Enhancement of Biocontrol Efficacy of Pichia carribbica to Postharvest Diseases of Strawberries by Addition of Trehalose to the Growth Medium

Abstract
:1. Introduction
2. Results and Discussion
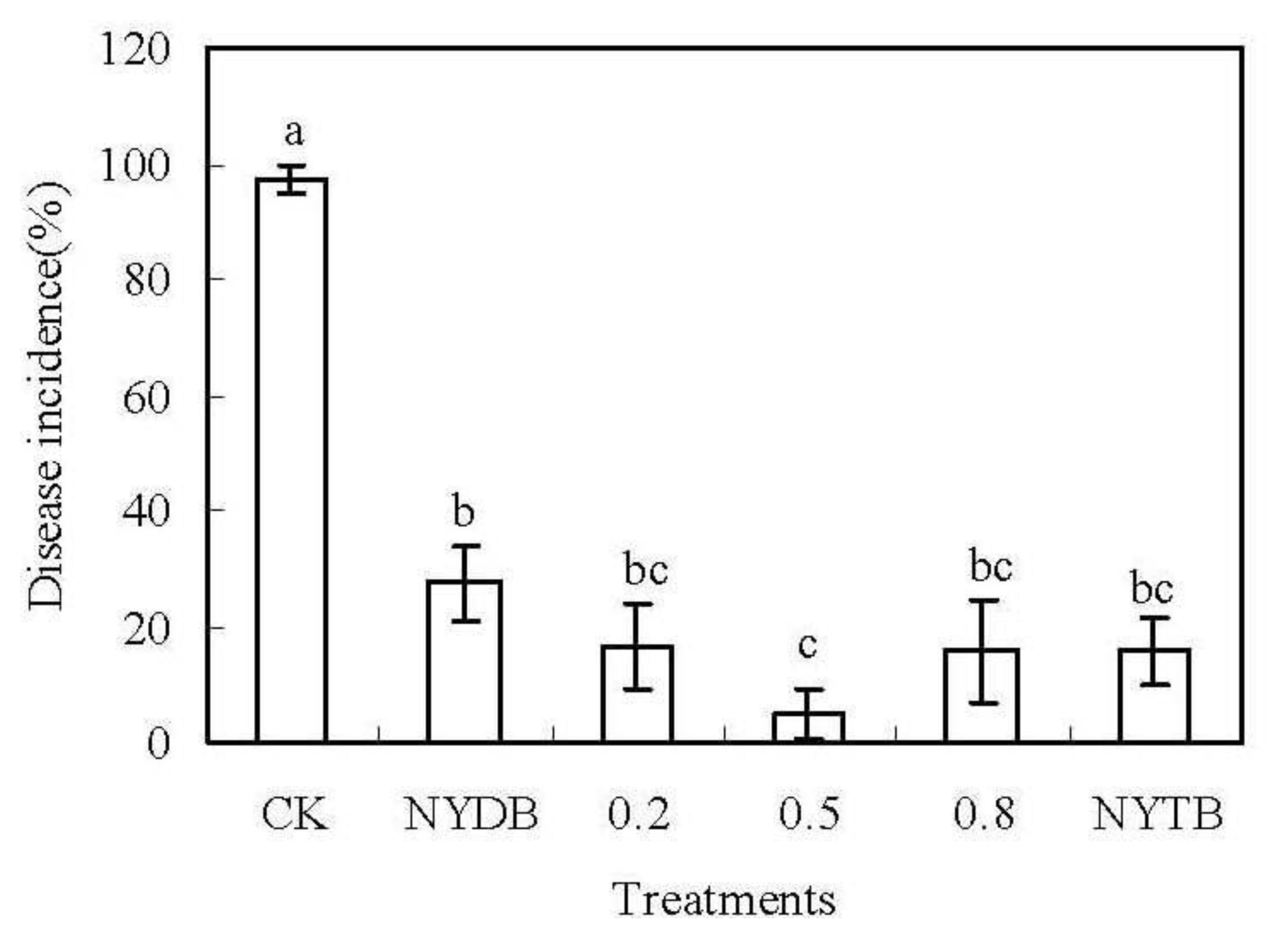
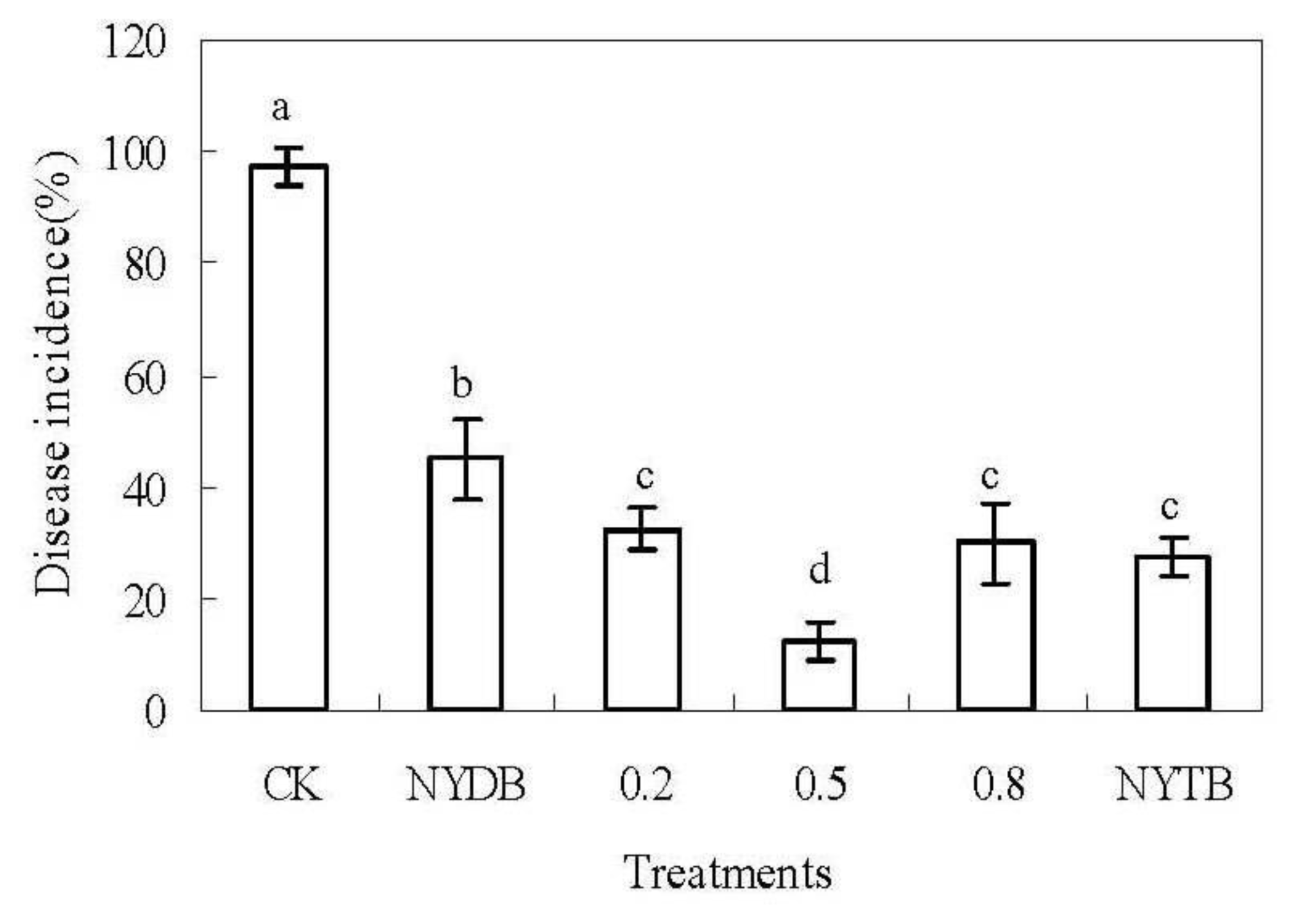
2.1. Efficacy of P. carribbica Harvested from Different Media in Controlling of Rhizopus Decay and Gray Mold Decay of Strawberries
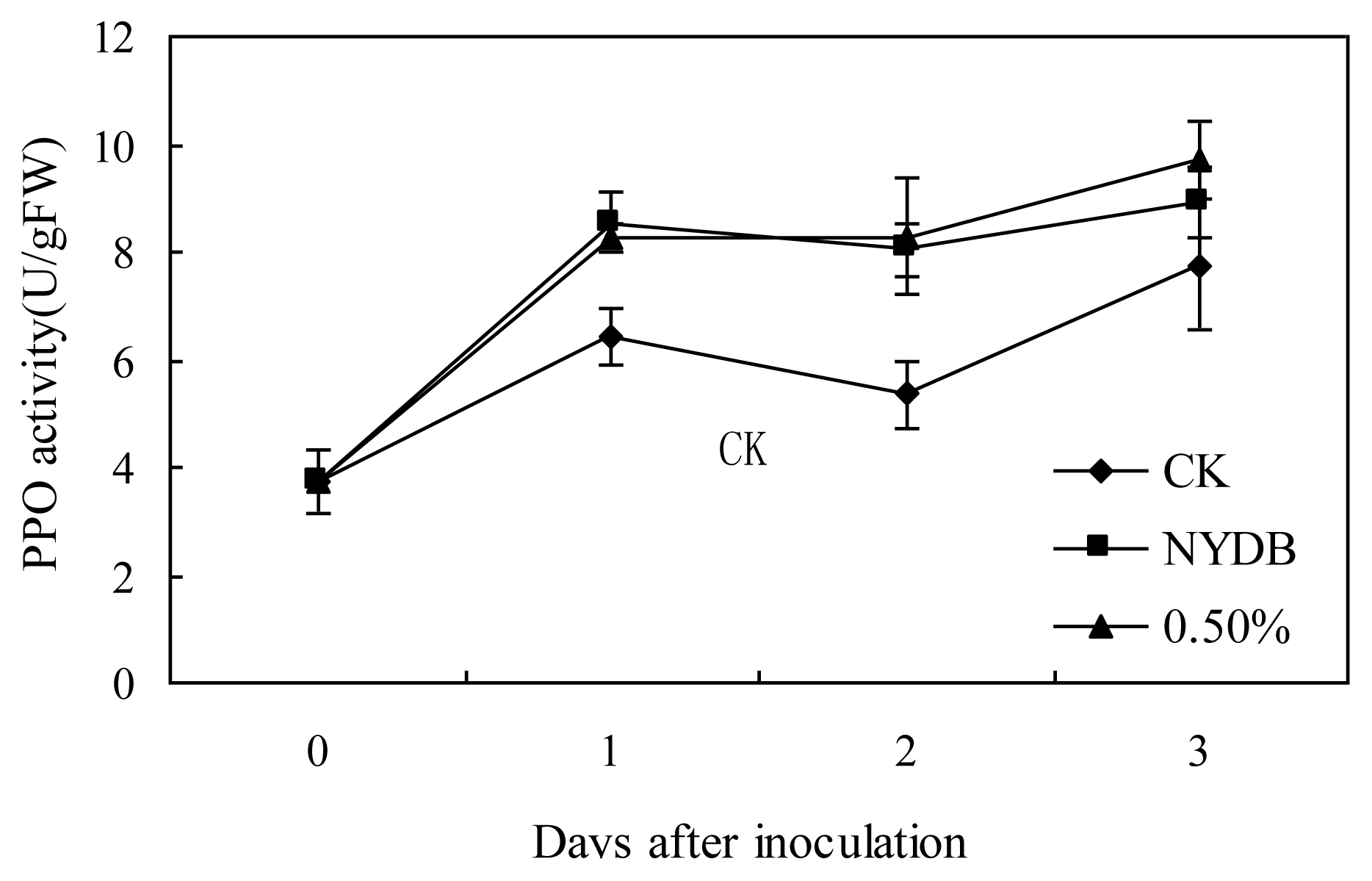
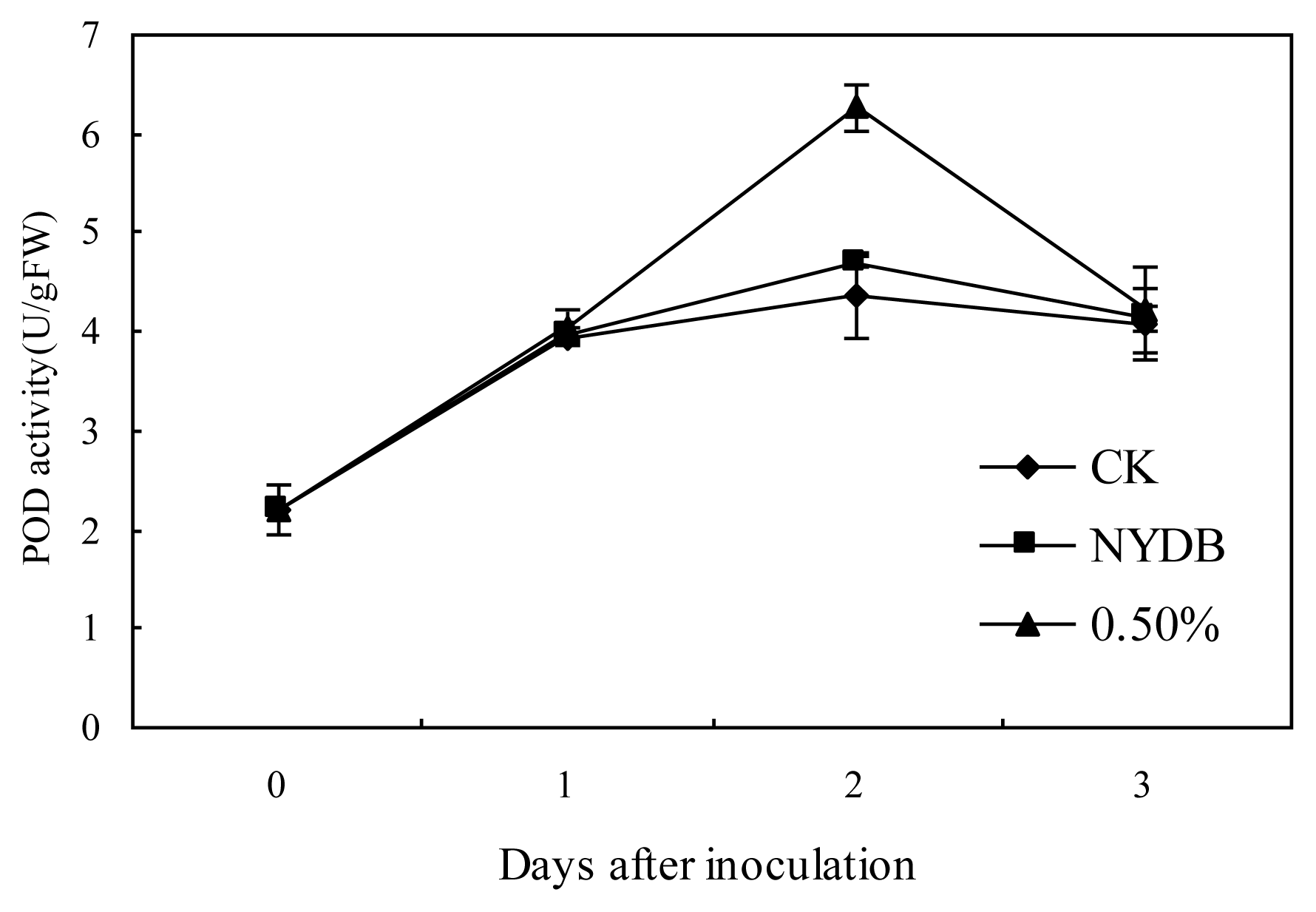
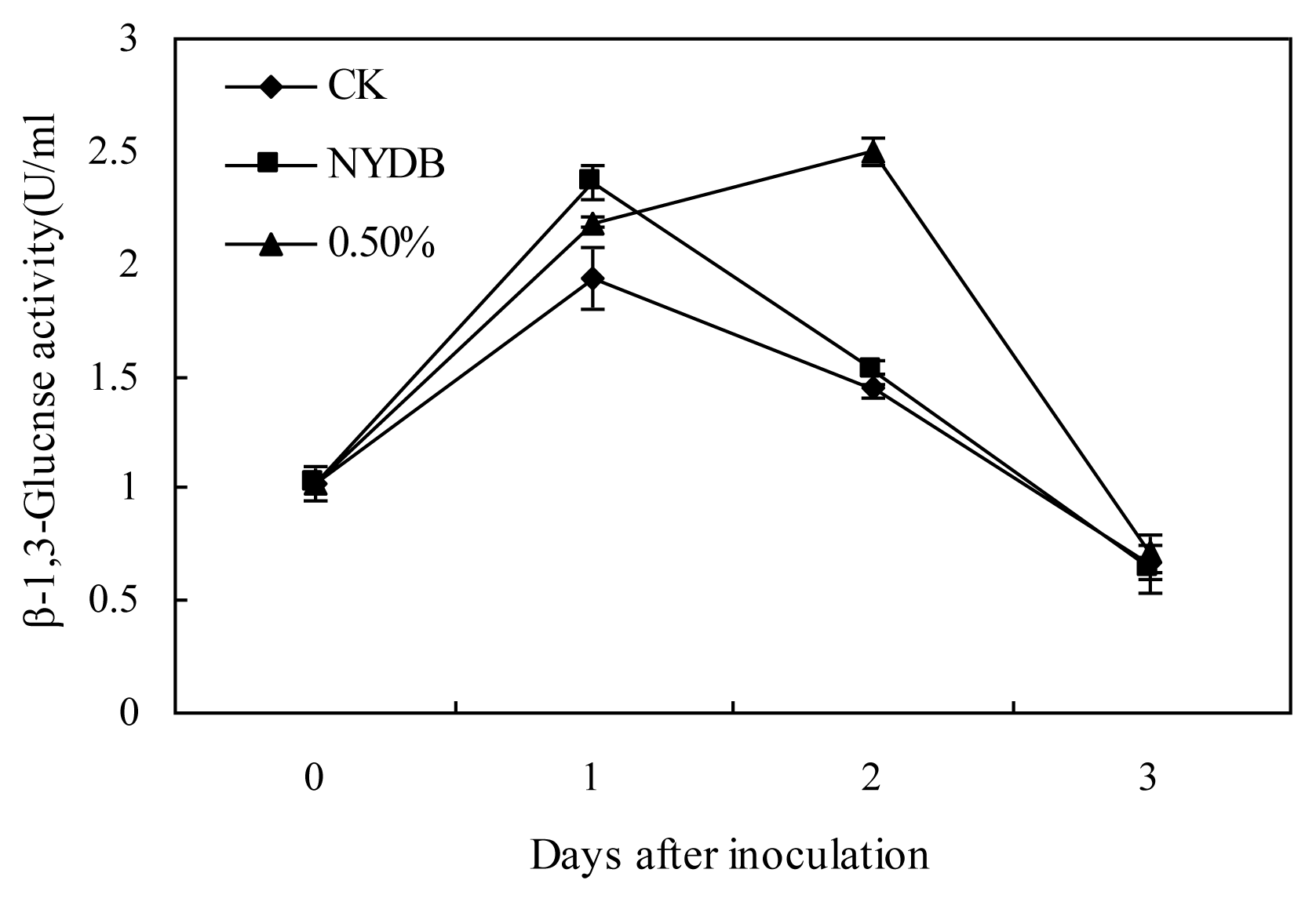
2.2. Effects of P. carribbica Harvested from Different Media on PPO, POD and β-1,3-glucanase Activities of Strawberries
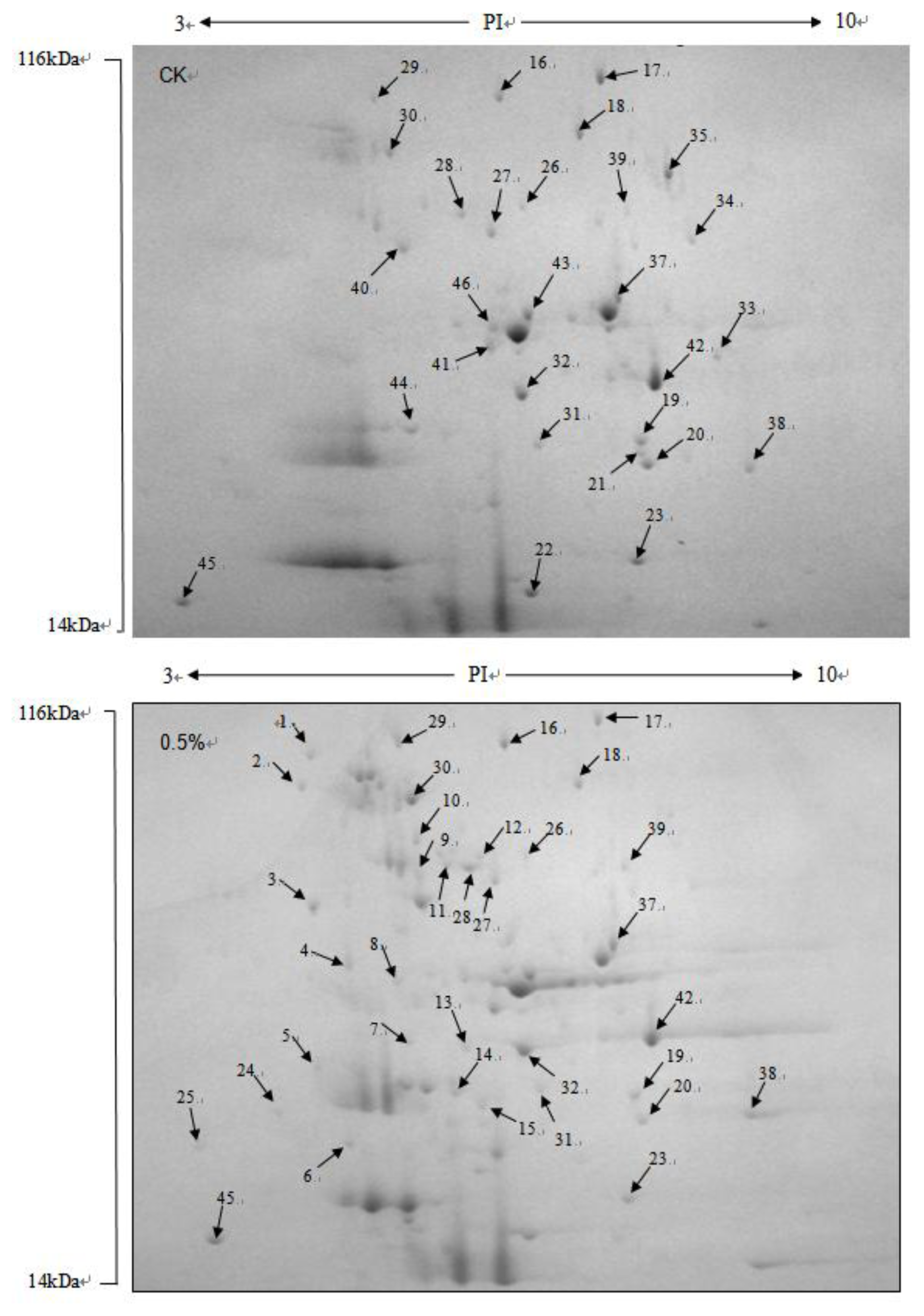
2.3. Identification of Differentially Expressed Proteins
3. Experimental Section
3.1. Antagonist and Growth Conditions
3.2. Fruits
3.3. Pathogen Inoculum
3.4. Efficacy of P. carribbica Harvested from Different Media in Controlling of Rhizopus Decay and Gray Mold Decay of Strawberries
3.5. Effects of P. carribbica Harvested from Different Media on PPO (Polyphenoloxidase), POD (Peroxidase) and β-1,3-glucanase Activities of Strawberries
3.6. Protein Sample Preparation
3.7. 2-DE and Image Analysis
3.8. Protein In-gel Digestion
3.9. Protein Identification by MALDI-TOF/TOF and Database Query
3.10. Statistical Analysis
4. Conclusions
Acknowledgements
References
- Romanazzi, G.; Nigro, F.; Ippolito, A.; Salerno, M. Effect of short hypobaric treatments on postharvest rots of sweet cherries, strawberries and table grapes. Postharvest Biol. Technol 2001, 22, 1–6. [Google Scholar]
- Wszelaki, A.L.; Mitcham, E.J. Effect of combinations of hot water dips, biological control and controlled atmospheres for control of gray mold on harvested strawberries. Postharvest Biol. Technol 2003, 27, 255–264. [Google Scholar]
- Norman, C. EPA sets new policy on pesticide cancer risks. Science 1988, 242, 366–367. [Google Scholar]
