Proteins Involved in Distinct Phases of Cold Hardening Process in Frost Resistant Winter Barley (Hordeum vulgare L.) cv Luxor

Abstract
:1. Introduction
2. Results and Discussion
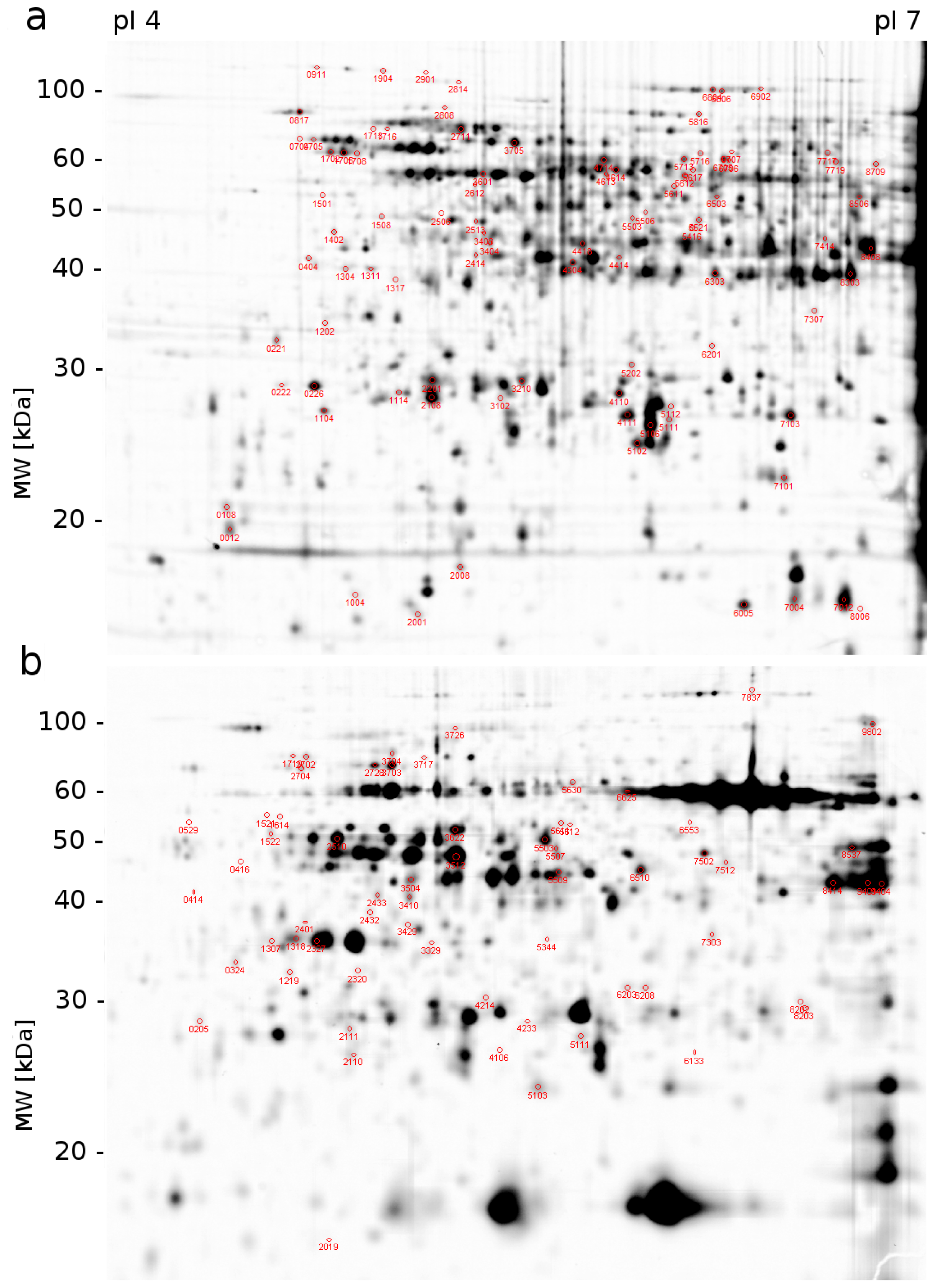
2.1. Proteomic Analysis
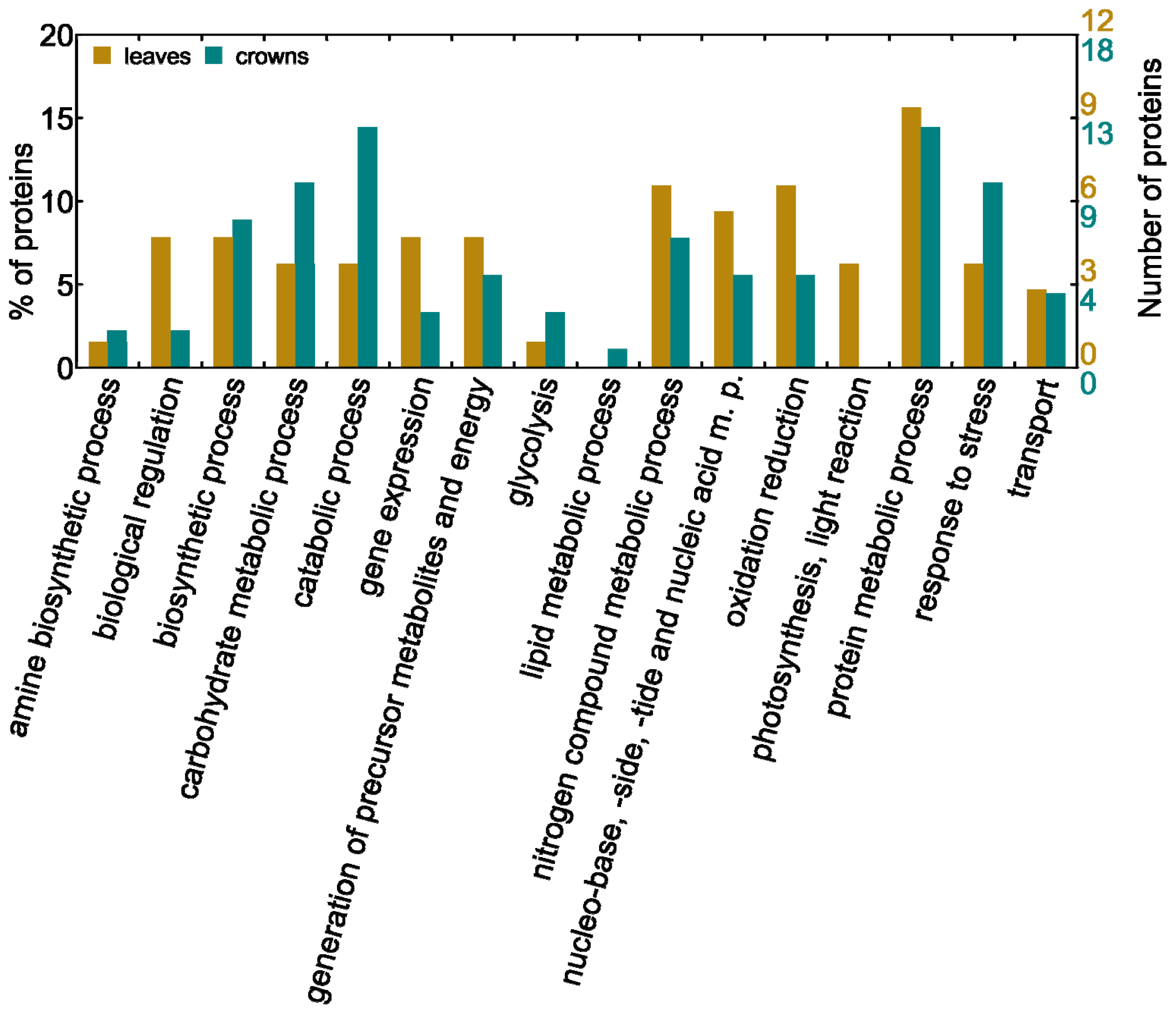
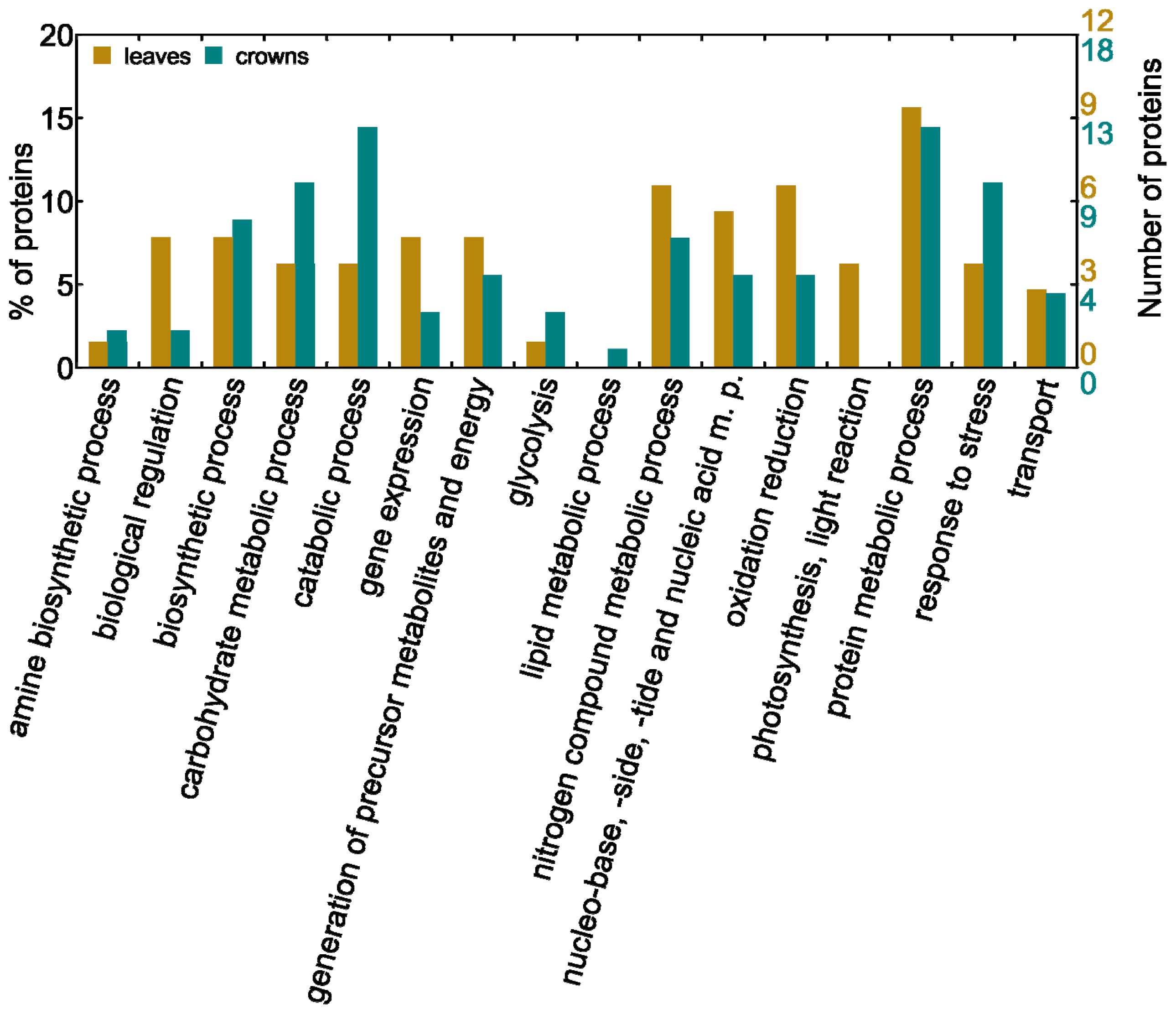
2.2. Biological Processes Influenced by Low Temperatures in Leaves and Crowns
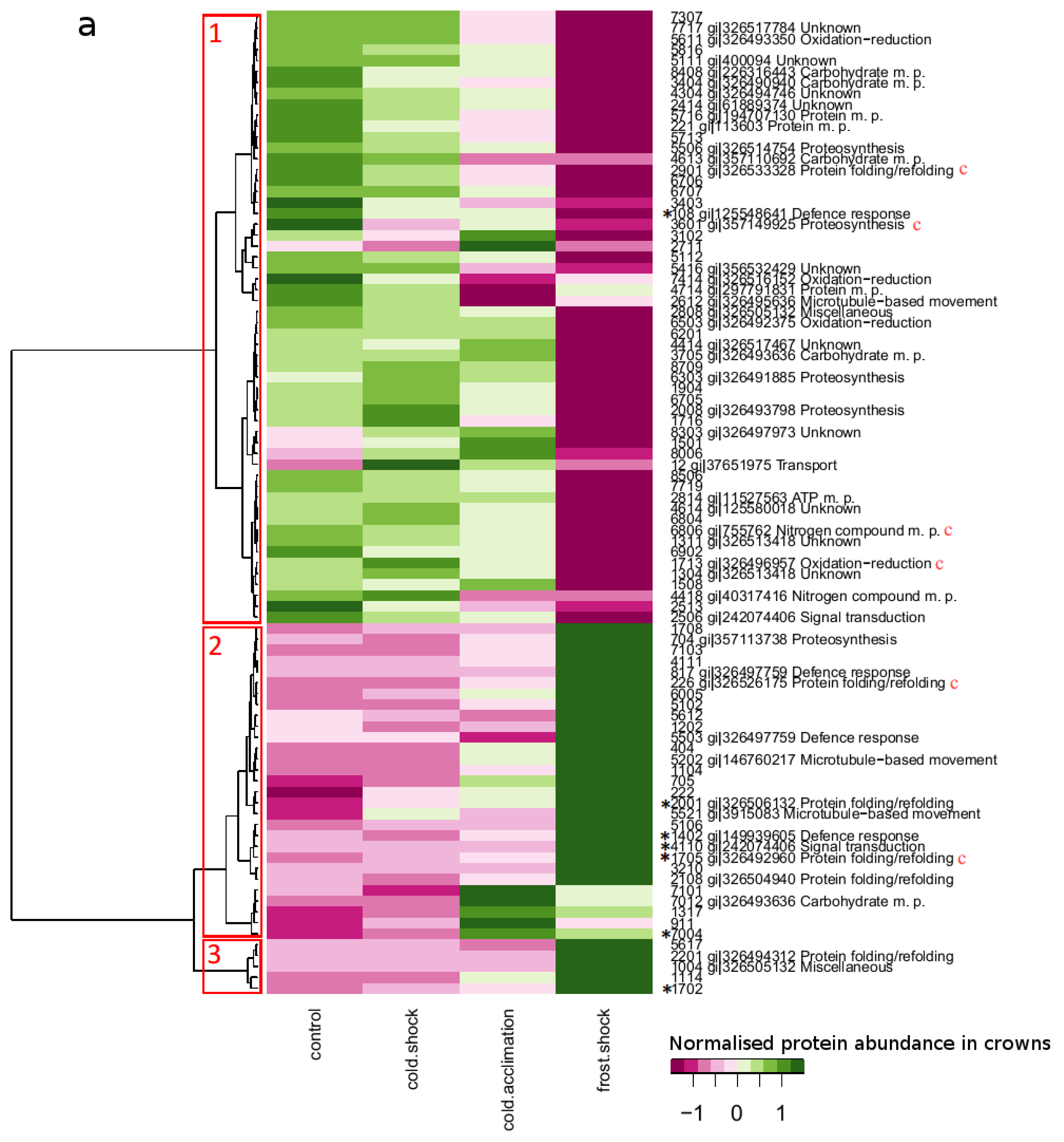
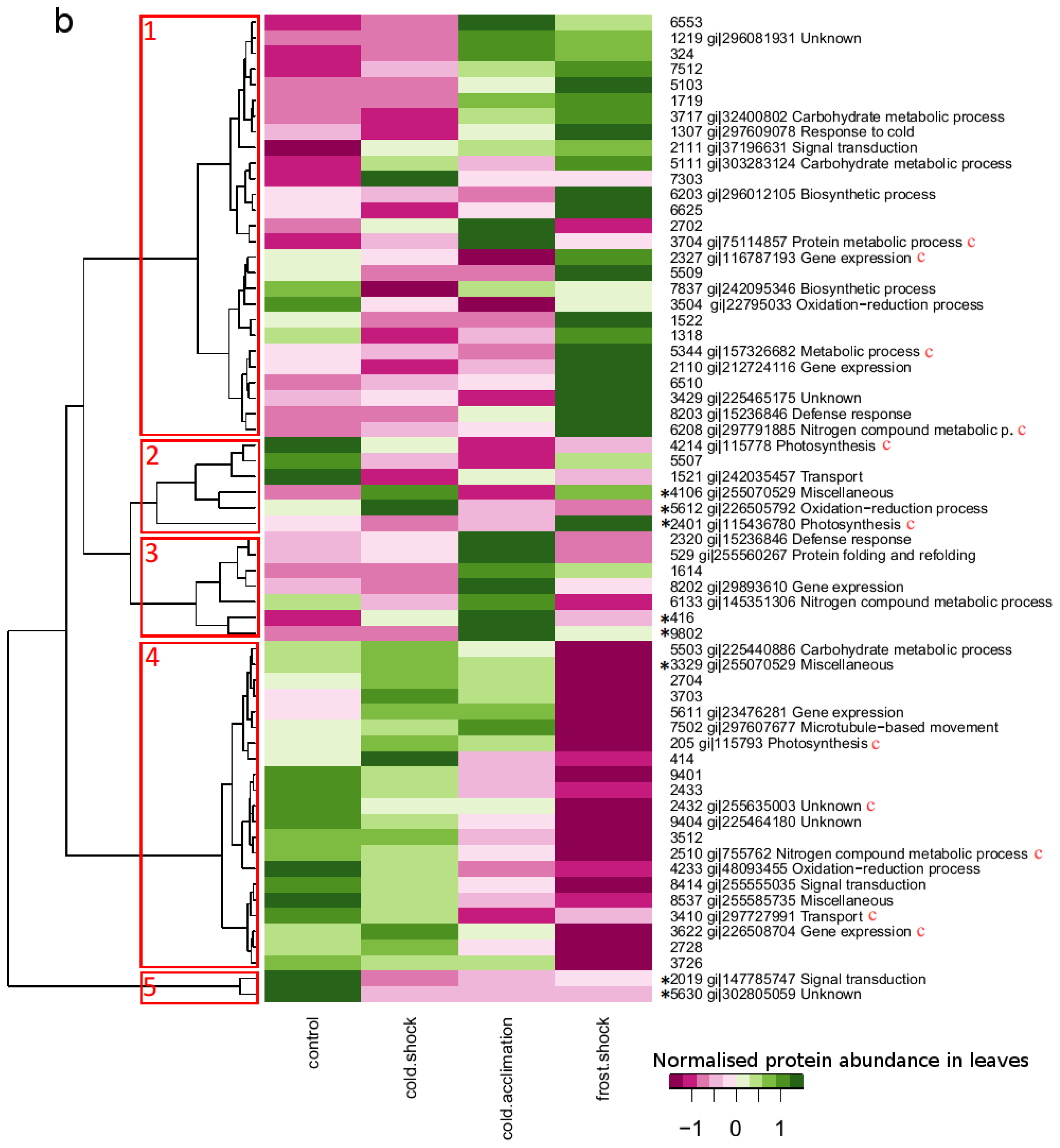
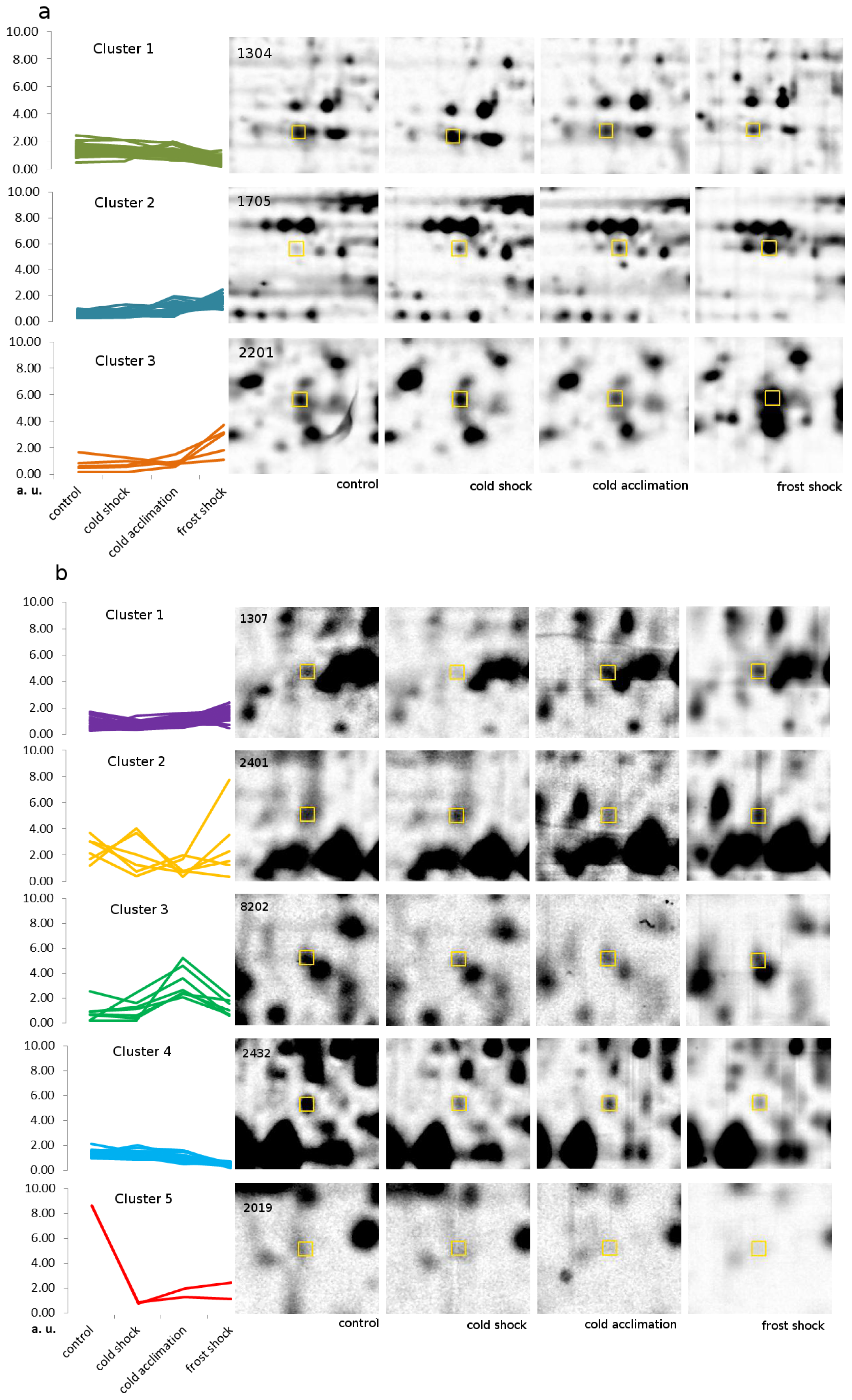
2.3. Proteins with Large Changes in Abundance
2.4. Protein Accumulation Modes Induced by Low Temperature
2.5. Importance of Chloroplasts in the Cold Hardening Process
3. Experimental Section
3.1. Plant Samples
3.2. Protein Extraction
3.3. 2D-DIGE and Image Analysis
3.4. Protein Identification
3.5. Biological Functions of Identified Proteins
3.6. Statistical Analysis of Differently Accumulated Proteins
4. Conclusions
Acknowledgments
Abbreviations
| 2D-DIGE | two dimensional difference gel electrophoresis |
| ACN | acetonitrile |
| CHAPS | 3-[(3-cholamidopropyl)dimethylammonio]-1-propanesulfonate |
| DHN | dehydrin |
| DTT | dithiothreitol |
| EDS1 | enhanced disease susceptibility 1 |
| GO | Gene Ontology |
| HSP70 | 70 kDa heat shock protein |
| IAA | iodoacetamide |
| IPG | immobilised pH gradient |
| JIP-23 | 23 kDa jasmonate-induced protein |
| LEA | late embryogenesis abundant |
| MALDI-ToF | Matrix-Assisted Laser Desorption/Ionization-Time of Flight |
| PsbO | 33 kDa oxygen evolving protein of photosystem II |
| SDS | sodium dodecyl sulfate |
| TCA | trichloroacetic acid |
| TFA | Trifluoroacetic acid |
| WCS | wheat cold-specific |
| WEGO | Web Gene Ontology Annotation Plotting |
Conflict of Interest
References
- Newton, A.C.; Flavell, A.J.; George, T.S.; Leat, P.; Mullholland, B.; Ramsay, L.; Revoredo-Giha, C.; Russell, J.; Steffenson, B.J.; Swanston, J.S.; et al. Crops that feed the world 4. Barley: A resilient crop? Strengths and weaknesses in the context of food security. Food Sec 2011, 3, 141–178. [Google Scholar]
- Limin, A.E.; Fowler, D.B. Developmental traits affecting low-temperature tolerance response in near-isogenic lines for the vernalization locus Vrn-A1 in wheat (Triticum. aestivum L. em Thell). Ann. Bot 2002, 89, 579–585. [Google Scholar]
