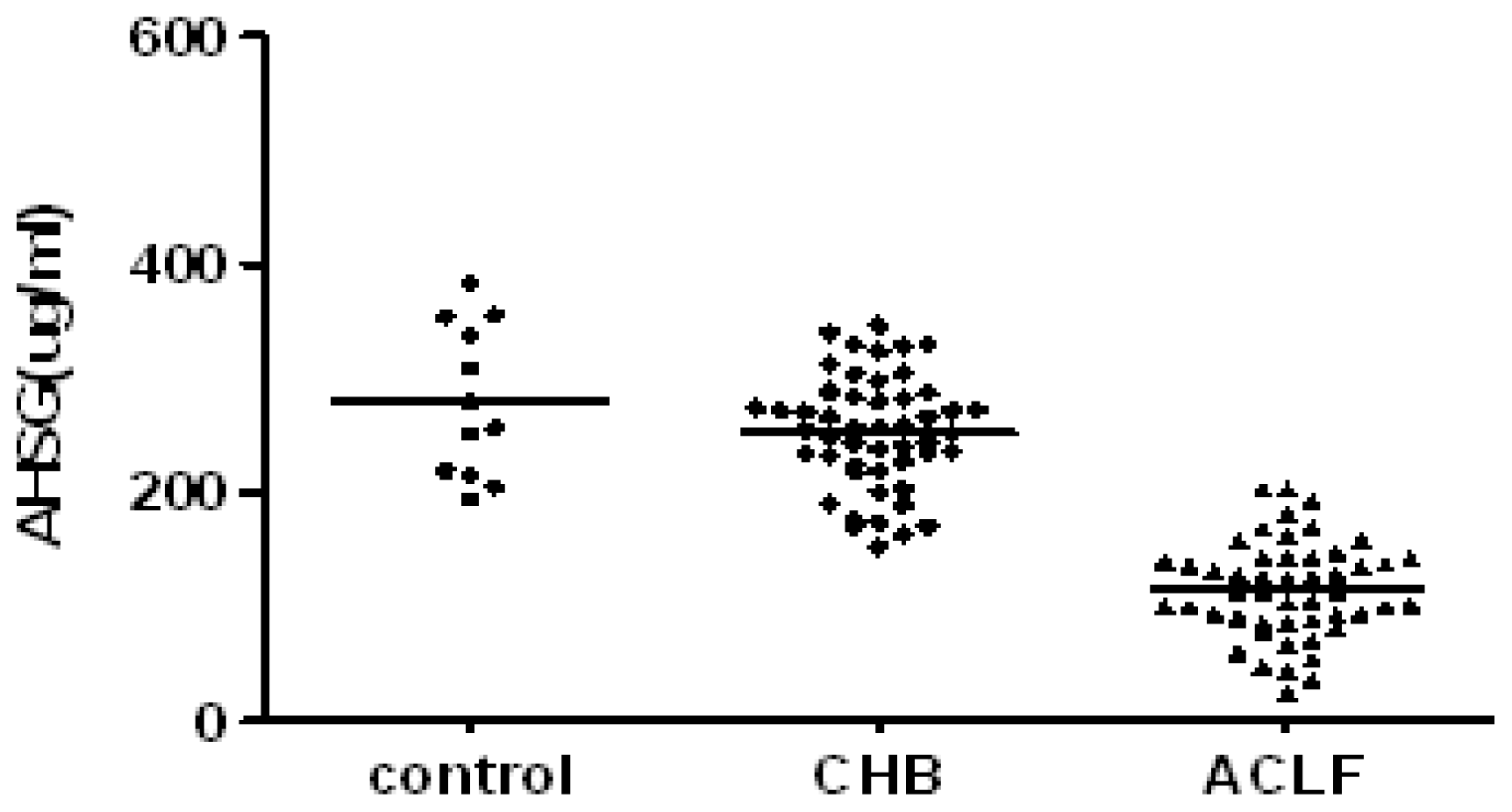
2.2. Serum AHSG and TNF-α in Subject with CHB and ACLF
2.3. Serum AHSG Levels and Short-Term Mortality
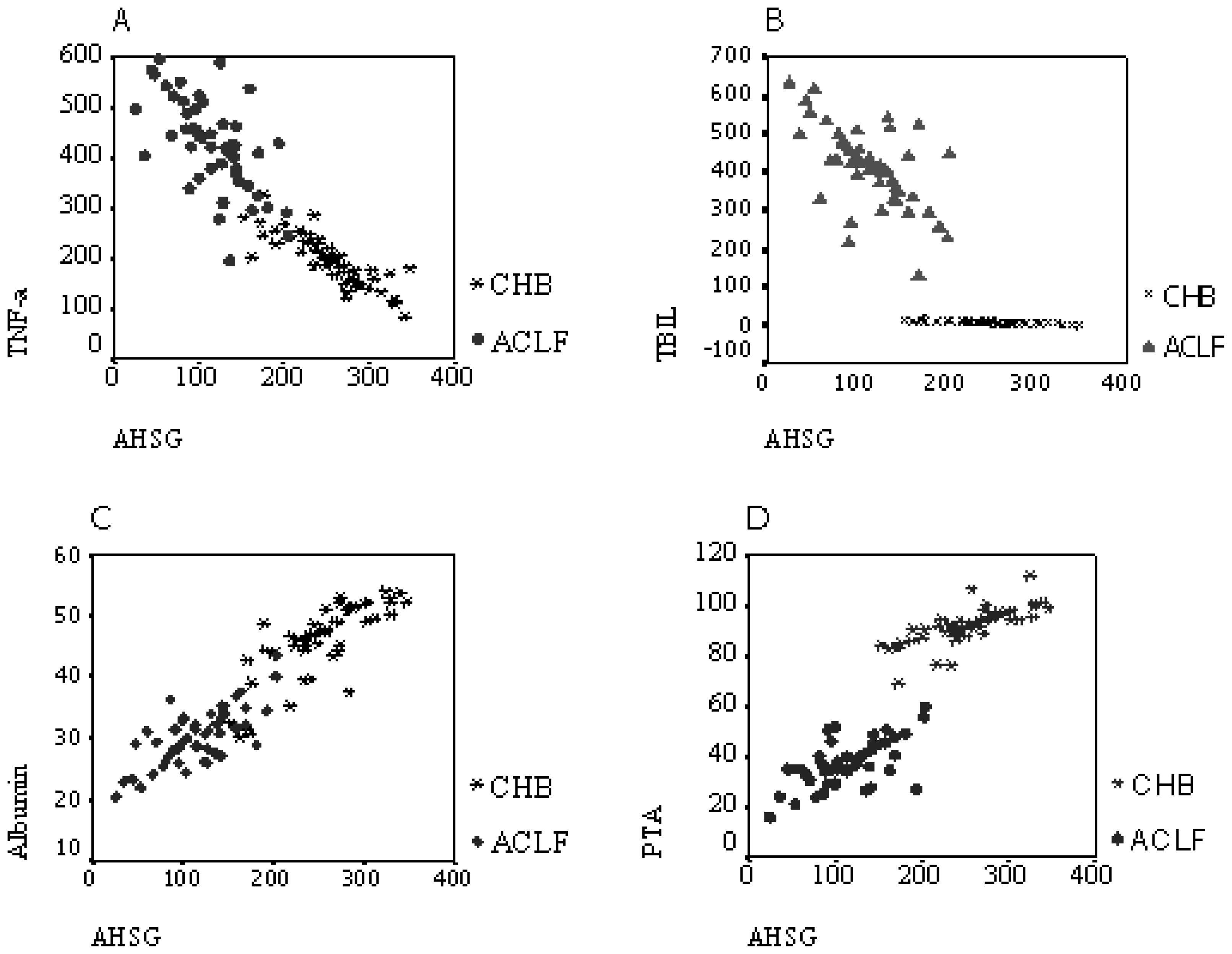
2.4. Correlation Analysis
2.5. Effects of AHSG on Cytokine Production in LPS-Stimulated PBMCs
3. Discussion
4. Materials and Methods
4.1. Determination of Serum AHSG and TNF-α Concentration
4.2. Preparation and Culture of PBMCs
4.3. Cell Culture and Cytokine ELISA
4.4. Statistical Analysis
5. Conclusions
Acknowledgments
References
- Dziegielewska, KM; Møllgård, K; Reynolds, ML; Saunders, NR. A fetuin-related glycoprotein (alpha 2HS) in human embryonic and fetal development. Cell Tissue Res 1987, 248, 33–41. [Google Scholar]
- Triffitt, JT; Gebauer, U; Ashton, BA; Owen, ME; Reynolds, JJ. Origin of plasma alpha2HS-glycoprotein and its accumulation in bone. Nature 1976, 262, 226–236. [Google Scholar]
- Ohnishi, T; Nakamura, O; Arakaki, N; Daikuhara, Y. Effect of phosphorylated rat fetuin on the growth of hepatocytes in primary culture in the presence of human hepatocyte-growth factor. Evidence that phosphorylated fetuin is a natural modulator of hepatocyte-growth factor. Eur. J. Biochem 1997, 243, 753–761. [Google Scholar]
- Szweras, M; Liu, D; Partridge, EA; Pawling, J; Sukhu, B; Clokie, C; Jahnen, DW; Tenenbaum, HC; Swallow, CJ; Grynpas, MD; Dennis, JW. Alpha 2-HS glycoprotein/fetuin, a transforming growth factor-beta/bone morphogenetic protein antagonist, regulates postnatal bone growth and remodeling. J. Biol. Chem 2002, 277, 19991–19997. [Google Scholar]
- Komaba, H; Fukagawa, M. Fetuin-mineral complex: A new potential biomarker for vascular calcification? Kidney Int 2009, 75, 874–876. [Google Scholar]
- Wang, HC; Li, W; Zhu, S; Li, J; Amore, J; Ward, MF; Yang, H; Wu, R; Jahnen, DW; Tracey, KJ; Wang, P; Sama, AE. Peripheral administration of fetuin-A attenuates early cerebral ischemic injury in rats. J. Cerebr. Blood Flow Metab 2010, 30, 493–504. [Google Scholar]