- Wisniewski, M.E.; Wilson, C.L. Biological control of postharvest diseases of fruits and vegetables: recent advance. HortScience 1992, 27, 94–98. [Google Scholar]
- Peng, G.; Sutton, J.C. Evaluation of microorganisms for biocontrol of Botrytis cinerea in strawberry. Can. J. Plant Pathol 1991, 13, 247–257. [Google Scholar]
- Swadling, I.R.; Jeffries, P. Isolation of microbial antagonists for biocontrol of grey mould disease of strawberries. Biocontrol Sci. Technol 1996, 6, 125–136. [Google Scholar]
- Lima, G.; Ippolito, A.; Nigro, F.; Salerno, M. Effectiveness of Aureobasidium pullulans and Candida oleophila against postharvest strawberry rots. Postharvest Biol. Technol 1997, 10, 169–178. [Google Scholar]
- Guinebretiere, M.H.; Nguyen-The, C.; Morrison, N.; Reich, M.; Nicot, P. Isolation and characterization of antagonists for the biocontrol of the postharvest wound pathogen Botrytis cinerea on strawberry fruits. J. Food Protect 2000, 63, 386–394. [Google Scholar]
- Zhang, H.Y.; Wang, L.; Dong, Y.; Jiang, S.; Cao, J.; Meng, R.J. Postharvest biological control of gray mold decay of strawberry with Rhodotorula glutinis. Biol. Control 2007, 40, 287–292. [Google Scholar]
- Droby, S.; Wsiniewski, M.; Ei-Ghaouth, A.; Wilson, C. Biological control of postharvest diseases of fruit and vegetables: Current achievements and future challenges. Acta Hortic 2003, 628, 703–713. [Google Scholar]
- Sharma, R.R.; Singh, D.; Singh, R. Biological control of postharvest diseases of fruits and vegetables by microbial antagonists: A review. Biol. Control 2009, 50, 205–221. [Google Scholar]
- Yu, T.; Wang, L.P.; Yin, Y.; Wang, Y.X.; Zheng, X.D. Effect of chitin on the antagonistic activity of Cryptococcus laurentii against Penicillium expansum in pear fruit. Int. J. Food Microbiol 2008, 122, 44–48. [Google Scholar]
