Modulation Role of Abscisic Acid (ABA) on Growth, Water Relations and Glycinebetaine Metabolism in Two Maize (Zea mays L.) Cultivars under Drought Stress

Abstract
:1. Introduction
2. Results and Discussion
2.1. Modulation Role of Abscisic Acid (ABA) on the Growth and Leaf Water Relations of Maize Seedlings under Drought Stress
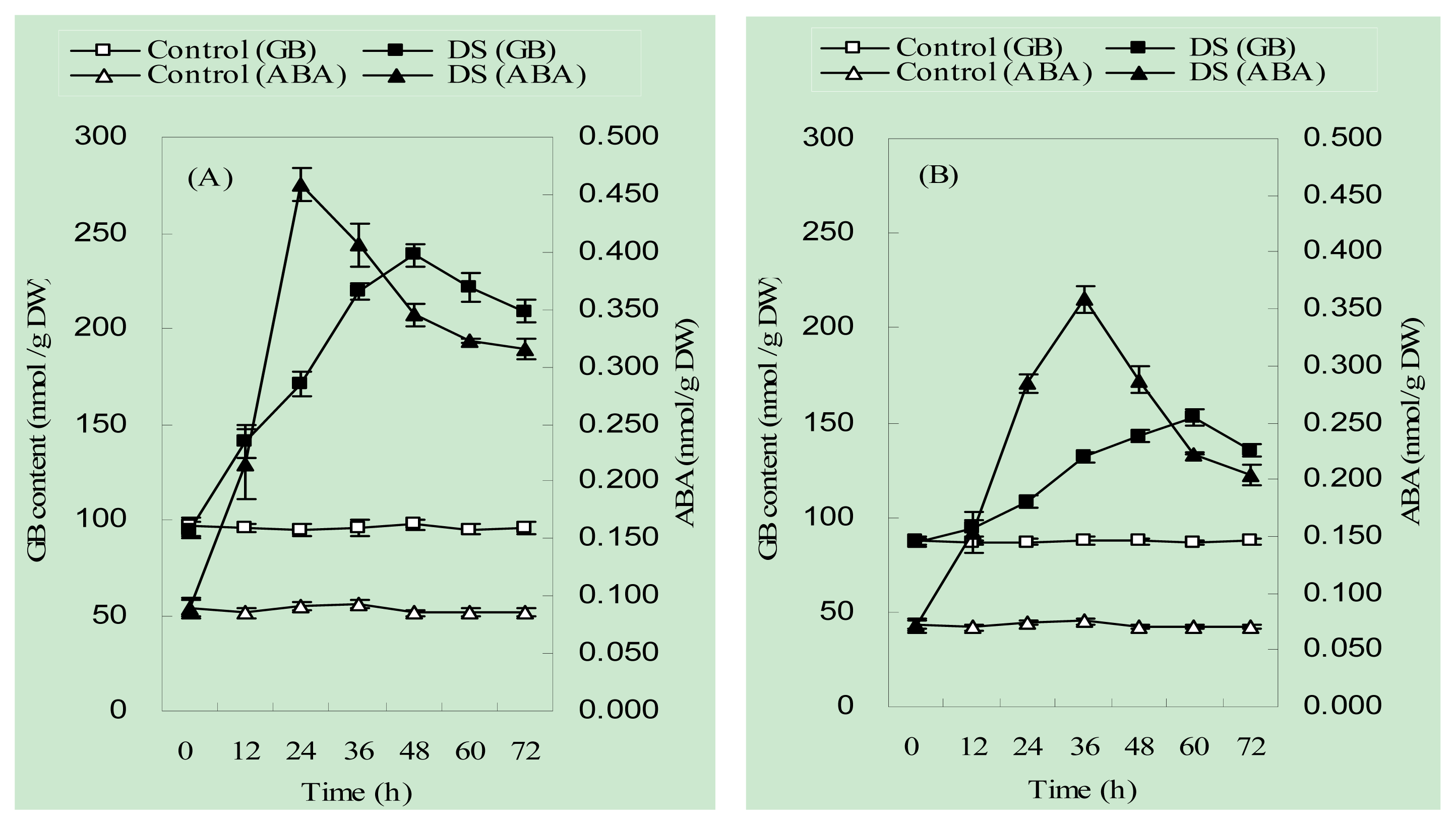
2.2. The Accumulation Pattern of Endogenous ABA and Glycinebetaine (GB) in Maize Seedlings under Drought Stress
2.3. Modulation Role of ABA on Endogenous ABA and Glycinebetaine (GB) Accumulation in the Leaves of Maize Plants under Drought Stress
2.4. Modulation Role of ABA on GB Metabolism in the Leaves of Maize Plants under Drought Stress
2.5. Correlations among All Parameters Measured
2.6. Interaction of Exogenous ABA or Flu Treatment and Water Regimes as well as Correlation Coefficients for All Parameters Measured
2.7. General Discussion
3. Materials and Methods
3.1. Plant Material and Trial Location
3.2. Plant Growth and Experimental Design
3.3. Sampling and Recording of Data
3.4. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Li, S.X. Dry Land Agriculture in China; China Science Press: Beijing, China, 2007; pp. 1–5. [Google Scholar]
- Zhang, L.X.; Li, S.X.; Zhang, H.; Liang, Z.S. Nitrogen rates and water stress effects on production, lipid peroxidation and antioxidative enzyme activities in two maize (Zea mays L.) genotypes. J. Agron. Crop Sci 2007, 193, 387–397. [Google Scholar]
- Lu, G.H.; Ren, D.L.; Wang, X.Q.; Wu, J.K.; Zhao, M.S. Evaluation on drought tolerance of maize hybrids in China. J. Maize Sci 2010, 3, 20–24. [Google Scholar]
- Ashraf, M. Inducing drought tolerance in plants: Some recent advances. Biotechnol. Adv 2010, 28, 169–183. [Google Scholar]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci 2002, 7, 405–410. [Google Scholar]
- Zhang, L.X.; Li, S.X.; Liang, Z.S.; Li, S.Q. Effect of foliar nitrogen application on nitrogen metabolism, water status and plant growth in two maize (Zea mays L.) cultivars under short-term moderate stress. J. Plant Nutr 2009, 32, 1–21. [Google Scholar]
- Ashraf, M.; Foolad, M.R. Roles of glycinebetaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot 2007, 59, 206–216. [Google Scholar]
- Zhang, L.X.; Li, S.X.; Liang, Z.S. Differential plant growth and osmotic effects of two maize (Zea mays L.) cultivars to exogenous glycinebetaine application under drought stress. Plant Growth Regul 2009, 58, 297–305. [Google Scholar]
- Rhodes, D.; Hanson, A.D. Quaternary ammonium and tertiary sulfonium compounds in higher-plants. Annu. Rev. Plant Physiol. Plant Mol. Biol 1993, 44, 357–384. [Google Scholar]
