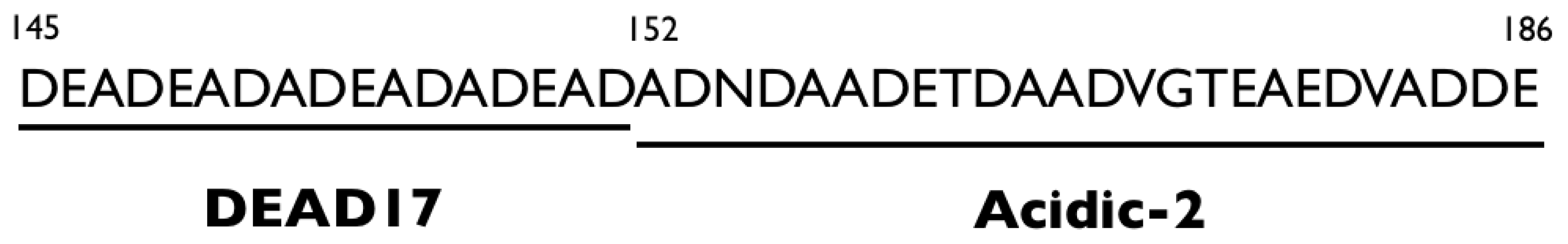
Cooperative Modulation of Mineral Growth by Prismatic-Associated Asprich Sequences and Mg(II)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
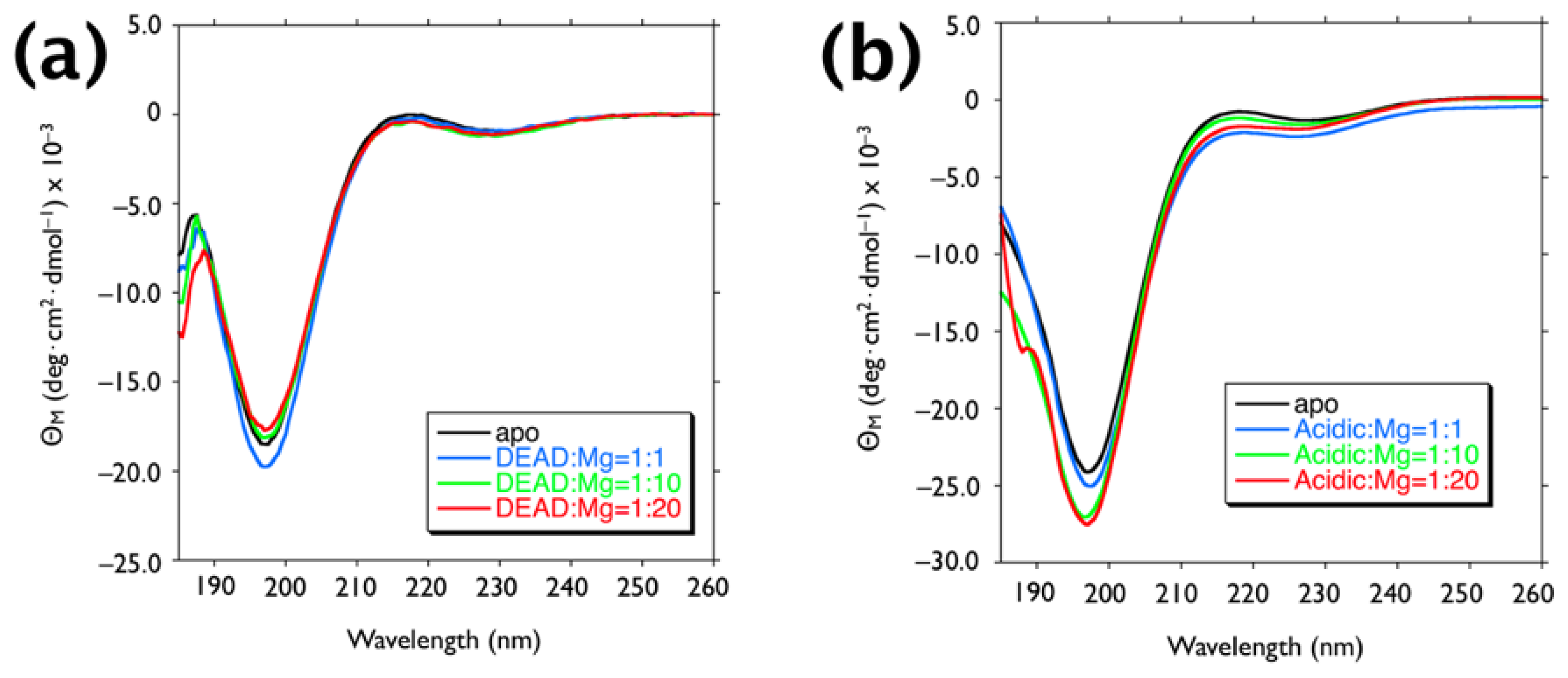
2.1. Mineralization of Calcite
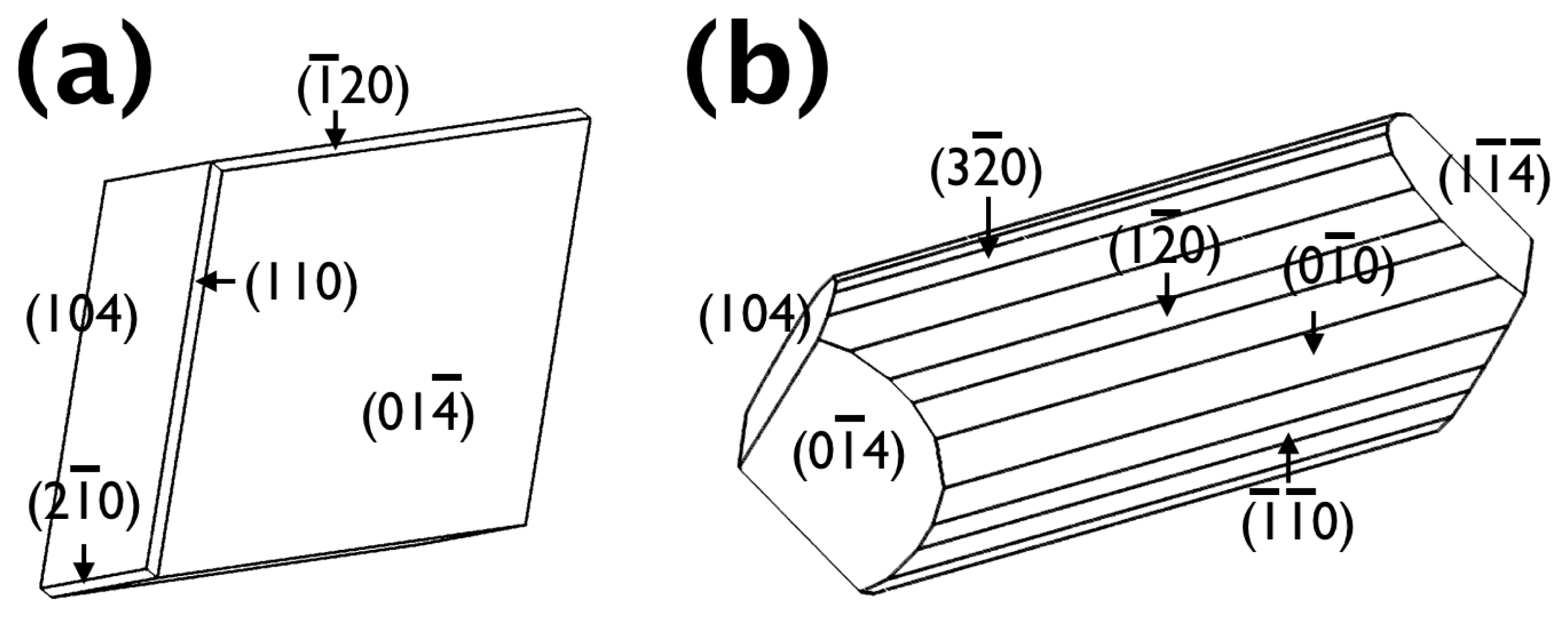
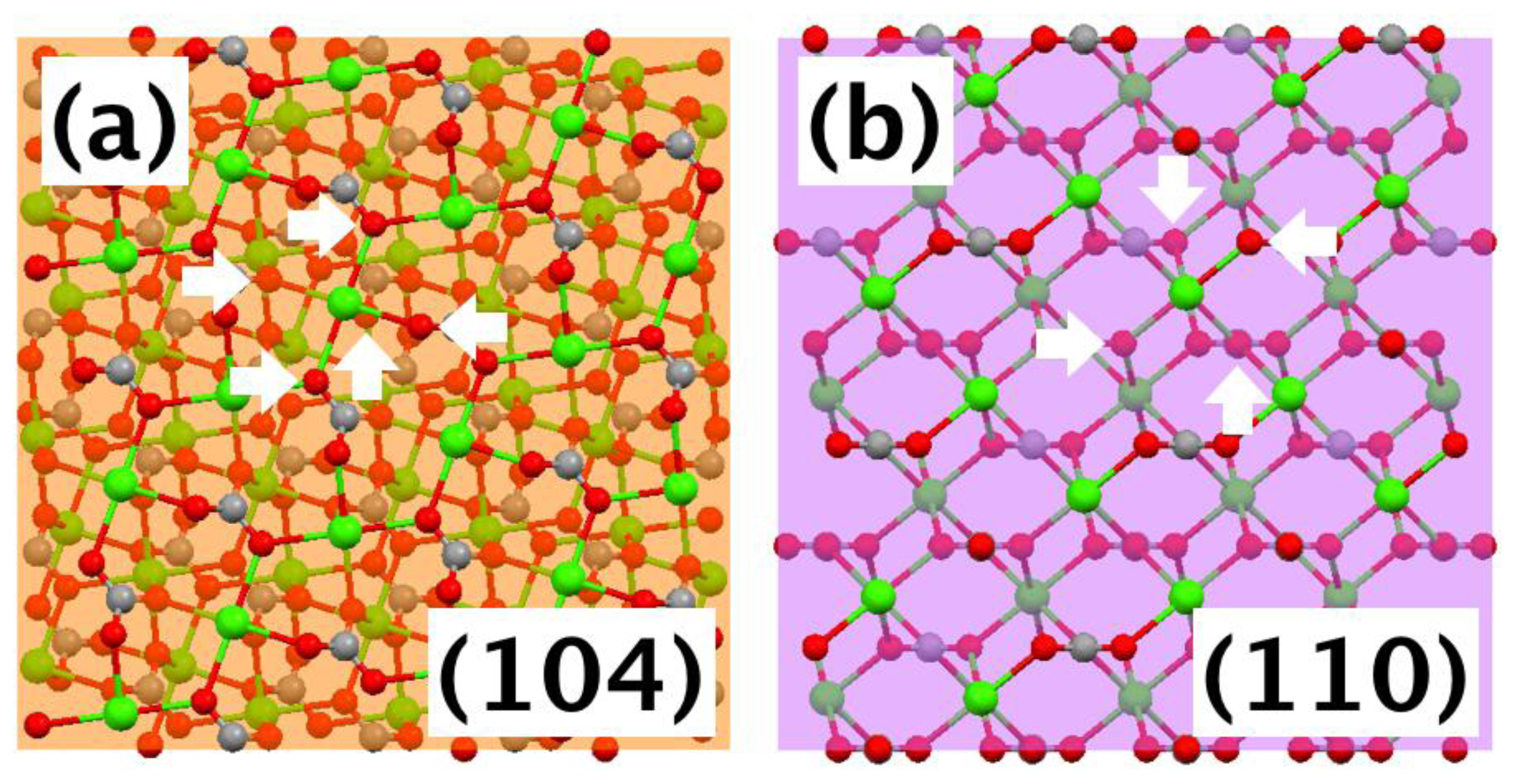
2.2. The Origin of the Morphology Modification
3. Experimental Section
3.1. In Vitro Mineralization and Crystal Analysis
3.2. Circular Dichroism Spectrometry
4. Conclusions
Acknowledgments
References
- Lowenstam, H.A.; Weiner, S. On Biomineralization; Oxford University Press: New York NY, USA, 1989. [Google Scholar]
- Weiner, S.; Addadi, L. Design strategies in mineralized biological materials. J. Mater. Chem 1997, 7, 689–702. [Google Scholar]