- Ge, L.L.; Zhang, H.Y.; Chen, K.P.; Ma, L.C.; Xu, Z.L. Effect of chitin on the antagonistic activity of Rhodotorula glutinis against Botrytis cinerea in strawberries and the possible mechanisms involved. Food Chem 2010, 120, 490–495. [Google Scholar]
- Schiraldi, C.; Lernia, I.D.; Rosa, M.D. Trehalose production: exploiting novel approaches. Trends Biotechnol 2002, 20, 420–425. [Google Scholar]
- Mazzobre, M.F.; Buera, M.D.P. Combined effects of trehalose and cations on the thermal resistance of beta-galactosidase in freeze-dried systems. BBA-Gen. subjects 1999, 1473, 337–344. [Google Scholar]
- Crowe, J.H.; Crowe, L.M.; Chapman, D. Preservation of membrane in anhydrobiotic organism: the role of trehalose. Science 1984, 223, 701–703. [Google Scholar]
- Gancedo, C.; Flores, C.L. The importance of a functional trehalose biosynthetic pathway for the life of yeasts and fungi. FEMS Yeast Res 2004, 4, 351–359. [Google Scholar]
- Li, B.Q.; Zhou, Z.W.; Tian, S.P. Combined effects of endo- and exogenous trehalose on stress tolerance and biocontrol efficacy of two antagonistic yeasts. Biol. Control 2008, 46, 187–193. [Google Scholar]
- Richards, A.B.; Krakowka, S.; Dexter, L.B.; Schmidd, H.; Wolterbeeke, A.P.M.; Waalkens-Berendsene, D.H.; Shigoyuki, A.; Kurimotof, M. Trehalose: a review of properties, history of use and human tolerance, and results of multiple safety studies. Food Chem. Toxicol 2002, 40, 871–898. [Google Scholar]
- Mayer, A.M.; Harel, E. Polyphenol oxidases in plants. Phytochemistry 1979, 18, 193–215. [Google Scholar]
- Jung, W.J.; Jin, Y.L.; Kim, Y.C.; Kim, K.Y.; Park, R.D.; Kim, T.H. Inoculation of Paenibacillus illinoisensis alleviates root mortality, activates of lignification-related enzymes, and induction of the isozymes in pepper plants infected by Phytophthora capsici. Biol. Control 2004, 30, 645–652. [Google Scholar]