- Hommo, L. Hardening of some winter wheat (Triticum. aestivum L.), rye (Secale. cereale L.), triticale (× Triticosecale Wittmack) and winter barley (Hordeum. vulgare L.) cultivars during autumn and the final winter survival in Finland. Plant Breed 1994, 112, 285–293. [Google Scholar]
- Kacperska, A. Plant Response to Low Temperature: Signaling Pathways Involved in Plant Acclimation. In Cold-Adapted Organisms—Ecology, Physiology, Enzymology and Molecular Biology; Margesin, R., Schinner, F., Eds.; Springer: Berlin, Germany, 1999; pp. 79–103. [Google Scholar]
- Thomashow, M.F. Plant cold acclimation: Freezing tolerance genes and regulatory mechanisms. Annu. Rev. Plant Physiol. Plant Mol. Biol 1999, 50, 571–599. [Google Scholar]
- Timperio, A.M.; Egidi, M.G.; Zolla, L. Proteomics applied on plant abiotic stresses: Role of heat shock proteins (HSP). J. Proteomics 2008, 71, 391–411. [Google Scholar]
- Kosová, K.; Vítámvás, P.; Prášil, I.T. Expression of dehydrins in wheat and barley under different temperatures. Plant Sci 2011, 180, 46–52. [Google Scholar]
- Kamal, A.H.M.; Kim, K.H.; Shin, K.H.; Choi, J.S.; Baik, B.K.; Tsujimoto, H.; Heo, H.Y.; Park, C.S.; Woo, S.H. Abiotic stress responsive proteins of wheat grain determined using proteomics technique. Aust. J. Crop. Sci 2010, 4, 196–208. [Google Scholar]
- Sarhadi, E.; Mahfoozi, S.; Hosseini, S.A.; Salekdeh, G.H. Cold acclimation proteome analysis reveals close link between the up-regulation of low-temperature associated proteins and vernalization fulfillment. J. Proteome Res 2010, 9, 5658–5667. [Google Scholar]
- Rinalducci, S.; Egidi, M.G.; Karimzadeh, G.; Jazii, F.R.; Zolla, L. Proteomic analysis of a spring wheat cultivar in response to prolonged cold stress. Electrophoresis 2011, 32, 1807–1818. [Google Scholar]
- Vítámvás, P.; Prášil, I.T.; Kosová, K.; Planchon, S.; Renaut, J. Analysis of proteome and frost tolerance in chromosome 5A and 5B reciprocal substitution lines between two winter wheats during long-term cold acclimation. Proteomics 2012, 12, 68–85. [Google Scholar]
- Fowler, D.B.; Breton, G.; Limin, A.E.; Mahfoozi, S.; Sarhan, F. Photoperiod and temperature interactions regulate low-temperature-induced gene expression in Barley. Plant Physiol 2001, 127, 1676–1681. [Google Scholar]
- Kosová, K.; Holková, L.; Prášil, I.T.; Prášilová, P.; Bradáčová, M.; Vítámvás, P.; Čapková, V. Expression of dehydrin 5 during the development of frost tolerance in barley (Hordeum. vulgare L.). J. Plant Physiol 2008, 165, 1142–1151. [Google Scholar]