- Sakamoto, A.; Murata, N. The role of glycine betaine in the protection of plants from stress: Clues from transgenic plants. Plant Cell Environ 2002, 25, 163–171. [Google Scholar]
- Sithtisarn, S.; Harinasut, P.; Pornbunlualap, S.; Cha-Um, S. Accumulation of glycinebetaine and betaine aldehyde dehydrogenase activity in Eucalyptus camaldulensis clone T5 under in vitro salt stress. Kasetsart J. (Nat. Sci.) 2009, 43, 146–152. [Google Scholar]
- Jakab, G.; Ton, J.; Flors, V.; Zimmerli, L.; Métraux, J.; Mauch-Mani, B. Enhancing Arabidopsis salt and drought stress tolerance by chemical priming for its abscisic acid responses. Plant Physiol 2005, 139, 267–274. [Google Scholar]
- Jiang, W.; Lafitte, R. Ascertain the effect of PEG and exogenous ABA on rice growth at germination stage and their contribution to selecting drought tolerant genotypes. Asian J. Plant Sci 2007, 6, 684–687. [Google Scholar]
- Maleki, M.; Ebrahimzade, H.; Gholami, M.; Niknam, V. The effect of drought stress and exogenous abscisic acid on growth, protein content and antioxidative enzyme activity in saffron (Crocus sativus L.). Afr. J. Biotechnol 2011, 45, 9068–9075. [Google Scholar]
- Kurahashi, Y.; Terashima, A.; Takumi, S. Variation in dehydration tolerance, ABA sensitivity and related gene expression patterns in D-Genome progenitor and synthetic hexaploid wheat lines. Int. J. Mol. Sci 2009, 10, 2733–2751. [Google Scholar]
- Hooker, S.T.; Thorpe, A.T. Effects of fluridone and abscisic acid on lateral root initiation and root elongation of excised tomato roots cultured in vitro. Plant Cell Tissue Org 1998, 52, 199–203. [Google Scholar]
- Ruggiero, B.; Koiwa, H.; Manabe, Y.; Quist, T.M.; Inan, G.; Saccardo, F.; Joly, R.J.; Hasegawa, P.M.; Bressan, R.A.; Maggio, A. Uncoupling the effects of abscisic acid on plant growth and water relations. Analysis of sto1/nced3, an abscisic acid-deficient but salt stress tolerant mutant in Arabidopsis. Plant Physiol 2004, 136, 3134–3147. [Google Scholar]
- Jiang, F.; Hartung, W. Long-distance signalling of abscisic acid (ABA): The factors regulating the intensity of the ABA signal. J. Exp. Bot 2007, 59, 37–43. [Google Scholar]
- Achard, P.; Cheng, H.; de Grauwe, L.; Decat, J.; Schoutteten, H.; Moritzn, T.; van Der Straeten, D.; Peng, J.R.; Harberd, N.P. Integration of plant responses to environmentally activated phytohormonal signals. Science 2006, 311, 91–94. [Google Scholar]
- Maggio, A.; Barbieri, G.; Raimondi, G.; Pascale, S.D. Contrasting effects of GA3 treatments on tomato plants exposed to increasing salinity. J. Plant Growth Regul 2010, 29, 63–72. [Google Scholar]
- Zeevart, J.A.D.; Creelman, R.A. Metabolism and physiology of abscisic acid. Annu. Rev. Plant Physiol 1998, 39, 439–473. [Google Scholar]