- Mohammadi, M.; Kazemi, H. Changes in peroxidase and polyphenol oxidase activities in susceptible and resistant wheat heads inoculated with Fusarium graminearum and induced resistance. Plant Sci 2002, 162, 491–498. [Google Scholar]
- Passardi, F.; Penel, C.; Dunand, C. Performing the paradoxical: how plant peroxidases modify the cell wall. Trends Plant Sci 2004, 9, 534–540. [Google Scholar]
- Lurie, S.; Fallik, E.; Handros, A.; Shapira, R. The possible involvement of peroxidase in resistance to Botrytis cinereain heat treated tomato fruit. Physiol. Mol. Plant P 1997, 50, 141–149. [Google Scholar]
- Yu, T.; Zheng, X.D. Salicylic acid enhances biocontrol efficacy of the antagonist Cryptococcus laurentii in apple fruit. J. Plant Growth Regul 2006, 25, 166–174. [Google Scholar]
- Joosten, M.H.A.J.; Verbakel, H.M.; Nettekoven, M.E.; van Leeuwen, J.; van der Vossen, R.T.M.; de Wit, P.J.G.M. The phytopathogenic fungus Cladosporium fulvum is not sensitive to the chitinase and β-1,3-glucanase defence proteins of its host, tomato. Physiol. Mol. Plant P 1995, 46, 45–59. [Google Scholar]
- Charles, M.T.; Tano, K.; Asselin, A.; Arul, J. Physiological basis of UV-C induced resistance to Botrytis cinerea in tomato fruit. V. Constitutive defence enzymes and inducible pathogenesis-related proteins. Postharvest Biol. Technol 2009, 51, 414–424. [Google Scholar]
- Qin, G.Z.; Tian, S.P.; Xu, Y.; Wan, Y.K. Enhancement of biocontrol efficacy of antagonistic yeasts by salicylic acid in sweet cherry fruit. Physiol. Mol. Plant Pathol 2003, 62, 147–154. [Google Scholar]
- Kurtzman, C.P.; Fell, J.W. Definition, Classification and Nomenclature of the Yeasts. In The Yeasts, 4th ed; CBS-Fungal Biodiversity Centre: Utrecht, The Netherlands, 1998; pp. 3–5. [Google Scholar]
- Li, S.S.; Cheng, C.; Li, Z.; Chen, J.Y.; Yan, B.; Han, B.Z.; Reeves, M. Yeast species associated with wine grapes in China. Int. J. Food Microbiol 2010, 138, 85–90. [Google Scholar]