- Vítámvás, P.; Saalbach, G.; Prášil, I.T.; Čapková, V.; Opatrná, J.; Ahmed, J. WCS120 protein family and proteins soluble upon boiling in cold-acclimated winter wheat. J. Plant Physiol 2007, 9, 1197–1207. [Google Scholar]
- Vítámvás, P.; Prášil, I.T. WCS120 protein family and frost tolerance during cold acclimation, deacclimation and reacclimation of winter wheat. Plant Physiol. Biochem 2008, 46, 970–976. [Google Scholar]
- Greenup, A.G.; Sasani, S.; Oliver, S.N.; Walford, S.A.; Millar, A.A.; Trevaskis, B. Transcriptome analysis of the vernalization response in barley (Hordeum. vulgare L.) seedlings. PLoS One 2011, 6, e17900. [Google Scholar]
- Svensson, J.T.; Crosatti, C.; Campoli, C.; Bassi, R.; Stanca, A.M.; Close, T.J.; Cattivelli, L. Transcriptome analysis of cold acclimation in barley Albina and Xantha Mutants. Plant Physiol 2006, 141, 257–270. [Google Scholar]
- Armbruster, U.; Pesaresi, P.; Pribil, M.; Hertle, A.; Leister, D. Update on chloroplast research: New tools, new topics and new trends. Mol. Plant 2001, 4, 1–16. [Google Scholar]
- Šimková, K.; Kim, C.; Gacek, K.; Baruah, A.; Laloi, C.; Apel, K. The chloroplast division mutant caa33 of Arabidopsis thaliana reveals the crucial impact of chloroplast homeostasis on stress acclimation and retrograde plastid-to-nucleus signaling. Plant J 2012, 69, 701–712. [Google Scholar]
- Tanino, K.K.; McKersie, B.D. Injury within the crown of winter wheat seedlings after freezing and icing stress. Can. J. Bot 1985, 63, 432–436. [Google Scholar]
- Livingston, D.P.; Premakumar, R.; Tallury, S.P. Carbohydrate partitioning between upper and lower regions of the crown in oat and rye during cold acclimation and freezing. Cryobiology 2006, 52, 200–208. [Google Scholar]
- Winfield, M.O.; Lu, C.; Wilson, I.D.; Coghill, J.A.; Edwards, K.J. Plant responses to cold: Transcriptome analysis of wheat. Plant Biotechnol. J 2010, 8, 749–771. [Google Scholar]
- Janská, A.; Aprile, A.; Zámečník, J.; Cattivelli, L.; Ovesná, J. Transcriptional responses of winter barley to cold indicate nucleosome remodelling as a specific feature of crown tissues. Funct. Integr. Genomics 2011, 11, 307–325. [Google Scholar]
- Renaut, J.; Lutts, S.; Hoffmann, L.; Hausman, J.F. Responses of poplar to chilling temperatures: Proteomic and physiological aspects. Plant Biol 2004, 6, 81–90. [Google Scholar]
- Hashimoto, M.; Komatsu, S. Proteomic analysis of rice seedlings during cold stress. Proteomics 2007, 7, 1293–302. [Google Scholar]
