Enhanced Inhibition of Prostate Tumor Growth by Dual Targeting the Androgen Receptor and the Regulatory Subunit Type Iα of Protein Kinase A in Vivo

 and
and 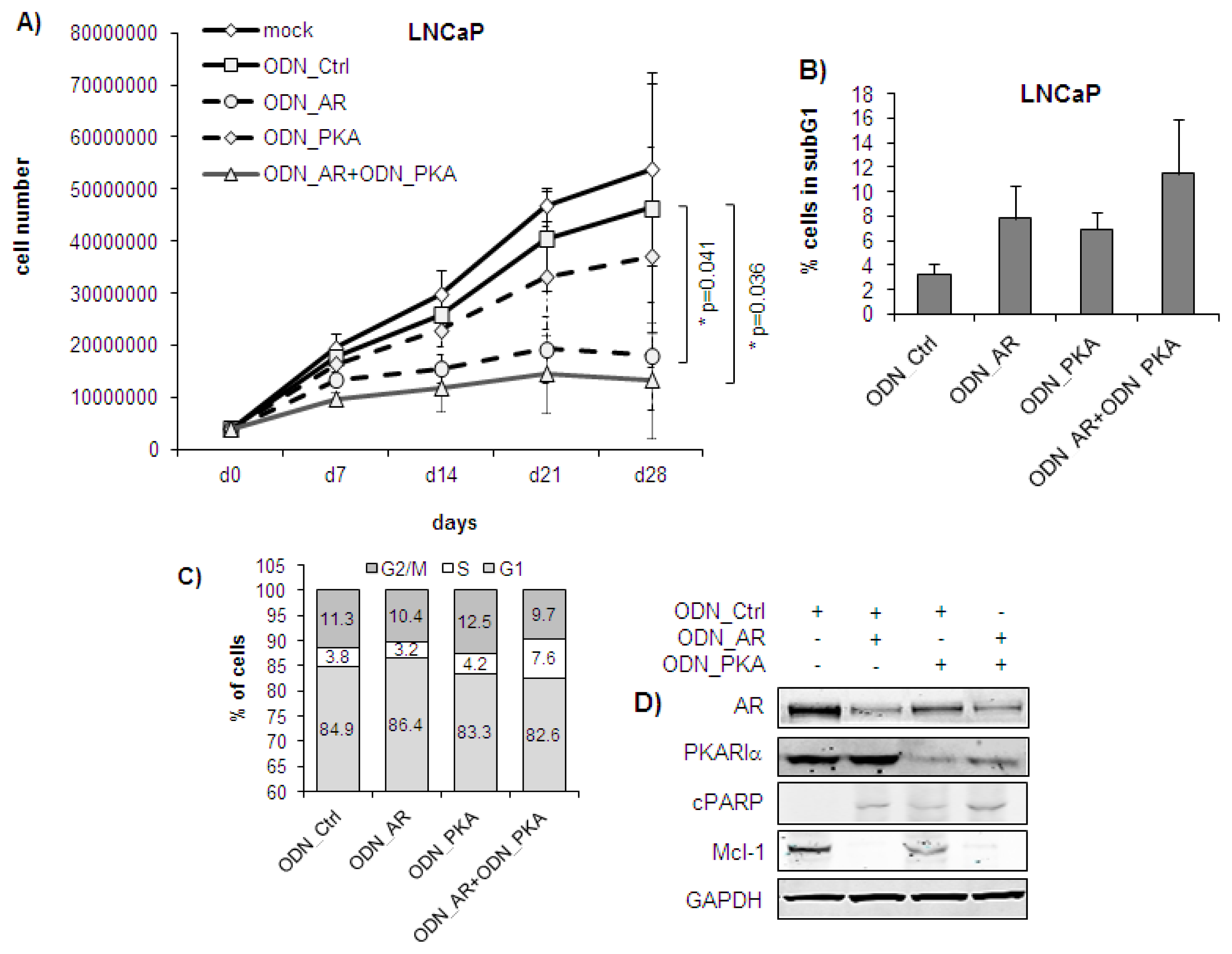{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
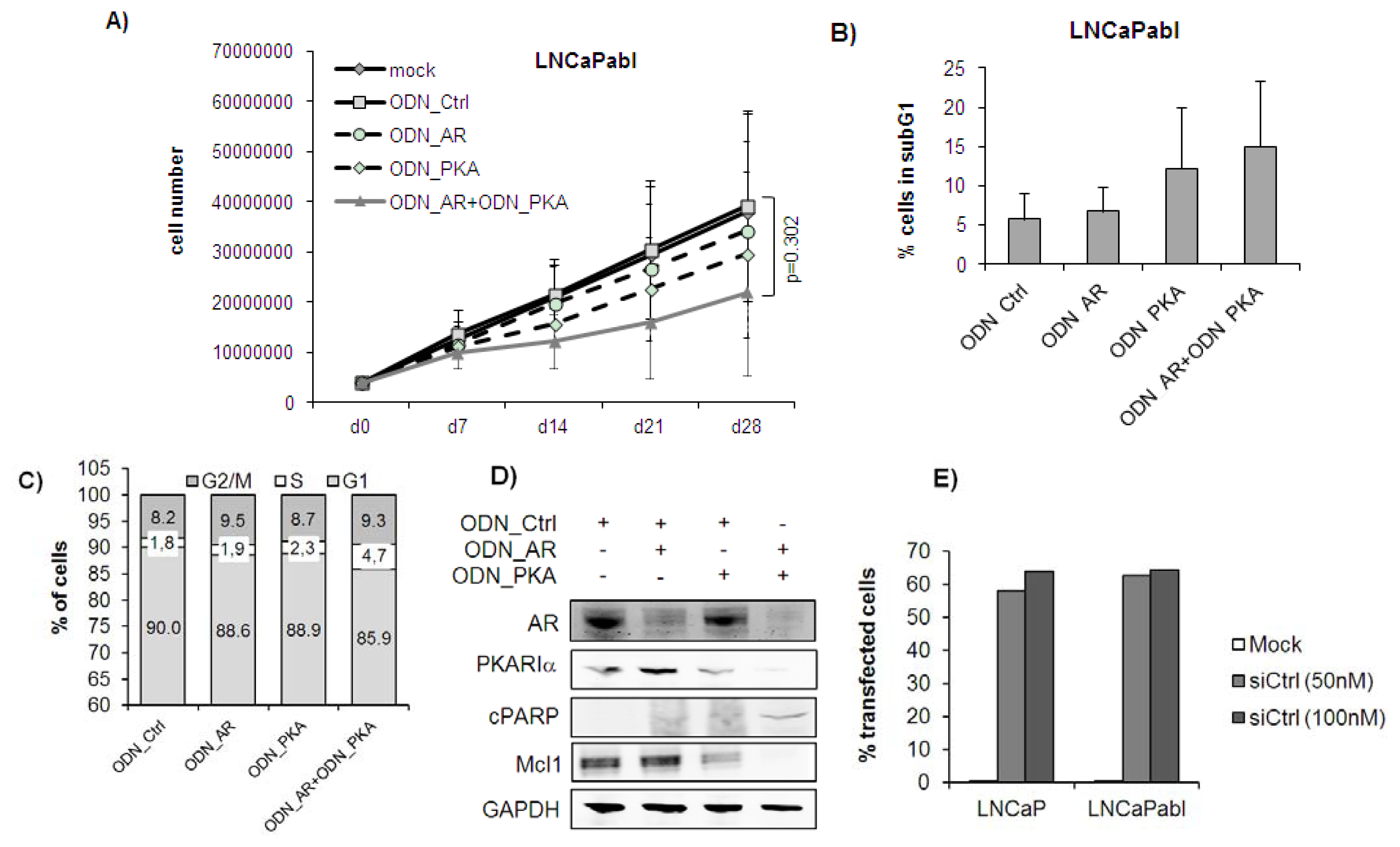
2.1. Single or Dual Targeting of AR and PKARIα with Second Generation ODNs Inhibits Prostate Cancer Cell Growth in Vitro by Induction of Apoptosis
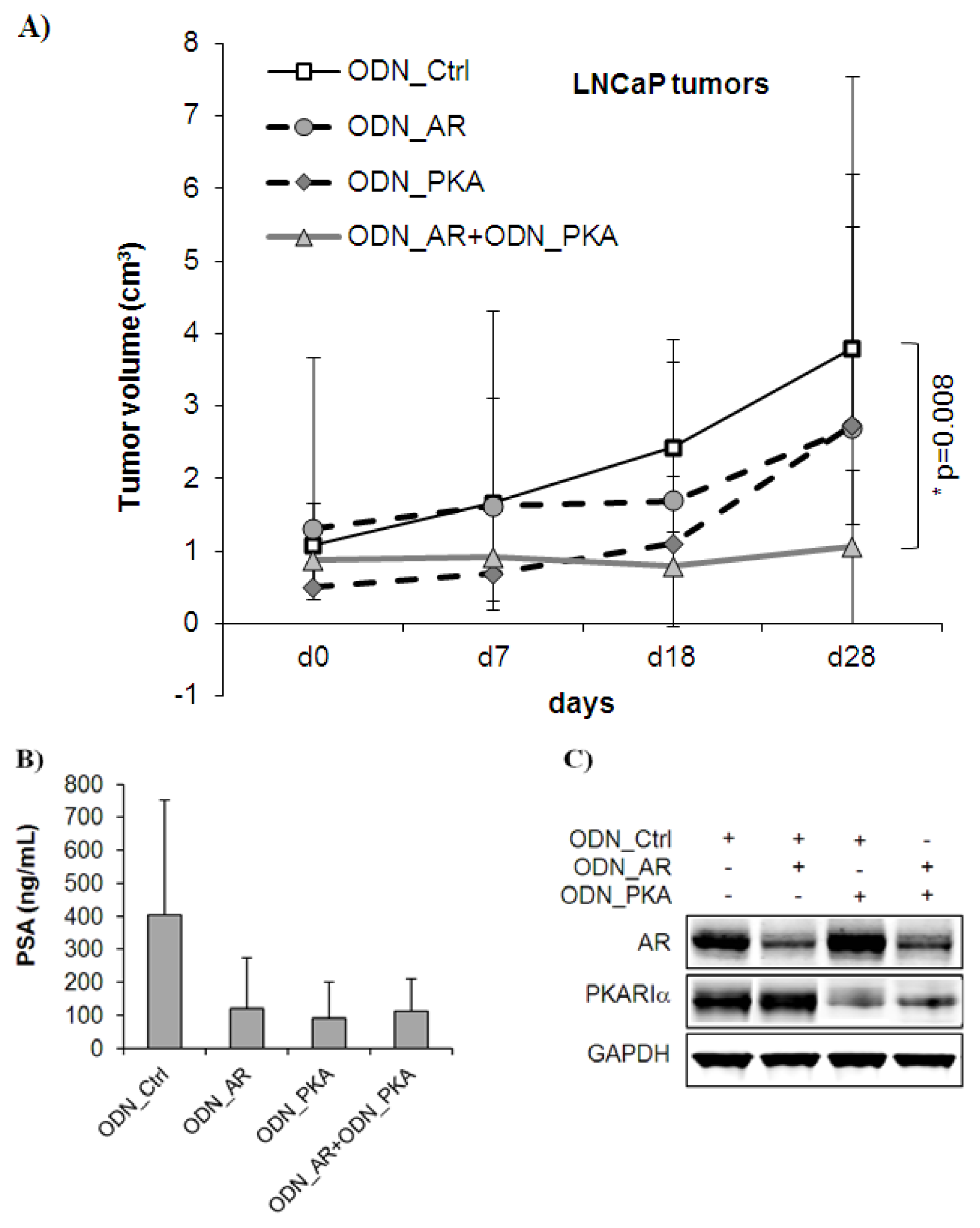
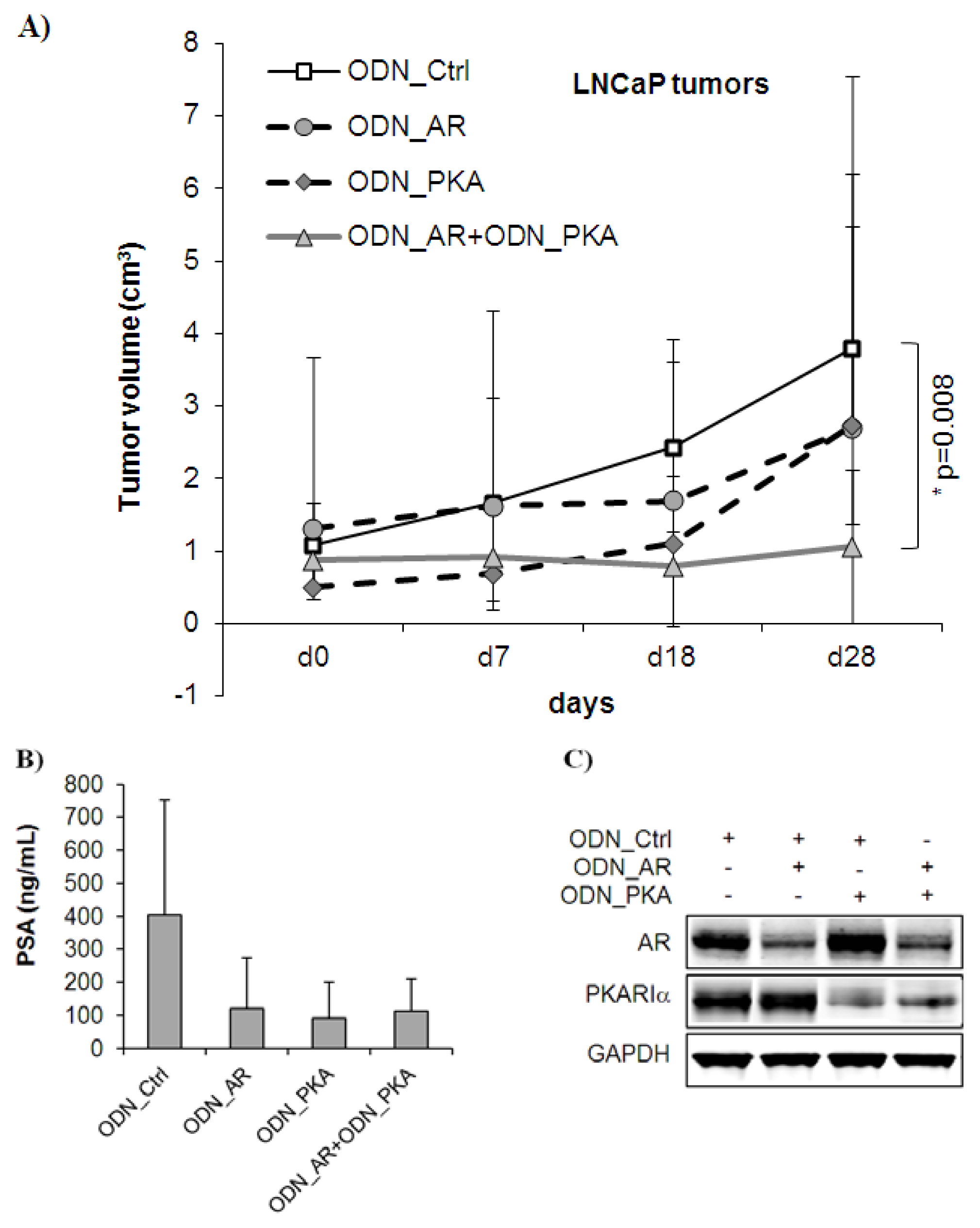
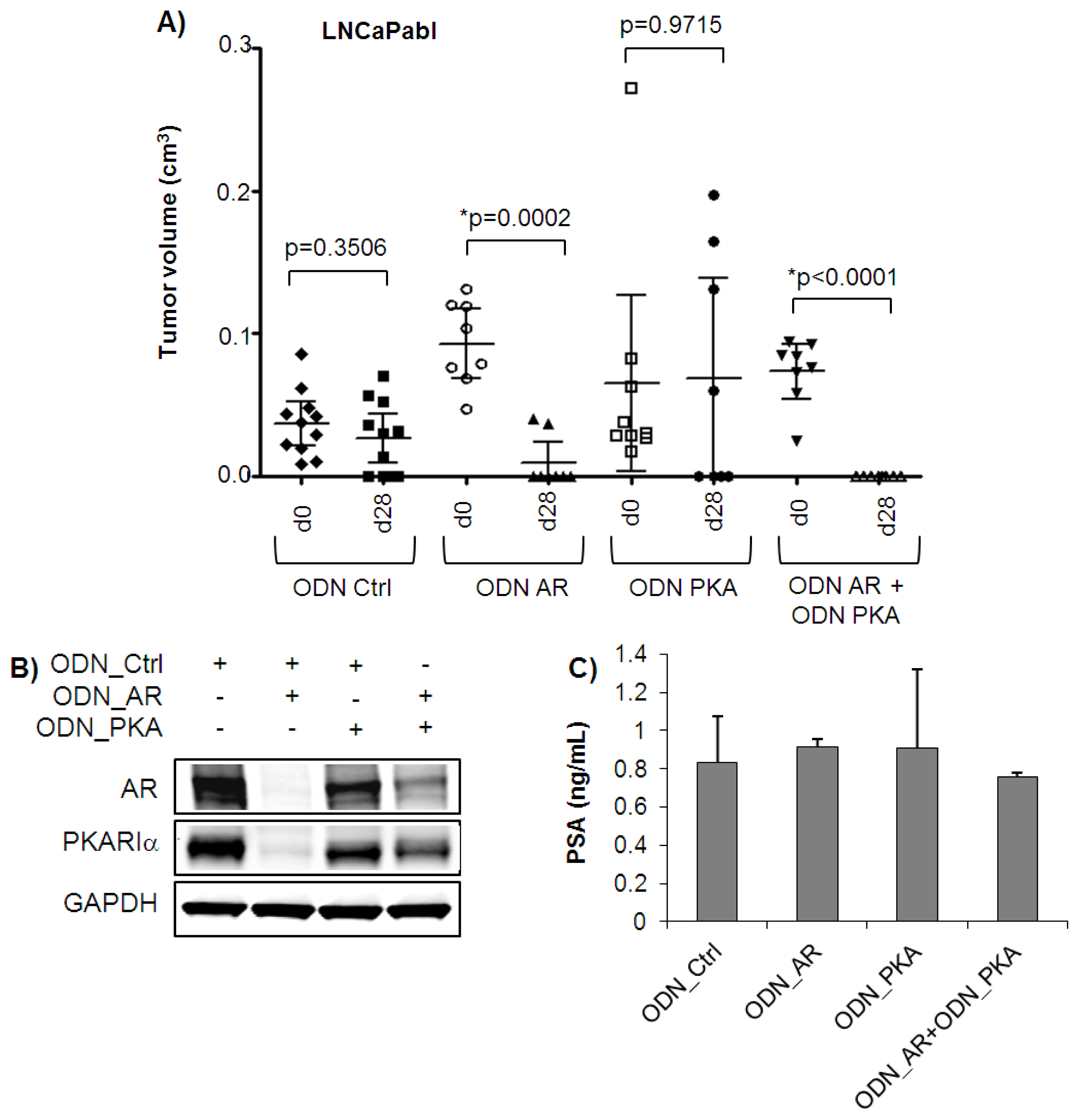
2.2. Combined Targeting of AR and PKARIα Results in Improved Growth Inhibition over Single Targeting in Androgen Sensitive LNCaP Tumors in Vivo
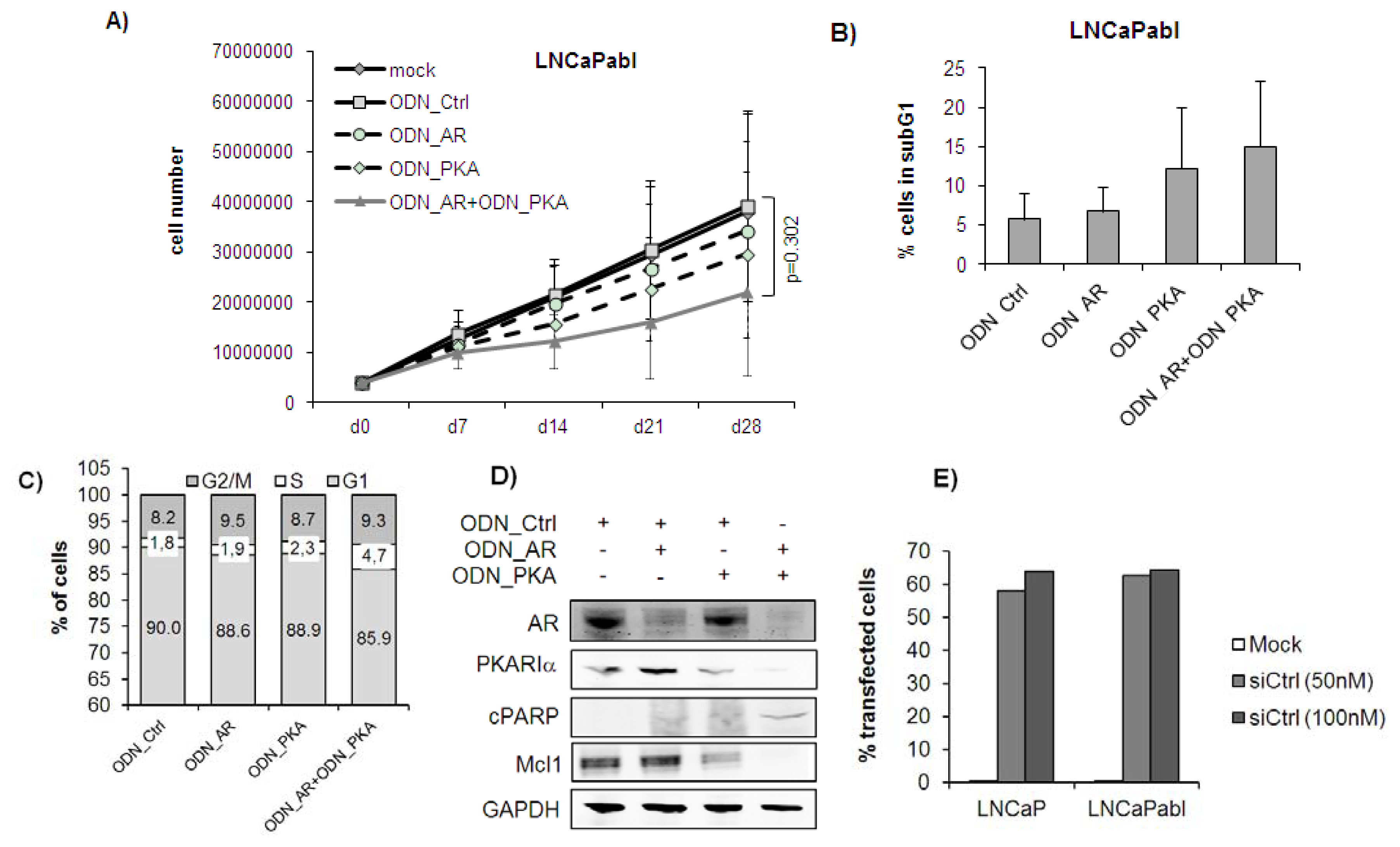
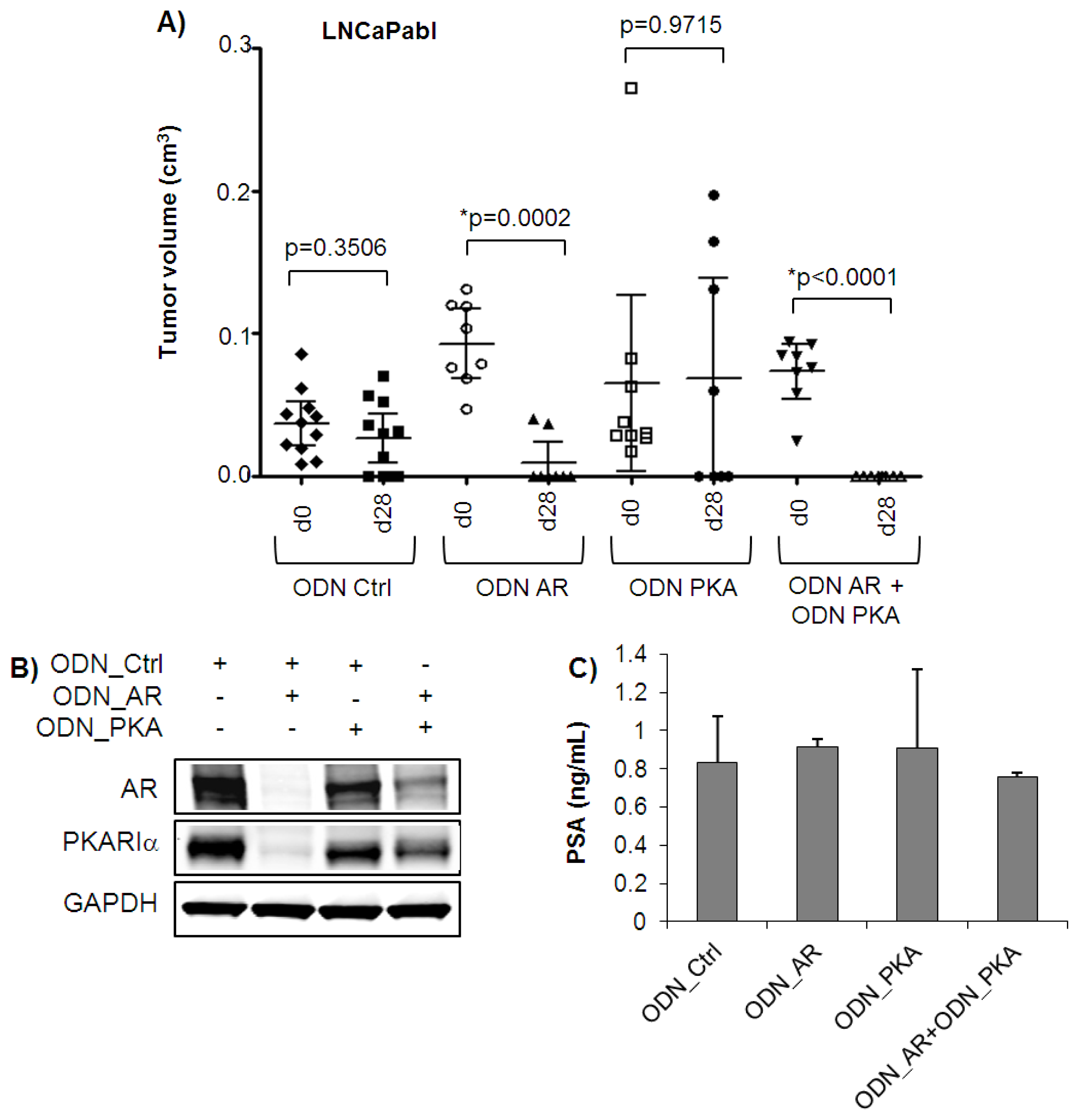
2.3. Complete Tumor Remission in Castration-Resistant LNCaPabl Tumors by Combined Targeting of AR and PKARIα
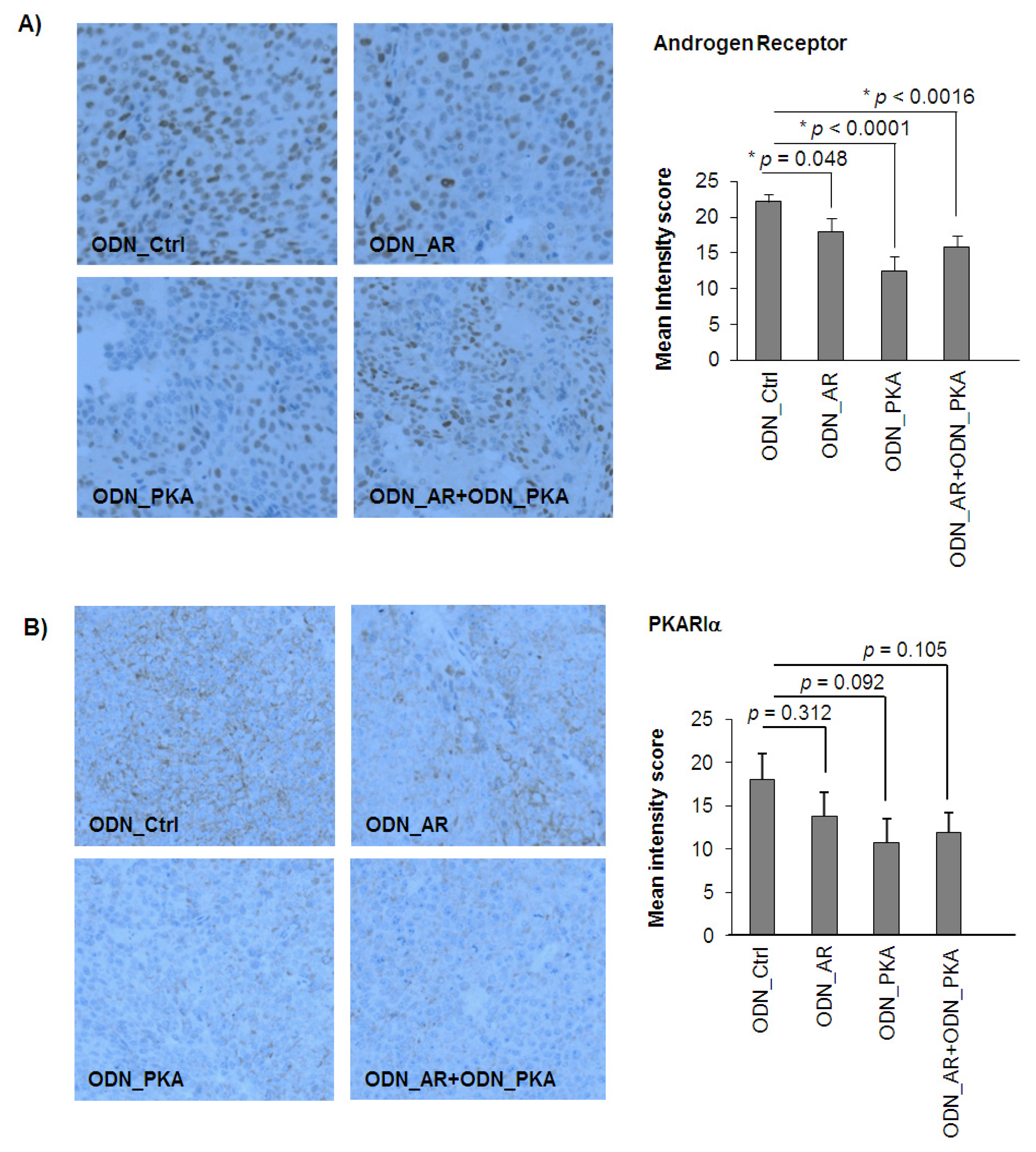
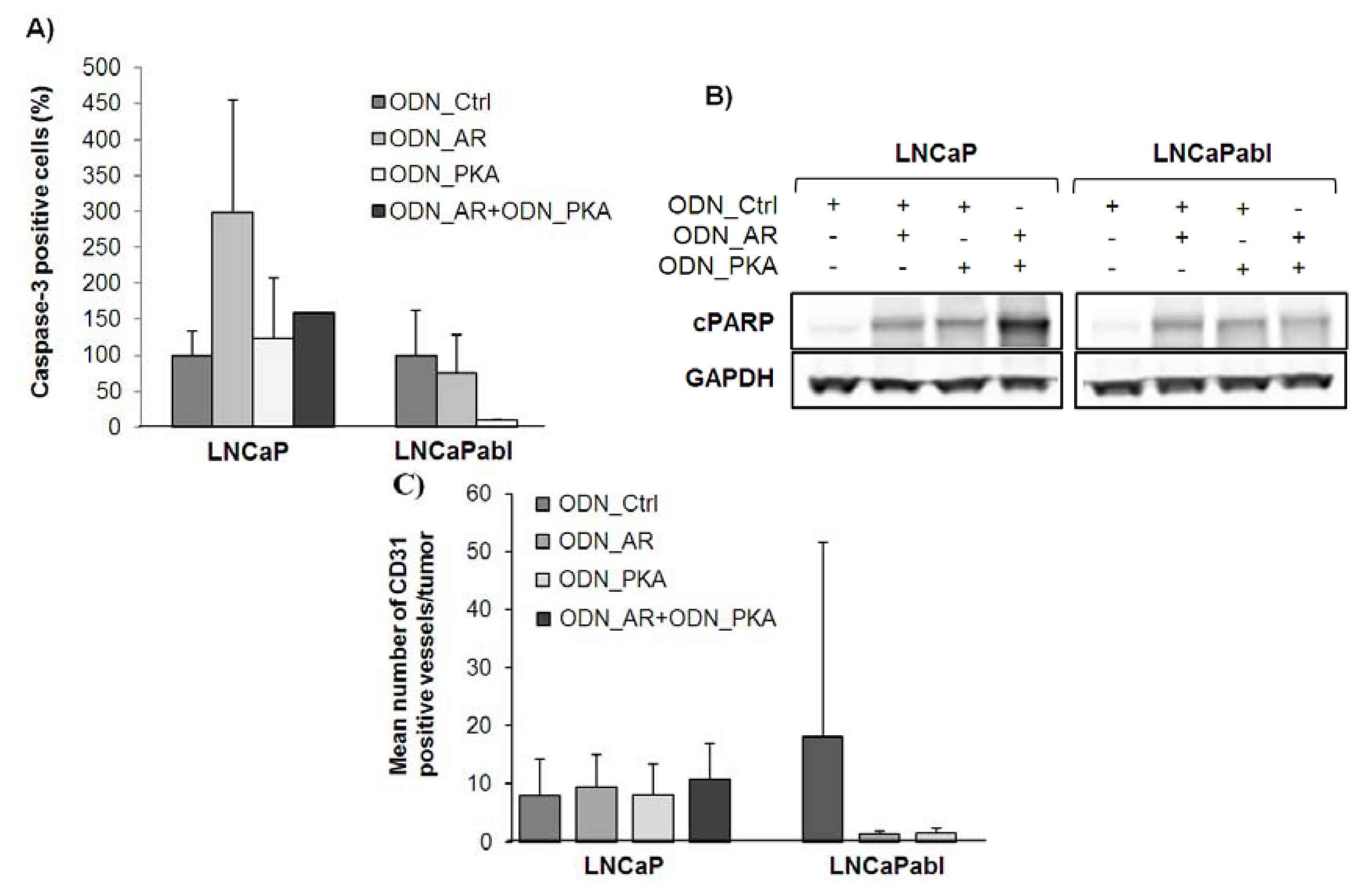
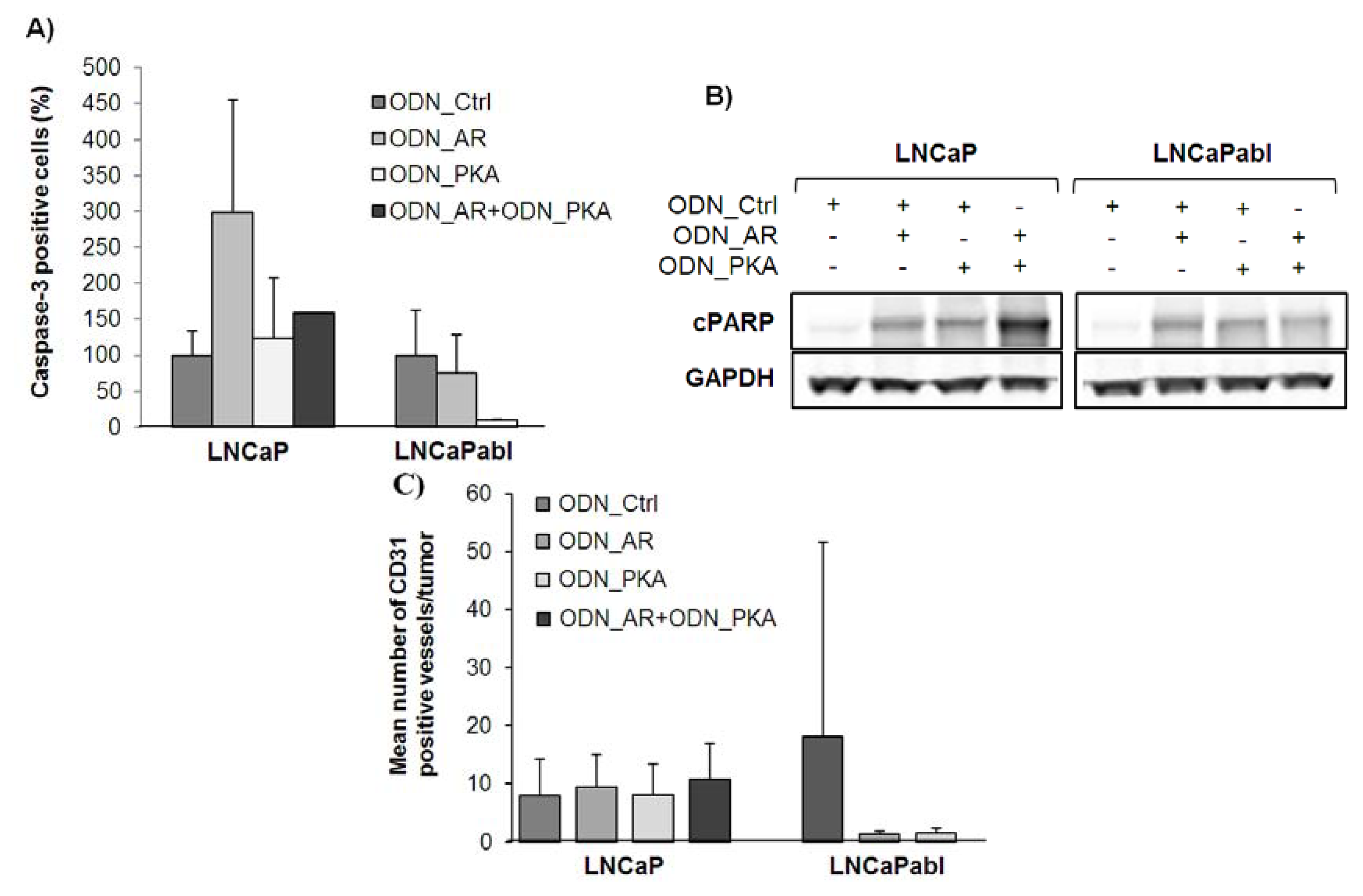
2.4. Effects of Single and Combined Targeting of AR and PKARIα on Apoptosis and Tumor Angiogenesis
2.5. Effects of Single and Combined Targeting of AR and PKARIα on Tumor Angiogenesis
3. Experimental Section
3.1. Drugs and Chemicals
3.2. Cell Lines
3.3. In Vitro Transfections with the ODNs
3.4. Flow Cytometric Detection of Apoptotic Cells
3.5. Immunoblotting
3.6. Establishment and Treatment of Human Prostate Tumor Xenografts in Nude Mice
3.7. PSA Measurement