- Aquino-Bolaños, E.N.; Mercado-Silva, E. Effects of polyphenol oxidase and peroxidase activity, phenolics and lignin content on the browning of cut jicama. Postharvest Biol. Technol 2004, 33, 275–283. [Google Scholar]
- Ippolito, A.; EI-Ghaouth, A.; Wilson, C.L.; Wisniewski, M. Control of postharvest decay of apple fruit by Aureobasidium pullulans and induction of defense responses. Postharvest Biol. Technol 2000, 19, 265–272. [Google Scholar]
- Li, B.Q.; Lai, T.F.; Qin, G.Z.; Tian, S.P. Ambient pH Stress Inhibits Spore Germination of Penicillium expansum by Impairing Protein Synthesis and Folding: A Proteomic-Based Study. J. Proteome Res 2010, 9, 298–307. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem 1976, 72, 248–254. [Google Scholar]
- Wang, Y.; Yang, L.M.; Xu, H.B.; Li, Q.F.; Ma, Z.Q.; Chu, C.G. Differential proteomic analysis of proteins in wheat spikes induced by Fusarium graminearum. Proteomics 2005, 5, 4496–4503. [Google Scholar]
- Zhang, L.; Yu, Z.F.; Jiang, L.; Jiang, J.; Luo, H.B.; Fu, L.R. Effect of post-harvest heat treatment on proteome change of peach fruit during ripening. J. Proteomics 2011, 74, 1135–1149. [Google Scholar]
- Matrixscience Home Page. http://www.matrixscience.com accessed on 21 March 2012.
- Majoul, T.; Bancel, E.; Tribol, E.; Hamida, J.B.; Branlard, G. Proteomic analysis of the effect of heat stress on hexaploid wheat grain: Characterization of heat-responsive proteins from total endosperm. Proteomics 2003, 3, 175–183. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot | Protein name | NCBI accession | Mass | PI | Species | Score |
|---|---|---|---|---|---|---|
| 1 | 50S ribosomal protein | gi|116495724 | 12511 | 4.54 | Lactobacillus casei ATCC 334 | 66 |
| 2 | UDP-galactopyranose mutase | gi|326332321 | 45474 | 5.11 | Nocardioidaceae bacterium Broad-1 | 68 |
| 3 | extracellular solute-binding protein | gi|148546296 | 45335 | 6.02 | Pseudomonas putida F1 | 62 |