- Ganeshan, S.; Vítámvás, P.; Fowler, D.B.; Chibbar, R.N. Quantitative expression analysis of selected COR genes reveals their differential expression in leaf and crown tissues of wheat (Triticum aestivum L.) during an extended low temperature acclimation regimen. J. Exp. Bot 2008, 59, 2393–2402. [Google Scholar]
- Limin, A.E.; Corey, A.; Hayes, P.; Fowler, D.B. Low-Temperature acclimation of barley cultivars used as parents in mapping populations: Response to photoperiod, vernalization and phenological development. Planta 2007, 226, 139–146. [Google Scholar]
- Kjellsen, T.D.; Shiryaeva, L.; Schröder, W.P.; Strimbeck, G.R. Proteomics of extreme freezing tolerance in Siberian spruce (Picea. obovata). J. Proteomics 2010, 73, 965–975. [Google Scholar]
- Ifuku, K.; Ishihara, S.; Sato, F. Molecular functions of oxygen-evolving complex family proteins in photosynthetic electron flow. J. Integr. Plant Biol 2010, 52, 723–734. [Google Scholar]
- Cui, S.; Huang, F.; Wang, J.; Ma, X.; Cheng, Y.; Liu, J. A proteomic analysis of cold stress responses in rice seedlings. Proteomics 2005, 5, 3162–3172. [Google Scholar]
- Schweighofer, A.; Hirt, H.; Meskiene, I. Plant PP2C phosphatases: Emerging functions in stress signaling. Trends Plant Sci 2004, 9, 236–243. [Google Scholar]
- Tahtiharju, S.; Palva, T. Antisense inhibition of protein phosphatase 2C accelerates cold acclimation in Arabidopsis thaliana. Plant J 2001, 26, 461–470. [Google Scholar]
- Frydman, J. Folding of newly translated proteins in vivo: The role of molecular chaperones. Annu. Rev. Biochem 2001, 70, 603–647. [Google Scholar]
- Straus, M.R.; Rietz, S.; ver Loren van Themaat, E.; Bartsch, M.; Parker, J.E. Salicylic acid antagonism of EDS1-driven cell death is important for immune and oxidative stress responses in Arabidopsis. Plant J 2010, 62, 628–640. [Google Scholar]
- Vítámvás, P.; Kosová, K.; Prášilová, P.; Prášil, I.T. Accumulation of WCS120 protein in wheat cultivars grown at 9 C or 17 C in relation to their winter survival. Plant Breed 2010, 129, 611–616. [Google Scholar]
- Oikawa, A.; Nagai, K.; Kato, K.; Kidou, S. Gene silencing of barley P23k involved in secondary wall formation causes abnormal tiller formation and intercalary elongation. Breed. Sci 2009, 59, 664–670. [Google Scholar]
- Lehmann, J.; Atzorn, R.; Bruckner, C.; Reinbothe, S.; Leopold, J.; Wasternack, C.; Parthier, B. Accumulation of jasmonate, abscisic-acid, specific transcripts and proteins in osmotically stressed barley leaf segmants. Planta 1995, 197, 156–162. [Google Scholar]