- Almqvist, N.; Thomson, N.H.; Smith, B.L.; Stucky, G.D.; Morse, D.E.; Hansma, P.K. Methods for fabricating and characterizing a new generation of biomimetic materials. Mater. Sci. Eng. C 1999, C7, 37–43. [Google Scholar]
- Li, X.D.; Chang, W.-C.; Chao, Y.J.; Wang, R.Z.; Chang, M. Nanoscale structural and mechanical characterization of a natural nanocomposite materials: the shell of red abalone. Nano Lett 2004, 4, 613–617. [Google Scholar]
- Belcher, A.M.; Wu, X.H.; Christensen, R.J.; Hansma, P.K.; Stucky, G.D.; Morse, D.E. Control of crystal phase switching and orientation by soluble mollusc-shell proteins. Nature 1996, 381, 56–58. [Google Scholar]
- Evans, J.S. “Apples” and “oranges”: comparing the structural aspects of biomineral- and ice-interaction proteins. Curr. Opin. Colloid Interface Sci 2003, 8, 48–54. [Google Scholar]
- Sarikaya, M.; Tamerler, C.; Jen, A.K.-Y.; Schulten, K.; Baneyx, F. Molecular biomimetics: nanotechnology through biology. Nat. Mater 2003, 2, 577–585. [Google Scholar]