3.8. Construction of LNCaP Tissue Microarray (TMA)
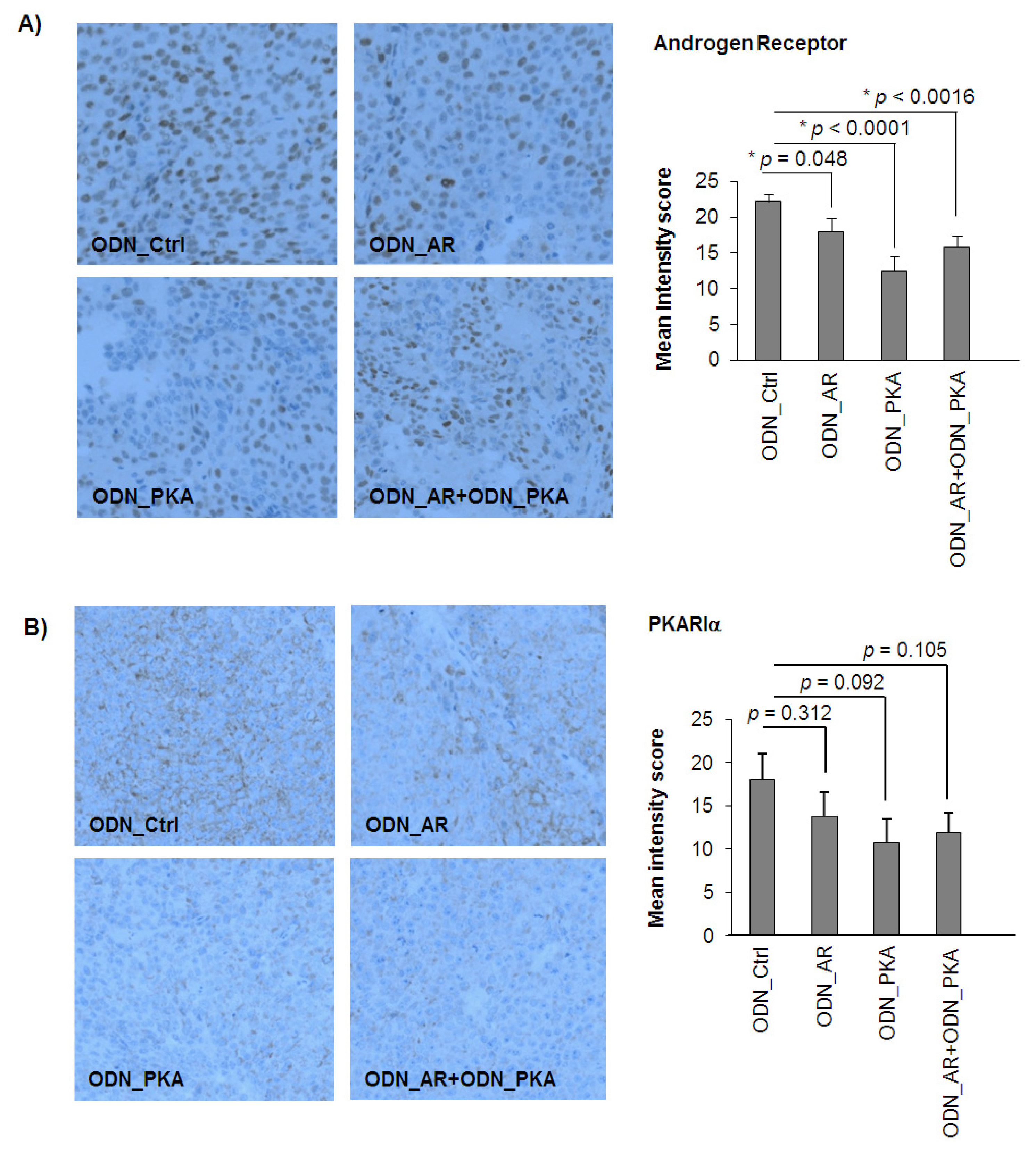
3.9. Immunohistochemistry
3.10. Statistics
4. Conclusions
Acknowledgements
Abbreviations
| AR | androgen receptor |
| CRPC | castration-resistant prostate cancer |
| PKARIα | regulatory subunit type I alpha of protein kinase A |
| PSA | prostate-specific antigen |
| ODN | oligodeoxynucleotide |
| TMA | tissue microarray |
| MVD | microvessel density |
| PARP-1 | poly(ADP-ribose)-polymerase 1 |
Conflict of Interest
References
- Wegiel, B.; Evans, S.; Hellsten, R.; Otterbein, L.E.; Bjartell, A.; Persson, J.L. Molecular pathways in the progression of hormone-independent and metastatic prostate cancer. Curr. Cancer Drug Targets 2010, 10, 392–401. [Google Scholar]
- Reis, L.O. Old issues and new perspectives on prostate cancer hormonal therapy: The molecular substratum. Med. Oncol 2011, 29, 1948–1955. [Google Scholar]
- Mohler, J.L.; Gregory, C.W.; Ford, O.H., III; Kim, D.; Weaver, C.M.; Petrusz, P.; Wilson, E.M.; French, F.S. The androgen axis in recurrent prostate cancer. Clin. Cancer Res 2004, 10, 440–448. [Google Scholar]
- Culig, Z.; Hobisch, A.; Cronauer, M.V.; Cato, A.C.; Hittmair, A.; Radmayr, C.; Eberle, J.; Bartsch, G.; Klocker, H. Mutant androgen receptor detected in an advanced-stage prostatic carcinoma is activated by adrenal androgens and progesterone. Mol. Endocrinol 1993, 7, 1541–1550. [Google Scholar]