- Dunn, M.A.; Morris, A.; Jack, P.L.; Hughes, M.A. A low-temperature-responsive translation elongation factor lα from barley (Hordeum. vulgare L.). Plant Mol. Biol 1993, 23, 221–225. [Google Scholar]
- Wallsgrove, R.M.; Turner, J.C.; Hall, N.P.; Kendall, A.C.; Bright, S.W. Barley mutants lacking chloroplast glutamine synthetase-biochemical and genetic analysis. Plant Physiol 1987, 83, 155–158. [Google Scholar]
- Pang, Q.Y.; Chen, S.X.; Dai, S.J.; Chen, Y.Z.; Wang, Y.; Yan, X.F. Comparative proteomics of salt tolerance in Arabidopsis thaliana and Thellungiella. halophila. J. Proteome Res 2010, 9, 2584–2599. [Google Scholar]
- Yan, S.P.; Tang, Z.C.; Su, W.; Sun, W.N. Proteomic analysis of salt stress-responsive proteins in rice root. Proteomics 2005, 5, 235–244. [Google Scholar]
- Cai, H.; Zhou, Y.; Xiao, J.; Li, X.; Zhang, Q.; Lian, X. Overexpressed glutamine synthetase gene modifies nitrogen metabolism and abiotic stress responses in rice. Plant Cell Rep 2009, 28, 527–537. [Google Scholar]
- Skinner, D.Z. Post-Acclimation transcriptome adjustment is a major factor in freezing tolerance of winter wheat. Funct. Integr. Genomics 2009, 9, 513–523. [Google Scholar]
- Hanano, A.; Burcklen, M.; Flenet, M.; Ivancich, A.; Louwagie, M.; Garin, J.; Blee, E. Plant seed peroxygenase is an original heme-oxygenase with an EF-hand calcium binding motif. J. Biol. Chem 2006, 281, 33140–33151. [Google Scholar]
- Thangasamy, S.; Chen, P.W.; Lai, M.H.; Chen, J.; Jauh, G.Y. Rice LGD1 containing RNA binding activity affects growth and development through alternative promoters. Plant J 2012, 71, 288–302. [Google Scholar]
- Liu, X.D.; Shen, Y.G. NaCl-Induced phosphorylation of light harvesting chlorophyll a/b proteins in thylakoid membranes from the halotolerant green alga, Dunaliella. salina. FEBS Lett 2004, 569, 337–340. [Google Scholar]
- Wang, W.; Tai, F.J.; Chen, S.N. Optimizing protein extraction from plant tissues for enhanced proteomics analysis. J. Sep. Sci 2008, 31, 2032–2039. [Google Scholar]
- Strohalm, M.; Kavan, D.; Novak, P.; Volny, M.; Havlicek, V. mMass 3: A Cross–Platform software environment for precise analysis of mass spectrometric data. Anal. Chem 2010, 11, 4648–4651. [Google Scholar]
- Barley1.na32.annot.csv. Available online: http://www.affymetrix.com/estore/ (accessed on 3 February 2013).