- Ren, H.; Zhihui, G.; Lin, C.; Kaifa, W.; Jing, L.; Yijuan, F.; William, J.D.; Wensuo, J.; Jianhua, Z. Dynamic analysis of ABA accumulation in relation to the rate of ABA catabolism in maize tissues under water deficit. J. Exp. Bot 2007, 58, 211–219. [Google Scholar]
- Zhang, J.H.; Jia, W.S.; Yang, J.C.; Ismail, A.M. Role of ABA in integrating plant responses to drought and salt stresses. Field Crops Res 2006, 97, 111–119. [Google Scholar]
- Hancock, J.T.; Neill, S.J.; Wilson, I.D. Nitric oxide and ABA in the control of plant function. Plant Sci 2011, 181, 555–559. [Google Scholar]
- Zhang, L.X.; Gao, M.; Li, S.Q.; Li, S.X.; Liang, Z.S. Water status and photosynthesis in two maize (Zea Mays L.) cultivars as affected by supplied nitrogen form and drought stress. Pak. J. Bot 2011, 43, 1995–2001. [Google Scholar]
- Zhang, Y.K.; Wang, L.X.; Yang, J.H.; Liang, D.J.; Wang, X.L.; Xi, L.Y. China maize potential yield developing technique advanced. Chin. Agri. Sci. Bull 2007, 7, 267–269. [Google Scholar]
- Hattori, T.; Mitsuya, S.; Fujiwara, T.; Jagendorf, A.T.; Takabe, T. Tissue specificity of glycinebetaine synthesis in barley. Plant Sci 2009, 176, 112–118. [Google Scholar]
- Hanson, A.D.; Wyse, R. Biosynthesis, translocation, and accumulation of betaine in sugar beet and its progenitors in relation to salinity. Plant Physiol 1982, 70, 1191–1198. [Google Scholar]
- Ashraf, M.; Harris, P.J.C. Potential biochemical indicators of salinity tolerance in plants. Plant Sci 2004, 166, 3–16. [Google Scholar]
- Taiz, L.; Zeiger, E. Plant Physiology, 3rd ed; Sinauer Associates, Inc: Sunderland, MA, USA, 2002; pp. 591–620. [Google Scholar]
- Ishitani, M.; Nakamura, T.; Han, S.Y.; Takabe, T. Expression of the betaine aldehyde dehydrogenase gene in barley in response to osmotic stress and abscisic acid. Plant Mol. Biol 1995, 27, 307–315. [Google Scholar]
- Gao, X.P.; Pan, Q.H.; Li, M.J.; Zhang, L.Y.; Wang, X.F.; Shen, Y.Y.; Lu, Y.F.; Chen, S.W.; Liang, Z.; Zhang, D.P. Abscisic acid is involved in the water stress-induced betaine accumulation in pear leaves. Plant Cell Physiol 2004, 45, 742–750. [Google Scholar]
- Saneoka, H.; Ishiguro, S.; Moghaieb, R.E.A. Effect of salinity and abscisic acid on accumulation of glycinebetaine and betaine aldehyde dehydrogenase mRNA in Sorghum leaves (Sorghum bicolor). J. Plant Physiol 2001, 158, 853–859. [Google Scholar]
- Maldonado, C.A.; Zuniga, G.E.; Corcuera, L.J.; Alberdi, M. Effect of water stress on frost resistance of oat leaves. Environ. Exp. Bot 1997, 38, 99–107. [Google Scholar]