- Buchanan, G.; Yang, M.; Harris, J.M.; Nahm, H.S.; Han, G.; Moore, N.; Bentel, J.M.; Matusik, R.J.; Horsfall, D.J.; Marshall, V.R.; et al. Mutations at the boundary of the hinge and ligand binding domain of the androgen receptor confer increased transactivation function. Mol. Endocrinol 2001, 15, 46–56. [Google Scholar]
- Veldscholte, J.; Ris-Stalpers, C.; Kuiper, G.G.J.M.; Jenster, G.; Berrevoets, C.; Claassen, E.; van Rooij, H.C.J.; Trapman, J.; Brinkmann, A.O.; Mulder, E. A mutation in the ligand binding domain of the androgen receptor of human LNCaP cells affects steroid binding characteristics and response to anti-androgens. Biochem. Biophys. Res. Comm 1990, 17, 534–540. [Google Scholar]
- Visakorpi, T.; Hyytinen, E.; Koivisto, P.; Tanner, M.; Keinänen, R.; Palmberg, C.; Palotie, A.; Tammela, T.; Isola, J.; Kallioniemi, O.-P. In vivo amplification of the androgen receptor gene and progression of human prostate cancer. Nat. Genet 1995, 9, 401–406. [Google Scholar]
- Gregory, C.W.; He, B.; Johnson, R.T.; Ford, O.H.; Mohler, J.L.; French, F.S.; Wilson, E.M. A mechanism for androgen receptor-mediated prostate cancer recurrence after androgen deprivation therapy. Cancer Res 2001, 61, 4315–4319. [Google Scholar]
- Culig, Z.; Hobisch, A.; Cronauer, M.V.; Radmayr, C.; Trapman, J.; Hittmair, A.; Bartsch, G.; Klocker, H. Androgen receptor activation in prostatic tumor cell lines by insulin-like growth factor-I, keratinocyte growth factor and epidermal growth factor. Cancer Res 1994, 54, 5474–5478. [Google Scholar]