| 4 | eukaryotic initiation factor 4A | gi|146422477 | 44615 | 4.91 | Meyerozyma guilliermondii ATCC 6260 | 100 |
| 5 | predicted protein | gi|145345294 | 42354 | 9.84 | Ostreococcus lucimarinus CCE9901 | 65 |
| 6 | hypothetical protein SCHCODRAFT_38806 | gi|302688397 | 10646 | 4.25 | Schizophyllum commune H4-8 | 51 |
| 8 | elongation factor Tu | gi|587590 | 43823 | 5.06 | Wolinella succinogenes | 75 |
| 9 | electron transfer flavoprotein subunit beta | gi|162447962 | 29409 | 8.87 | Acholeplasma laidlawii PG-8A | 69 |
| 10 | hypothetical protein bthur0013_57560 | gi|228911633 | 45801 | 6.33 | Bacillus thuringiensis IBL 200 | 62 |
| 11 | HNH nuclease | gi|220919264 | 38869 | 9.73 | Anaeromyxobacter dehalogenans 2CP-1 | 65 |
| 12 | hypothetical protein PGUG_04322 | gi|146415246 | 69936 | 5.30 | Meyerozyma guilliermondii ATCC 6260 | 71 |
| 13 | hypothetical protein bcere0017_55820 | gi|229119349 | 28408 | 8.55 | Bacillus cereus Rock1-3 | 67 |
| 14 | hypothetical protein PGUG_00294 | gi|146421948 | 35820 | 5.22 | Meyerozyma guilliermondii ATCC 6260 | 104 |
| 15 | xylose reductase | gi|4103055 | 36076 | 5.58 | Meyerozyma guilliermondii | 66 |
| 16 | conserved hypothetical protein | gi|146421560 | 84405 | 5.92 | Meyerozyma guilliermondii ATCC 6260 | 90 |
| 17 | conserved hypothetical protein | gi|146420955 | 85681 | 5.68 | Meyerozyma guilliermondii ATCC 6260 | 134 |
| 18 | hypothetical protein CLOSCI_01190 | gi|167758847 | 66473 | 4.76 | Clostridium scindens ATCC 35704 | 68 |
| 19 | translation elongation factor | gi|47176804 | 22272 | 5.46 | Meyerozyma guilliermondii | 68 |
| 20 | conserved hypothetical protein | gi|146417765 | 34916 | 7.17 | Meyerozyma guilliermondii ATCC 6260 | 96 |
| 21 | translation elongation factor | gi|47176804 | 22272 | 5.46 | Meyerozyma guilliermondii | 82 |
| 22 | Melibiase subfamily, putative | gi|254503502 | 77433 | 5.48 | Labrenzia alexandrii DFL-11 | 86 |