- Rai, S.P.; Luthra, R.; Gupta, M.M.; Kumar, S. Pleiotropic morphological and abiotic stress resistance phenotypes of the hyper-abscisic acid producing Abo-mutant in the periwinkle Catharanthus roseus. J. Biosci 2001, 26, 57–70. [Google Scholar]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Gene expression and signal transduction in water-stress response. Plant Physiol 1997, 115, 327–334. [Google Scholar]
- Hoagland, D.R.; Arnon, D.I. The water culture method for growing plants without soils. Calif. Agric. Exp. Sta. Cir 1950, 347, 1–32. [Google Scholar]
- Wang, J.; Li, D.Q. Effects of water stress on AsA-GSH cycle and H2O2 content in maize root. Chin. J. Eco-Agric 2002, 2, 94–96. [Google Scholar]
- Gao, J.F. Experiment Technique of Plant Physiology; Xi’an World Books Press Company: Xi’an, China, 2000; pp. 23–25. [Google Scholar]
- Grieve, C.M.; Grattan, S.R. Rapid assay for determination of water soluble quaternary ammonium compounds. Plant Soil 1983, 70, 303–307. [Google Scholar]
- Daniell, H.; Muthukumar, B.; Lee, S.B. Marker free transgenic plants: Engineering the chloroplast genome without the use of antibiotic selection. Curr. Genet 2001, 39, 109–116. [Google Scholar]
- Feng, Z.X.; Ren, A.N. Mensuration of the content of choline in diospyros leaves. Tianjin J. Tradit. Chin. Med 2004, 21, 248–249. [Google Scholar]
- Richard, J.W.; Emily, R.M. Spectrophotometric determination of small amounts of choline. J. Biol. Chem 1945, 159, 395–397. [Google Scholar]
- Weiler, E. An enzyme-immunoassay for cis-(+)-abscisic acid. Physiol. Plant 1982, 54, 510–514. [Google Scholar]
- SAS Software, version 8.2; SAS Institute, Inc: Cary, NC, USA, 1996.



{kind=link}
{kind=link}
| Treatment | DM (g/plant) | RWC (%) | ||
|---|---|---|---|---|
| Zhengdan 958 | Jundan 20 | Zhengdan 958 | Jundan 20 | |
| Control | 0.362 ± 0.011 a | 0.397 ± 0.013 a | 91.78 ± 1.20 a | 92.11 ± 1.12 a |
| (B) | (A) | (A) | (A) | |
| PEG | 0.268 ± 0.012 b | 0.239 ± 0.015 b | 82.83 ± 1.15 b | 79.01 ± 1.15 b |
| (A) | (B) | (A) | (B) | |
| Control + ABA | 0.347 ± 0.021 a | 0.394 ± 0.010 a | 90.17 ± 1.46 a | 91.09 ± 1.25 a |
| (B) | (A) | (A) | (A) | |
| PEG + ABA | 0.311 ± 0.007 c | 0.288 ± 0.009 c | 86.77 ± 1.50 c | 83.02 ± 1.07 c |
| (A) | (B) | (A) | (B) | |
| Control + Flu | 0.338 ± 0.011 ab | 0.381 ± 0.014 a | 91.12 ± 1.65 a | 92.32 ± 1.55 a |
| (A) | (A) | (A) | (A) | |
| PEG + Flu | 0.225 ± 0.008 d | 0.204 ± 0.011 d | 79.22 ± 1.75 d | 76.02 ± 1.60 d |
| (A) | (B) | (A) | (B) | |
| Parameter | DM | RWC | ABAC | GBC | CC | BADHA |
|---|---|---|---|---|---|---|
| DM | 0.946 *** | 0.930 *** | 0.941 *** | 0.684 ** | 0.943 *** | |
| RWC | 0.356 | 0.912 *** | 0.988 *** | 0.789 *** | 0.974 *** | |
| ABAC | 0.422 | 0.122 | 0.941 *** | 0.501 * | 0.888 *** | |