- Nazareth, L.V.; Weigel, N.L. Activation of the human androgen receptor through a protein kinase A signaling pathway. J. Biol. Chem 1996, 271, 19900–19907. [Google Scholar]
- Craft, N.; Shostak, Y.; Carey, M.; Sawyers, C.L. A mechanism for hormone-independent prostate cancer through modulation of androgen receptor signaling by the HER-2/neu tyrosine kinase. Nat. Med 1999, 5, 280–285. [Google Scholar]
- Feldman, B.J.; Feldman, D. The development of androgen-independent prostate cancer. Nat. Rev. Cancer 2001, 1, 34–45. [Google Scholar]
- Koivisto, P.; Kolmer, M.; Visakorpi, T.; Kallioniemi, O.P. Androgen receptor gene and hormonal therapy failure of prostate cancer. Am. J. Pathol 1998, 152, 1–9. [Google Scholar]
- Coen, J.J.; Chung, C.S.; Shipley, W.U.; Zietman, A.L. Influence of follow-up bias on PSA failure after external beam radiotherapy for localized prostate cancer: Results from a 10-year cohort analysis. Int. J. Radiat. Oncol. Biol. Phys 2003, 57, 621–628. [Google Scholar]
- Eder, I.E.; Hoffmann, J.; Rogatsch, H.; Schäfer, G.; Zopf, D.; Bartsch, G.; Klocker, H. Inhibition of LNCaP prostate tumor growth in vivo by an antisense oligonucleotide directed against the human androgen receptor. Cancer Gene Therapy 2002, 9, 117–125. [Google Scholar]
- Chen, C.D.; Welsbie, D.S.; Tran, C.; Baek, S.H.; Chen, R.; Vessella, R.; Rosenfeld, M.G.; Sawyers, C.L. Molecular determinants of resistance to antiandrogen therapy. Nat. Med 2004, 10, 33–39. [Google Scholar]