| 24 | hypothetical protein PGUG_05024 | gi|146414197 | 32004 | 7.77 | Meyerozyma guilliermondii ATCC 6260 | 80 |
| 25 | unnamed protein product | gi|189054178 | 65980 | 7.62 | Homo sapiens | 87 |
| 26 | hypothetical protein | gi|67601196 | 32428 | 9.67 | Cryptosporidium hominis TU502 | 63 |
| 27 | hypothetical protein PGUG_02894 | gi|146418399 | 57751 | 5.57 | Meyerozyma guilliermondii ATCC 6260 | 89 |
| 28 | transcriptional regulator, laci family | gi|315498361 | 34890 | 5.36 | Asticcacaulis excentricus CB 48 | 87 |
| 29 | nitrite and sulfite reductase 4Fe-4S region | gi|118579082 | 23982 | 8.66 | Pelobacter propionicus DSM 2379 | 71 |
| 30 | heat shock protein SSB1 | gi|146420661 | 66250 | 5.29 | Meyerozyma guilliermondii ATCC 6260 | 162 |
| 31 | cytochrome d ubiquinol oxidase subunit III | gi|156973199 | 16378 | 4.72 | Vibrio harveyi ATCC BAA-1116 | 71 |
| 32 | FAD dependent oxidoreductase | gi|225011369 | 41131 | 8.66 | Flavobacteria bacterium MS024-2A | 73 |
| 33 | PREDICTED: uncharacterized glycosyltransferase AER61- like | gi|109036798 | 62132 | 6.39 | Macaca mulatta | 61 |
| 34 | hypothetical protein KSE_48990 | gi|311898269 | 37649 | 8.94 | Kitasatospora setae KM-6054 | 48 |
| 35 | isocitrate lyase | gi|146413757 | 61766 | 6.31 | Meyerozyma guilliermondii ATCC 6260 | 88 |
| 36 | conserved hypothetical protein | gi|238064394 | 46346 | 11.42 | Micromonospora sp. ATCC 39149 | 78 |
| 37 | DEHA2D06160p | gi|50420381 | 54282 | 5.68 | Debaryomyces hansenii CBS767 | 72 |
| 38 | glyceraldehyde-3-phosphate dehydrogenase | gi|146419367 | 35717 | 6.60 | Meyerozyma guilliermondii ATCC 6260 | 82 |
| 39 | hypothetical protein HMU03290 | gi|291276562 | 8800 | 9.70 | Helicobacter mustelae 12198 | 67 |
| 40 | conserved hypothetical protein | gi|313836798 | 12685 | 6.31 | Propionibacterium acnes HL037PA2 78 | |