| GBC | 0.567 * | 0.516 * | 0.375 | 0.915 *** | 0.978 *** | |
| CC | 0.448 | 0.422 | 0.374 | 0.739 *** | 0.808 *** | |
| BADHA | 0.208 | 0.342 | 0.251 | 0.705 ** | 0.611 ** |
| Variation | W | Cv | A | W × Cv | W × A | Cv × A | W × Cv × A |
|---|---|---|---|---|---|---|---|
| DM | 1133.5 *** | 5.8 * | 42.6 *** | 128.5 *** | 86.6 *** | 8.9 * | 8.5 * |
| RWC | 3129.9 *** | 496.2 *** | 532.6 *** | 205.5 *** | 619.7 *** | 9.3 ** | 9.7 ** |
| GBC | 3406.9 *** | 261.2 *** | 276.8 *** | 217.7 *** | 290.6 *** | 11.7 ** | 12.9 ** |
| ABAC | 15932.8 *** | 140.7 *** | 829.1 *** | 83.6 *** | 220.5 *** | 10.07 ** | 10.07 ** |
| CC | 3982.9 *** | 258.3 *** | 10.9 ** | 215.1 *** | 34.4 *** | 0.03 | 0.01 |
| BADHA | 2344.0 *** | 48.5 *** | 53.1 *** | 17.5 *** | 67.4 *** | 3.4 | 0.34 |
| W | Cv | Flu | W × Cv | W × Flu | Cv × Flu | W × Cv × Flu | |
| DM | 613.7 *** | 5.83 * | 97.4 *** | 9.3 ** | 12.2 ** | 8.0 * | 12.0 ** |
| RWC | 6798.1 *** | 308.4 *** | 381.6 *** | 159.7 *** | 74.3 *** | 56.5 *** | 10.6 ** |
| GBC | 5641.3 *** | 264.6 *** | 283.0 *** | 246.4 *** | 212.8 *** | 8.3 * | 9.9 ** |
| ABAC | 12090.5 *** | 86.4 *** | 1390.9 *** | 80.2 *** | 868.6 *** | 27.2 *** | 8.0 * |
| CC | 1751.4 *** | 70.7 *** | 4.17 * | 80.2 *** | 13.7 ** | 10.5 ** | 2.6 |
| BADHA | 2611.6 *** | 137.0 *** | 244.2 *** | 5.6 * | 5.1 * | 35.6 ** | 4.1 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, L.; Gao, M.; Hu, J.; Zhang, X.; Wang, K.; Ashraf, M. Modulation Role of Abscisic Acid (ABA) on Growth, Water Relations and Glycinebetaine Metabolism in Two Maize (Zea mays L.) Cultivars under Drought Stress. Int. J. Mol. Sci. 2012, 13, 3189-3202. https://doi.org/10.3390/ijms13033189
Zhang L, Gao M, Hu J, Zhang X, Wang K, Ashraf M. Modulation Role of Abscisic Acid (ABA) on Growth, Water Relations and Glycinebetaine Metabolism in Two Maize (Zea mays L.) Cultivars under Drought Stress. International Journal of Molecular Sciences. 2012; 13(3):3189-3202. https://doi.org/10.3390/ijms13033189
Chicago/Turabian StyleZhang, Lixin, Mei Gao, Jingjiang Hu, Xifeng Zhang, Kai Wang, and Muhammad Ashraf. 2012. "Modulation Role of Abscisic Acid (ABA) on Growth, Water Relations and Glycinebetaine Metabolism in Two Maize (Zea mays L.) Cultivars under Drought Stress" International Journal of Molecular Sciences 13, no. 3: 3189-3202. https://doi.org/10.3390/ijms13033189
APA StyleZhang, L., Gao, M., Hu, J., Zhang, X., Wang, K., & Ashraf, M. (2012). Modulation Role of Abscisic Acid (ABA) on Growth, Water Relations and Glycinebetaine Metabolism in Two Maize (Zea mays L.) Cultivars under Drought Stress. International Journal of Molecular Sciences, 13(3), 3189-3202. https://doi.org/10.3390/ijms13033189