- Cheng, H.; Snoek, R.; Ghaidi, F.; Cox, M.E.; Rennie, P.S. Short hairpin RNA knockdown of the androgen receptor attenuates ligand-independent activation and delays tumor progression. Cancer Res 2006, 66, 10613–10620. [Google Scholar]
- Haag, P.; Bektic, J.; Bartsch, G.; Klocker, H.; Eder, I.E. Androgen receptor down-regulation by small interference RNA induces cell growth inhibition in androgen sensitive as well as in androgen-independent prostate cancer cells. J. Steroid Biochem. Mol. Biol 2005, 96, 251–258. [Google Scholar]
- Liao, X.; Tang, S.; Thrasher, J.B.; Griebling, T.L.; Li, B. Small-interfering RNA-induced androgen receptor silencing leads to apoptotic cell death in prostate cancer. Mol. Cancer Ther 2005, 4, 505–515. [Google Scholar]
- Yang, Q.; Fung, K.M.; Day, W.V.; Kropp, B.P.; Lin, H.K. Androgen receptor signaling is required for androgen-sensitive human prostate cancer cell proliferation and survival. Cancer Cell. Int 2005, 5, 8. [Google Scholar]
- Desiniotis, A.; Schafer, G.; Klocker, H.; Eder, I.E. Enhanced antiproliferative and proapoptotic effects on prostate cancer cells by simultaneously inhibiting androgen receptor and cAMP-dependent protein kinase A. Int. J. Cancer 2010, 126, 775–789. [Google Scholar]
- Snoek, R.; Cheng, H.; Margiotti, K.; Wafa, L.A.; Wong, C.A.; Wong, E.C.; Fazli, L.; Nelson, C.C.; Gleave, M.E.; Rennie, P.S. In vivo knockdown of the androgen receptor results in growth inhibition and regression of well-established, castration-resistant prostate tumors. Clin. Cancer Res 2009, 15, 39–47. [Google Scholar]