- UniProtKB. Available online: http://www.uniprot.org/ (accessed on 3 February 2013).
- The Gene Ontology Consortium. Gene ontology: Tool for the unification of biology. Nat. Genet 2000, 1, 25–29.
- Emanuelsson, O.; Nielsen, H.; Brunak, S.; von Heijne, G. Predicting subcellular localization of proteins based on their N-terminal amino acid sequence. J. Mol. Biol 2000, 300, 1005–1016. [Google Scholar]
- Ye, J.; Fang, L.; Zheng, H.K.; Zhang, Y.; Chen, J.; Zhang, Z.J.; Wang, J.; Li, S.T.; Li, R.Q.; Bolund, L.; et al. WEGO: A web tool for plotting GO annotations. Nucleic Acids Res 2006, 34, W293–W297. [Google Scholar]
- WEGO. Available online: http://wego.genomics.org.cn/cgi-bin/wego/index.pl (accessed on 1 February 2013).
- R 2.15.0. Available online: http://www.r-project.org/ (accessed on 1 October 2012).
- Grubbs, F.E. Procedures for detecting outlying observations in samples. Technometrics 1969, 11, 1–21. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot No 1 | NCBI identifier (Probe Set ID) 2 | Protein name (organism) | Score 3 | MWo/pIo (MWt/pIt) 4 | SC (%)/MP 5 | Biological function 6 | Cluster 7 | CL, O 8 |
|---|---|---|---|---|---|---|---|---|
| Crowns | ||||||||
| 2814 | gi|11527563 (Contig2786_at) | vacuolar proton-ATPase (Hordeum vulgare) | 187 | 136/5.4 (69/5.2) | 37/20 | ATP m. p. | 1 | |
| 8408 | gi|226316443 | fructose-bisphosphate aldolase (Hordeum vulgare) | 120 | 70/6.7 (39/6.1) | 36/11 | Carbohydrate m. p. | 1 | |
| 3404 | gi|326490940 | malate dehydrogenase (Hordeum vulgare) | 83 | 69/5.5 (36/5.6) | 25/10 | Carbohydrate m. p. | 1 | |
| 1304 | gi|326513418 | fructokinase-2 (Hordeum vulgare) | 139 | 62/5.2 (36/6.1) | 39/17 | Carbohydrate m. p. | 1 | |
| 1311 | gi|326513418 | fructokinase-2 (Hordeum vulgare) | 179 | 62/5.1 (36/6.1) | 39/17 | Carbohydrate m. p. | 1 | |
| 4613 | gi|357110692 | 6-phosphogluconate dehydrogenase, decarboxylating-like 1 (Brachypodium distachyon) | 80 | 94/5.9 (53/5.6) | 22/12 | Carbohydrate m. p. | 1 | |
| 3705 | gi|326493636 | enolase (Hordeum vulgare) | 188 | 107/5.6 (48/5.4) | 32/11 | Carbohydrate m. p. | 1 | |
| 2612 | gi|326495636 | actin (Hordeum vulgare) | 225 | 89/5.4 (42/5.3) | 47/21 | Cytoskeleton organization | 1 | |
| 108 | gi|125548641 | h. p. OsI_16233 (Oryza sativa) | 85 | 22/4.6 (17/4.8) | 47/9 | Defence response | 1 | * |
| 2506 | gi|242074406 | h. p. SORBIDRAFT_06g029250 (Sorghum bicolor) | 84 | 79/5.3 (183/6.0) | 11/15 | Defence response | 1 | |
| 12 | gi|37651975 | chopper chaperone (Hordeum vulgare) | 75 | 20/4.6 (11/4.7) | 41/6 | Metal ion transport | 1 | |
| 4418 | gi|40317416 | glutamine synthetase GSr1 (Triticum aestivum) | 114 | 69/5.8 (39/5.4) | 34/11 | Nitrogen compound m. p. | 1 | |
| 6806 | gi|755762 (HVSMEa0004F18f2_s_at) | glutamine synthetase GS2 (Hordeum vulgare) | 150 | 137/6.2 (47/5.8) | 19/12 | Nitrogen compound m. p. | 1 | c |
| 1713 | gi|326496957 | thioredoxin peroxidase (Hordeum vulgare) | 118 | 113/5.1 (28/6.3) | 42/8 | Oxidation-reduction p. | 1 | c |
| 6503 | gi|326492375 | predicted protein (Hordeum vulgare) | 77 | 85/6.2 (41/5.9) | 17/7 | Oxidation-reduction p. | 1 | |
| 5611 | gi|326493350 | isocitrate dehydrogenase (Hordeum vulgare) | 87 | 89/6.7 (46/6.0) | 20/8 | Oxidation-reduction p. | 1 | |
| 7414 | gi|326516152 | predicted protein (Hordeum vulgare) | 122 | 71/6.6 (39/6.4) | 32/14 | Oxidation-reduction p. | 1 | |
| 5716 | gi|194707130 | unknown (Zea mays) | 95 | 104/6.1 (40/8.5) | 26/10 | Protein catabolic process | 1 | |
| 2901 | gi|326533328 | 60 kDa chaperonin (Hordeum vulgare) | 98 | 141/5.3 (62/5.4) | 16/10 | Protein folding/refolding | 1 | c |
| 221 | gi|113603 (Contig86_at) | thiol protease aleurain (Hordeum vulgare) | 76 | 45/4.8 (40/6.4) | 20/7 | Protein m. p. | 1 | |
| 4714 | gi|297791831 | zinc carboxypeptidase family protein (Arabidopsis lyrata) | 79 | 100/5.8 (53/6.9) | 22/12 | Protein m. p. | 1 | |
| 2008 | gi|326493798 | glycine rich protein, RNA binding protein (Hordeum vulgare) | 139 | 18/5.4 (17/5.4) | 50/10 | Response to cold | 1 | |
| 4304 | gi|326494746 | predicted protein (Hordeum vulgare) | 148 | 63/5.7 (38/5.7) | 37/12 | Seed storage protein | 1 | |
| 3601 | gi|357149925 | elongation factor Tu (Brachypodium distachyon) | 148 | 93/5.5 (51/5.9) | 40/23 | Translational elongation | 1 | c |
| 5506 | gi|326514754 | elongation factor Tu (Hordeum vulgare) | 124 | 80/6.0 (48/6.0) | 48/18 | Translational elongation | 1 | |