| 41 | possible gp16 protein | gi|227496471 | 16688 | 4.70 | Actinomyces urogenitalis DSM 15434 | 79 |
| 42 | conserved hypothetical protein | gi|301168105 | 15154 | 9.27 | Bacteriovorax marinus SJ | 82 |
| 43 | enolase 1 | gi|146415384 | 46951 | 5.42 | Meyerozyma guilliermondii ATCC 6260 | 84 |
| 44 | rod shape-determining protein MreC, putative | gi|21673403 | 32080 | 9.73 | Chlorobium tepidum TLS | 86 |
| 45 | hypothetical protein LVIS_1868 | gi|116334433 | 7652 | 5.55 | Lactobacillus brevis ATCC 367 | 85 |
| 46 | enolase 1 | gi|146415384 | 46951 | 5.42 | Meyerozyma guilliermondii ATCC 6260 | 97 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhao, L.; Zhang, H.; Li, J.; Cui, J.; Zhang, X.; Ren, X. Enhancement of Biocontrol Efficacy of Pichia carribbica to Postharvest Diseases of Strawberries by Addition of Trehalose to the Growth Medium. Int. J. Mol. Sci. 2012, 13, 3916-3932. https://doi.org/10.3390/ijms13033916
Zhao L, Zhang H, Li J, Cui J, Zhang X, Ren X. Enhancement of Biocontrol Efficacy of Pichia carribbica to Postharvest Diseases of Strawberries by Addition of Trehalose to the Growth Medium. International Journal of Molecular Sciences. 2012; 13(3):3916-3932. https://doi.org/10.3390/ijms13033916
Chicago/Turabian StyleZhao, Lina, Hongyin Zhang, Jun Li, Jinghua Cui, Xiaoyun Zhang, and Xiaofeng Ren. 2012. "Enhancement of Biocontrol Efficacy of Pichia carribbica to Postharvest Diseases of Strawberries by Addition of Trehalose to the Growth Medium" International Journal of Molecular Sciences 13, no. 3: 3916-3932. https://doi.org/10.3390/ijms13033916
APA StyleZhao, L., Zhang, H., Li, J., Cui, J., Zhang, X., & Ren, X. (2012). Enhancement of Biocontrol Efficacy of Pichia carribbica to Postharvest Diseases of Strawberries by Addition of Trehalose to the Growth Medium. International Journal of Molecular Sciences, 13(3), 3916-3932. https://doi.org/10.3390/ijms13033916