- Mohler, M.L.; Coss, C.C.; Duke, C.B., III; Patil, S.A.; Miller, D.D.; Dalton, J.T. Androgen receptor antagonists: A patent review (2008–2011). Expert Opin. Ther. Pat 2012, 22, 541–565. [Google Scholar]
- Kim, W.; Ryan, C.J. Androgen receptor directed therapies in castration-resistant metastatic prostate cancer. Curr. Treat. Options Oncol 2012, 13, 189–200. [Google Scholar]
- Lee, S.O.; Ma, Z.; Yeh, C.R.; Luo, J.; Lin, T.H.; Lai, K.P.; Yamashita, S.; Liang, L.; Tian, J.; Li, L.; et al. New therapy targeting differential androgen receptor signaling in prostate cancer stem/progenitor vs non-stem/progenitor cells. J. Mol. Cell. Biol 2013, 5, 14–26. [Google Scholar]
- Cho-Chung, Y.S.; Nesterova, M.V. Tumor reversion: Protein kinase A isozyme switching. Ann. N. Y. Acad. Sci 2005, 1058, 76–86. [Google Scholar]
- Cho, Y.S.; Lee, Y.N.; Cho-Chung, Y.S. Biochemical characterization of extracellular cAMP-dependent protein kinase as a tumor marker. Biochem. Biophys. Res. Commun 2000, 278, 679–684. [Google Scholar]
- Cho, Y.S.; Cho-Chung, Y.S. Antisense protein kinase A RIalpha acts synergistically with hydroxycamptothecin to inhibit growth and induce apoptosis in human cancer cells: Molecular basis for combinatorial therapy. Clin. Cancer Res 2003, 9, 1171–1178. [Google Scholar]
- Merkle, D.; Hoffmann, R. Roles of cAMP and cAMP-dependent protein kinase in the progression of prostate cancer: Cross-talk with the androgen receptor. Cell Signal 2011, 23, 507–515. [Google Scholar]