| 4614 | gi|125580018 | h. p. OsJ_36811 (Oryza sativa) | 74 | 95/5.9 (32/6.8) | 42/10 | Unknown | 1 | |
| 6303 | gi|326491885 | predicted protein (Hordeum vulgare) | 158 | 61/6.2 (37/6.0) | 39/14 | Unknown | 1 | |
| 8303 | gi|326497973 | predicted protein (Hordeum vulgare) | 153 | 61/6.6 (39/6.3) | 55/15 | Unknown | 1 | |
| 2808 | gi|326505132 | predicted protein (Hordeum vulgare) | 105 | 123/5.3 (56/4.7) | 18/12 | Unknown | 1 | |
| 4414 | gi|326517467 | predicted protein (Hordeum vulgare) | 142 | 65/5.9 (36/5.7) | 51/16 | Unknown | 1 | |
| 7717 | gi|326517784 | predicted protein (Hordeum vulgare) | 105 | 104/6.6 (47/6.1) | 32/14 | Unknown | 1 | |
| 5416 | gi|356532429 | LOC100792980 (Glycine max) | 86 | 74/6.1 (75/7.2) | 18/11 | Unknown | 1 | |
| 5111 | gi|400094 (Contig1675_s_at) | JIP-23 (Hordeum vulgare) | 84 | 32/6.5 (23/5.9) | 38/8 | Unknown | 1 | |
| 2414 | gi|61889374 (Contig112_at) | C2 domain-containing protein (Hordeum vulgare) | 86 | 66/5.4 (33/5.6) | 21/7 | Unknown | 1 | |
| 7012 | gi|326493636 | enolase (Hordeum vulgare) | 79 | 15/6.6 (48/5.4) | 32/11 | Carbohydrate m. p. | 2 | |
| 1402 | gi|149939605 | enhanced disease susceptibility 1 (Arabidopsis lyrata) | 75 | 73/5.0 (70/6.1) | 15/11 | Defence response | 2 | * |
| 4110 | gi|242074406 | h. p. SORBIDRAFT_06g029250 (Sorghum bicolor) | 88 | 36/5.9 (183/6.0) | 9/12 | Defence response | 2 | * |
| 5202 | gi|146760217 (Contig1508_s_at) | beta tubulin 6 (Hordeum vulgare) | 132 | 41/5.9 (51/4.8) | 26/16 | Microtubule-based mov. | 2 | |
| 5521 | gi|3915083 | beta tubulin (Cicer arietinum) | 74 | 78/6.1 (51/4.8) | 26/12 | Microtubule-based mov. | 2 | |
| 704 | gi|357113738 | 40S ribosomal protein SA-like (Brachypodium distachyon) | 185 | 108/4.9 (33/4.9) | 44/16 | Proteosynthesis | 2 | |
| 226 | gi|326526175 | predicted protein (Hordeum vulgare) | 82 | 37/4.9 (26/6.2) | 33/7 | Response to cold | 2 | c |
| 2001 | gi|326506132 | 70 kDa heat shock protein (Hordeum vulgare) | 257 | 15/5.2 (74/5.0) | 12/8 | Response to cold | 2 | |
| 1705 | gi|326492960 | 70 kDa heat shock protein (Hordeum vulgare) | 82 | 102/5.1 (74/5.0) | 12/8 | Response to cold | 2 | c,* |
| 2108 | gi|326504940 | predicted protein (Hordeum vulgare) | 84 | 35/5.3 (26/6.2) | 33/7 | Response to cold | 2 | |
| 817 | gi|326497759 | predicted protein (Hordeum vulgare) | 86 | 120/4.9 (42/6.0) | 28/11 | Unknown | 2 | |
| 5503 | gi|326497759 | predicted protein (Hordeum vulgare) | 87 | 77/5.9 (42/6.0) | 20/9 | Unknown | 2 | |
| 2201 | gi|326494312 | predicted protein (Hordeum vulgare) | 160 | 39/5.3 (26/6.2) | 33/7 | Protein folding | 3 | |
| 1004 | gi|326505132 | predicted protein (Hordeum vulgare) | 175 | 15/5.4 (39/6.4) | 32/14 | Unknown | 3 | * |
| Spot No 1 | NCBI identifier (Probe Set ID) 2 | Protein name (organism) | Score 3 | MWo/pIo (MWt/pIt) 4 | SC (%)/MP 5 | Biological function 6 | Cluster 7 | CL, O 8 |
|---|---|---|---|---|---|---|---|---|
| Leaves | ||||||||
| 6203 | gi|296012105 | granule-bound starch synthase I (Solanum caripense) | 73 | 90/5.8 (37/6.7) | 33/9 | Biosynthetic process | 1 | |
| 3717 | gi|32400802 | phosphoglycerate mutase (Triticum aestivum) | 78 | 114/5.3 (30/5.4) | 28/6 | Carbohydrate m. p. | 1 | |
| 5111 | gi|303283124 | pyruvate kinase (Micromonas pusilla CCMP1545) | 76 | 48/5.5 (58/6.0) | 21/11 | Carbohydrate m. p. | 1 | |
| 7837 | gi|242095346 | h. p. SORBIDRAFT_10g009020 (Sorghum bicolor) | 73 | 101/6.0 (68/8.8) | 14/9 | Cell wall biogenesis | 1 | |
| 5344 | gi|157326682 | 1(10),5-germacradien-4-ol synthase (Pinus sylvestris) | 76 | 34/5.7 (73/5.7) | 17/11 | Metabolic process | 1 | c |
| 6208 | gi|297791885 | adenylate kinase (Arabidopsis lyrata) | 73 | 90/5.8 (33/9.1) | 22/6 | Nucleobase-containing compound m. p. | 1 | c |
| 3504 | gi|22795033 | putative cytochrome P450 (Populus tremula) | 96 | 70/5.3 (24/7.1) | 31/7 | Oxidation-reduction p. | 1 | |
| 2327 | gi|116787193 | unknown (Picea sitchensis) | 77 | 43/5.2 (33/9.3) | 27/9 | Protein m. p. | 1 | c |
| 3704 | gi|75114857 | ATP-dependent zinc metalloprotease FTSH 2 (Oryza sativa Japonica) | 73 | 89/5.4 (73/5.5) | 15/8 | Protein m. p. | 1 | c |
| 3429 | gi|225465175 | 60S ribosomal protein L3-like (Vitis vinifera) | 82 | 59/5.4 (25/10.0) | 45/9 | Proteosynthesis | 1 | |
| 1307 | gi|297609078 | Os09g0133600 (Oryza sativa Japonica Group) | 78 | 52/5.0 (27/5.1) | 28/6 | Response to cold | 1 | |
| 8203 | gi|15236846 | peroxygenase 1 (Arabidopsis thaliana) | 82 | 49/6.3 (28/5.8) | 34/12 | Response to freezing | 1 | |