- Cölfen, H.; Antonietti, M. Mesocrystals: Inorganic superstructures made by highly parallel crystallization and controlled alignment. Angew. Chem. Int. Ed 2005, 44, 5576–5591. [Google Scholar]
- Lippmann, F. Sedimentary Carbonate Minerals; Springer-Verlag: Berlin, Germany, 1973. [Google Scholar]
- Raz, S.; Hamilton, P.C.; Wilt, F.H.; Weiner, S.; Addadi, L. The transient phase of amorphous calcium carbonate in sea urchin larval spicules: the involvement of proteins and magnesium ions in its formation and stabilization. Adv. Funct. Mater 2003, 13, 480–486. [Google Scholar]
- Han, Y.-J.; Wysocki, L.M.; Thanawala, M.S.; Siegrist, T.; Aizenberg, J. Template-dependent morphogenesis of oriented calcite crystals in the presence of magnesium ions. Angew. Chem. Int. Ed 2005, 44, 2386–2390. [Google Scholar]
- Davis, K.J.; Dove, P.M.; Wasylenki, L.E.; DeYoreo, J.J. Morphological consequences of differential Mg2+ incorporation at structurally distinct steps on calcite. Am. Mineral 2004, 89, 714–720. [Google Scholar]
- Gotliv, B.-A.; Kessler, N.; Sumerel, J.L.; Morse, D.E.; Tuross, N.; Addadi, L.; Weiner, S. Asprich: A novel aspartic acid-rich protein family from the prismatic shell matrix of the bivalve Atrina rigida. Chem. Bio. Chem 2005, 6, 304–314. [Google Scholar]
- Ndao, M.; Keene, E.; Amos, F.A.; Rewari, G.; Ponce, C.B.; Estroff, L.; Evans, J.S. Intrinsically disordered mollusk shell prismatic protein that modulates calcium carbonate crystal growth. Biomacromolecules 2010, 11, 2539–2544. [Google Scholar]
- Ndao, M.; Ponce, C.B.; Evans, J.S. Evidence of self-association and aggregation-promoting sequences within the “acidic” biomineralization protein, Asprich 3. Biochemistry.