- Ombrellino, M; Wang, H; Yang, H; Zhang, M; Vishnubhakat, J; Frazier, A; Scher, LA; Friedman, SG; Tracey, KJ. Fetuin, a negative acute phase protein, attenuates TNF synthesis and the innate inflammatory response to carrageenan. Shock 2001, 15, 181–185. [Google Scholar]
- Zhang, M; Caragine, T; Wang, H; Cohen, PS; Botchkina, G; Soda, K; Bianchi, M; Ulrich, P; Cerami, A; Sherry, B; Tracey, KJ. Spermine inhibits proinflammatory cytokine synthesis in human mononuclear cells: A counterregulatory mechanism that restrains the immune response. J. Exp. Med 1997, 185, 1759–1768. [Google Scholar]
- Wang, H; Li, W; Zhu, S; Li, J. Fetuin protects mice against lethal sepsis by modulating bacterial endotoxin-induced hmgb1 release and autophagy. Shock 2010, 33, 6–7. [Google Scholar]
- Wang, H; Zhang, MH; Kuniyasu, S; Andrew, S; Kevin, JT. Fetuin protects the fetus from TNF. Lancet 1997, 350, 861–862. [Google Scholar]
- Liu, Q. Role of cytokines in the pathophysiology of acute-on-chronic liver failure. Blood Purif 2009, 28, 331–341. [Google Scholar]
- Koziel, MJ. The immunopathogenesis of HBV infection. Antivir. Ther 1998, 3, 13–24. [Google Scholar]
- Chisari, FV; Ferrari, C. Hepatitis B virus immunopathogenesis. Annu. Rev. Immunol 1995, 13, 29–60. [Google Scholar]
- Zhang, JY; Zhang, Z; Lin, F; Zou, ZS; Xu, RN; Jin, L; Fu, JL; Shi, F; Shi, M; Wang, HF; Wang, FS. Interleukin-17-producing CD4(+) T cells increase with severity of liver damage in patients with chronic hepatitis B. Hepatology 2010, 51, 81–91. [Google Scholar]
- Sarin, SK; Kumar, A; Almeida, JA; Chawla, YK; Fan, ST; Garg, H; Silva, HJ; Hamid, SS; Jalan, R; Komolmit, P; et al. Acute-on-chronic liver failure: Consensus recommendations of the Asian Pacific Association for the study of the liver (APASL). Hepatol. Int 2009, 3, 269–282. [Google Scholar]