- Bagchi, G.; Wu, J.; French, J.; Kim, J.; Moniri, N.H.; Daaka, Y. Androgens transduce the G alphas-mediated activation of protein kinase A in prostate cells. Cancer Res 2008, 68, 3225–3231. [Google Scholar]
- Sadar, M.D. Androgen-independent induction of prostate-specific antigen gene expression via cross-talk between the androgen receptor and protein kinase A signal transduction pathways. J. Biol. Chem 1999, 274, 7777–7783. [Google Scholar]
- Sadar, M.D.; Hussain, M.; Bruchovsky, N. Prostate cancer: Molecular biology of early progression to androgen independence. Endocr. Relat. Cancer 1999, 6, 487–502. [Google Scholar]
- Chen, S.H.; Zhaori, G. Potential clinical applications of siRNA technique: Benefits and limitations. Eur. J. Clin. Invest 2011, 41, 221–232. [Google Scholar]
- LaCasse, E.C.; Cherton-Horvat, G.G.; Hewitt, K.E.; Jerome, L.J.; Morris, S.J.; Kandimalla, E.R.; Yu, D.; Wang, H.; Wang, W.; Zhang, R.; et al. Preclinical characterization of AEG35156/GEM 640, a second-generation antisense oligonucleotide targeting X-linked inhibitor of apoptosis. Clin. Cancer Res 2006, 12, 5231–5241. [Google Scholar]
- Pfeil, K.; Eder, I.E.; Putz, T.; Ramoner, R.; Culig, Z.; Ueberall, F.; Bartsch, G.; Klocker, H. Long-term androgen-ablation causes increased resistance to PI3K/Akt pathway inhibition in prostate cancer cells. Prostate 2004, 58, 259–268. [Google Scholar]