- Kim, I.W.; Darragh, M.R.; Orme, C.; Evans, J.S. Molecular “tuning” of crystal growth by nacre-associated polypeptides. Cryst. Growth Des 2006, 6, 5–10. [Google Scholar]
- Kim, I.W.; Giocondi, J.L.; Orme, C.; Collino, S.; Evans, J.S. Morphological and kinetic transformation of calcite crystal growth by prismatic-associated Asprich sequences. Cryst. Growth Des 2008, 8, 1154–1160. [Google Scholar]
- Collino, S.; Kim, I.W.; Evans, J.S. Identification of an “acidic” C-terminal mineral modification sequence from the mollusk shell protein Asprich. Cryst. Growth Des 2006, 6, 839–842. [Google Scholar]
- Graf, D.L. Crystallographic tables for the rhombohedral carbonates. Am. Mineral 1961, 46, 1283–1316. [Google Scholar]
- Chung, J.; Kim, I.W. Oriented crystallization of xanthine derivatives sublimated on self-assembled monolayers. Korean J. Chem. Eng 2011, 28, 232–238. [Google Scholar]
- Falk, R.L. The natural history of crystalline calcium carbonate: effect of magnesium content and salinity. J. Sediment. Petrol 1974, 44, 40–53. [Google Scholar]
- DeYoreo, J.J.; Dove, P.M. Shaping crystals with biomolecules. Science 2004, 306, 1301–1302. [Google Scholar]
- Hartman, P.; Perdok, W.G. On the relations between structure and morphology of crystals. Acta Crystallogr 1955, 8, 49–52. [Google Scholar]
- Heijnen, W.M.M. The morphology of gel grown calcite (in Russian). N. Jb. Miner. Mh 1985, 8, 357–381. [Google Scholar]
- Aquilano, D.; Calleri, M.; Natoli, E.; Rubbo, M.; Sgualdino, G. The {104} cleavage rhombohedron of calcite: theoretical equilibrium properties. Mater. Chem. Phys 2000, 66, 159–163. [Google Scholar]
- Walton, A.G. The Formation and Properties of Precipitates; Interscience Publishers: New York, USA, 1967; Volume Chapter 5. [Google Scholar]
- Schäffer, T.E.; Ionescu-Zanetti, C.; Proksch, R.; Fritz, M.; Walters, D.A.; Almqvist, N.; Zaremba, C.M.; Belcher, A.M.; Smith, B.L.; Stucky, G.D.; et al. Does abalone nacre form by heteroepitaxial nucleationor by growth through mineral bridges? Chem. Mater 1997, 9, 1731–1740. [Google Scholar]
- Pokroy, B.; Quintana, J.P.; Caspi, E.N.; Berner, A.; Zolotoyabko, E. Anisotropic lattice distortions in biogenic aragonite. Nat. Mater 2004, 3, 900–902. [Google Scholar]





© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kim, I.W.; Collino, S.; Evans, J.S. Cooperative Modulation of Mineral Growth by Prismatic-Associated Asprich Sequences and Mg(II). Int. J. Mol. Sci. 2012, 13, 3949-3958. https://doi.org/10.3390/ijms13033949
Kim IW, Collino S, Evans JS. Cooperative Modulation of Mineral Growth by Prismatic-Associated Asprich Sequences and Mg(II). International Journal of Molecular Sciences. 2012; 13(3):3949-3958. https://doi.org/10.3390/ijms13033949
Chicago/Turabian StyleKim, Il Won, Sebastiano Collino, and John Spencer Evans. 2012. "Cooperative Modulation of Mineral Growth by Prismatic-Associated Asprich Sequences and Mg(II)" International Journal of Molecular Sciences 13, no. 3: 3949-3958. https://doi.org/10.3390/ijms13033949