- Papatheodoridis, GV; Manolakopoulos, S. EASL clinical practice guidelines on the management of chronic hepatitis B: The need for liver biopsy. J. Hepatol 2009, 51, 226–237. [Google Scholar]
- Ix, JH; Sharma, K. Mechanisms linking obesity, chronic kidney disease, and fatty liver disease: The roles of fetuin-A, adiponectin, and AMPK. J. Am. Soc. Nephrol 2010, 21, 406–412. [Google Scholar]
- Sari, I; Kebapcilar, L; Taylan, A; Bilgir, O; Kozaci, DL; Yildiz, Y; Yuksel, A. Fetuin-A and interleukin-18 levels in ankylosing spondylitis. Int. J. Rheum. Dis 2010, 13, 75–81. [Google Scholar]
- Kalabay, L; Jakab, L; Prohászka, Z; Füst, G; Benkö, Z; Telegdy, L; Lörincz, Z; Závodszky, P; Arnaud, P; Fekete, B. Human fetuin/alpha 2HS-glycoprotein level as a novel indicator of liver cell function and short-term mortality in patients with liver cirrhosis and liver cancer. Eur. J. Gastroenterol. Hepatol 2002, 14, 389–394. [Google Scholar]
- Kalabay, L; Gráf, L; Vörös, K; Jakab, L; Benkő, Z; Telegdy, L; Fekete, B; Prohászka, Z; George, F. Human serum fetuin A/alpha 2HS-glycoprotein level is associated with long-term survival in patients with alcoholic liver cirrhosis, comparison with the Child-Pugh and MELD scores. BMC Gastroenterol 2007, 6, 7–14. [Google Scholar]
- Yilmaz, Y; Yonal, O; Kurt, R; Ari, F; Oral, AY; Celikel, CA; Korkmaz, S; Ulukaya, E; Ozdogan, O; Imeryuz, N; Avsar, E; Kalayci, C. Serum fetuin A/alpha2HS-glycoprotein levels in patients with non-alcoholic fatty liver disease: Relation with liver fibrosis. Ann. Clin. Biochem 2010, 47, 549–553. [Google Scholar]
- Liu, Q. Role of cytokines in the pathophysiology of acute-on-chronic liver failure. Blood Purif 2009, 28, 331–341. [Google Scholar]
- Yuan, GJ; Gong, ZJ; Zhou, XR; Zhang, QP; Sun, XM; Li, X. Epigallocatechin-3-gallate ameliorates alcohol-induced liver injury in rats. Int. J. Mol. Sci 2006, 7, 204–219. [Google Scholar]