| 2110 | gi|212724116 | uncharacterised protein (Zea mays) | 74 | 116/5.2 (42/9.1) | 18/8 | tRNA processing | 1 | |
| 2111 | gi|37196631 | resistance protein candidate (Helianthus annuus) | 75 | 19/5.1 (18/5.1) | 31/7 | Unknown | 1 | |
| 1219 | gi|296081931 | unnamed protein product (Vitis vinifera) | 74 | 25/4.9 (242/6.5) | 5/11 | Unknown | 1 | |
| 4106 | gi|255070529 | AAA ATPase (Micromonas sp. RCC299) | 90 | 117/5.4 (84/8.0) | 21/14 | Diverse cellular processes | 2 | * |
| 5612 | gi|226505792 | UDP-glucose 6-dehydrogenase (Zea mays) | 73 | 85/5.7 (53/6.1) | 23/10 | Oxidation-reduction p. | 2 | * |
| 4214 | gi|115778 | chlorophyll a-b binding protein 1 (Sinapis alba) | 73 | 128/5.5 (24/5.2) | 27/7 | Photosynthesis | 2 | c |
| 2401 | gi|115436780 | 33kDa oxygen evolving protein of photosystem II (Oryza sativa Japonica Group) | 79 | 53/5.2 (35/6.1) | 23/5 | Photosynthesis | 2 | c,* |
| 1521 | gi|242035457 | Sb01g032420 (Sorghum bicolor) | 74 | 59/5.2 (80/5.6) | 12/10 | Transport | 2 | |
| 6133 | gi|145351306 | F-ATPase (Ostreococcus lucimarinus) | 75 | 76/5.8 (68/5.0) | 17/9 | ATP m. p. | 3 | |
| 8202 | gi|29893610 | SR-rich pre-mRNA splicing activator (Oryza sativa) | 75 | 143/6.0 (23/5.9) | 36/7 | mRNA processing | 3 | |
| 529 | gi|255560267 | chaperonin 60 kDa (Ricinus communis) | 96 | 91/4.7 (61/6.2) | 25/14 | Protein folding/refolding | 3 | |
| 2320 | gi|15236846 | peroxygenase 1 (Arabidopsis thaliana) | 83 | 39/5.2 (28/5.8) | 30/8 | Response to freezing | 3 | |
| 3329 | gi|255070529 | AAA ATPase (Micromonas sp. RCC299) | 74 | 112/5.6 (84/8.0) | 12/8 | Diverse cellular processes | 4 | * |
| 8414 | gi|255555035 | ATP binding protein (Ricinus communis) | 74 | 81/6.2 (75/5.8) | 21/10 | MAPK cascade | 4 | |
| 7502 | gi|297607677 | kinesin (Oryza sativa) | 75 | 76/6.0 (41/5.4) | 33/14 | Microtubule-based mov. | 4 | |
| 2510 | gi|755762 (HVSMEa0004F18f2_s_at) | glutamine synthetase GS2 (Hordeum vulgare) | 101 | 66/5.2 (47/5.8) | 46/10 | Nitrogen compound m. p. | 4 | c |
| 4233 | gi|48093455 | ADH-like UDP-glucose dehydrogenase (Nicotiana tabacum) | 75 | 36/5.5 (42/6.2) | 24/10 | Oxidation-reduction p. | 4 | |
| 205 | gi|115793 (Contig636_at) | chlorophyll a-b binding protein of LHCII type III (Hordeum vulgare) | 73 | 43/4.8 (29/5.0) | 26/6 | Photosynthesis | 4 | c |
| 3410 | gi|297727991 | Os11g0153600 (Oryza sativa) | 73 | 113/5.2 (27/10.8) | 41/7 | GTP catabolic process | 4 | c |
| 3622 | gi|226508704 | elongation factor Tu (Zea mays) | 101 | 75/5.3 (51/6.1) | 33/15 | Translational elongation | 4 | c |
| 5611 | gi|23476281 | myb-like transcription factor 1 (Gossypium raimondii) | 75 | 59/5.7 (31/7.1) | 19/6 | Transription | 4 | |
| 2432 | gi|255635003 | unknown (Glycine max) | 73 | 60/5.1 (34/6.4) | 21/6 | Unknown | 4 | c |
| 5503 | gi|225440886 | h. p. (Vitis vinifera) | 74 | 42/5.8 (18/6.0) | 49/8 | Unknown | 4 | |
| 8537 | gi|255585735 | calmodulin binding protein (Ricinus communis) | 75 | 78/6.2 (54/8.9) | 26/13 | Unknown | 4 | |
| 9404 | gi|225464180 | h. p. (Vitis vinifera) | 74 | 46/6.5 (37/6.1) | 25/7 | Unknown | 4 | |
| 2019 | gi|147785747 | h. p. VITISV_033291 (Vitis vinifera) | 81 | 112/4.9 (25/5.3) | 38/8 | Protein dephosphorylation | 5 | * |
| 5630 | gi|302805059 | h. p. SELMODRAFT_423400 (Selaginella moellendorffii) | 73 | 83/5.7 (48/8.7) | 29/12 | Unknown | 5 | * |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hlaváčková, I.; Vítámvás, P.; Šantrůček, J.; Kosová, K.; Zelenková, S.; Prášil, I.T.; Ovesná, J.; Hynek, R.; Kodíček, M. Proteins Involved in Distinct Phases of Cold Hardening Process in Frost Resistant Winter Barley (Hordeum vulgare L.) cv Luxor. Int. J. Mol. Sci. 2013, 14, 8000-8024. https://doi.org/10.3390/ijms14048000
Hlaváčková I, Vítámvás P, Šantrůček J, Kosová K, Zelenková S, Prášil IT, Ovesná J, Hynek R, Kodíček M. Proteins Involved in Distinct Phases of Cold Hardening Process in Frost Resistant Winter Barley (Hordeum vulgare L.) cv Luxor. International Journal of Molecular Sciences. 2013; 14(4):8000-8024. https://doi.org/10.3390/ijms14048000
Chicago/Turabian StyleHlaváčková, Iva, Pavel Vítámvás, Jiří Šantrůček, Klára Kosová, Sylva Zelenková, Ilja Tom Prášil, Jaroslava Ovesná, Radovan Hynek, and Milan Kodíček. 2013. "Proteins Involved in Distinct Phases of Cold Hardening Process in Frost Resistant Winter Barley (Hordeum vulgare L.) cv Luxor" International Journal of Molecular Sciences 14, no. 4: 8000-8024. https://doi.org/10.3390/ijms14048000