- Liguori, MJ; Ditewig, AC; Maddox, JF; Luyendyk, JP; Lehman-McKeeman, LD; Nelson, M; Bhaskaran, VM; Waring, JF; Ganey, PE; Roth, RA; Blomme, EA. Comparison of TNF alpha to lipopolysaccharide as an inflammagen to characterize the idiosyncratic hepatotoxicity potential of drugs: Trovafloxacin as an example. Int. J. Mol. Sci 2010, 11, 4697–4714. [Google Scholar]
- Sen, S; Davies, NA; Mookerjee, RP; Cheshire, LM; Hodges, SJ; Williams, R; Jalan, R. Pathophysiological effects of albumin dialysis in acute-on-chronic liver failure: A randomized controlled study. Liver Transpl 2004, 10, 1109–1119. [Google Scholar]
- Ambrosino, G; Naso, A; Feltracco, P; Carraro, P; Basso, SM; Varotto, S; Cillo, U; Zanus, G; Boccagni, P; Brolese, A; Plebani, M; Giron, G; Amico, DF. Cytokines and liver failure: Modification of TNF- and IL-6 in patients with acute on chronic liver decompensation treated with Molecular Adsorbent Recycling System (MARS). Acta Biomed 2003, 74, 7–9. [Google Scholar]
- Wang, JH; Redmond, HP; Watson, RW; Bouchier, HD. Role of lipopolysaccharide and tumor necrosis factor-alpha in induction of hepatocyte necrosis. Am. J. Physiol 1995, 269, 297–304. [Google Scholar]
- Shah, VO; Ferguson, JE; Hunsaker, LA; Deck, LM; Vander, DL. Natural products inhibit LPS-induced activation of pro-inflammatory cytokines in peripheral blood mononuclear cells. Nat. Prod. Res 2010, 24, 1177–1188. [Google Scholar]
- Li, W; Zhu, S; Li, J; Huang, Y; Zhou, R; Fan, X; Yang, H; Gong, X; Eissa, NT; Jahnen, DW; Wang, P; Tracey, KJ; Sama, AE; Wang, H. A hepatic protein, fetuin-A, occupies a protective role in lethal systemic inflammation. Plos One 2011, 6, e16945. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Controls | CHB | ACLF | |
|---|---|---|---|
| Sex (M/F) | 12 (8/4) | 50 (37/13) | 50 (38/12) |
| Average Age (yr) | 38.3 ± 9.5 | 40.3 ± 5.8 | 39.0 ± 6.0 |
| Average ALT (U/L) | 21.96 ± 4.72 | 55.48 ± 10.60 | 449.21 ± 57.40 |
| Average TBIL (μmol/L) | 8.42 ± 1.40 | 9.81 ± 3.64 | 414.08 ± 102.63 |
| Average A (g/L) | 44.65 ± 5.39 | 46.03 ± 6.18 | 30.28 ± 4.65 |
| PTA (%) | 93.34 ± 2.13 | 92.00 ± 7.39 | 37.60 ± 9.13 |
| Serum HBV-DNA (+) | 0 | 30 | 50 |
| Groups | n | AHSG (μg/mL) | PTA (%) | TBIL (μmol/L) | A (g/L) | TNF-α (pg/mL) |
|---|---|---|---|---|---|---|
| A | 15 | 85.26 ± 20.54# | 29.9 ± 8.22* | 453.09 ± 117.98 | 28.53 ± 4.85 | 448.27 ± 100.58 |
| B | 35 | 128.05 ± 36.52 | 40.90 ± 7.26 | 397.36 ± 92.13 | 31.03 ± 4.43 | 417.47 ± 87.03 |
| Group | Supernatant TNF-α levels (pg/mL) | Supernatant IL-6 levels (pg/mL) | |
|---|---|---|---|
| 1 | PBMC | 6.44 ± 3.88 | 19.21 ± 5.59 |
| 2 | PBMC + LPS | 423.73 ± 97.66* | 584.73 ± 82.36& |
| 3 | PBMC + LPS + spermine | 72.97 ± 18.32** | 239.96 ± 48.26** |
| 4 | PBMC + LPS + spermine + AHSG | 25.11 ± 5.61*** | 114.90 ± 30.83*** |
| 5 | PBMC + LPS + spermine + AHSG antibody | 261.43 ± 54.07# | 555.42 ± 69.78@ |
| 6 | PBMC + LPS + albumin | 223.92 ± 46.65## | 403.46 ± 78.80## |
| 7 | PBMC + LPS + AHSG | 258.55 ± 54.87### | 503.78 ± 90.87### |
© 2011 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Dai, X.-H.; Zhang, P.; Xiao, M.-F.; Zhou, R.-R.; Zhang, B.-X.; Hu, G.-S.; Huang, Z.-B.; Fan, X.-G. Protective Role of α2HS-Glycoprotein in HBV-Associated Liver Failure. Int. J. Mol. Sci. 2011, 12, 3846-3856. https://doi.org/10.3390/ijms12063846
Dai X-H, Zhang P, Xiao M-F, Zhou R-R, Zhang B-X, Hu G-S, Huang Z-B, Fan X-G. Protective Role of α2HS-Glycoprotein in HBV-Associated Liver Failure. International Journal of Molecular Sciences. 2011; 12(6):3846-3856. https://doi.org/10.3390/ijms12063846
Chicago/Turabian StyleDai, Xia-Hong, Pan Zhang, Mei-Fang Xiao, Rong-Rong Zhou, Bao-Xin Zhang, Guan-Sheng Hu, Ze-Bing Huang, and Xue-Gong Fan. 2011. "Protective Role of α2HS-Glycoprotein in HBV-Associated Liver Failure" International Journal of Molecular Sciences 12, no. 6: 3846-3856. https://doi.org/10.3390/ijms12063846