- Chan, S.C.; Li, Y.; Dehm, S.M. Androgen receptor splice variants activate androgen receptor target genes and support aberrant prostate cancer cell growth independent of canonical androgen receptor nuclear localization signal. J. Biol. Chem 2012, 287, 19736–19749. [Google Scholar]
- Kozlowski, P.; Wong, J.; Goldenberg, S.L. Serial tumour blood-flow measurements in androgen-dependent and -independent Shionogi tumour models. BJU Int 2005, 95, 644–649. [Google Scholar]
- Weidner, N.; Carroll, P.R.; Flax, J.; Blumenfeld, W.; Folkman, J. Tumor angiogenesis correlates with metastasis in invasive prostate carcinoma. Am. J. Pathol 1993, 143, 401–409. [Google Scholar]
- Offersen, B.V.; Borre, M.; Overgaard, J. Quantification of angiogenesis as a prognostic marker in human carcinomas: A critical evaluation of histopathological methods for estimation of vascular density. Eur. J. Cancer 2003, 39, 881–890. [Google Scholar]
- Bono, A.V.; Celato, N.; Cova, V.; Salvadore, M.; Chinetti, S.; Novario, R. Microvessel density in prostate carcinoma. Prostate Cancer Prostatic Dis 2002, 5, 123–127. [Google Scholar]
- Pallares, J.; Rojo, F.; Iriarte, J.; Morote, J.; Armadans, L.I.; de Torres, I. Study of microvessel density and the expression of the angiogenic factors VEGF, bFGF and the receptors Flt-1 and FLK-1 in benign, premalignant and malignant prostate tissues. Histol. Histopathol 2006, 21, 857–865. [Google Scholar]
- Bates, M.; Kovalenko, B.; Wilson, E.L.; Moscatelli, D. Endothelial cells support the growth of prostate tissue in vivo. Prostate 2008, 68, 893–901. [Google Scholar]
- Godoy, A.; Watts, A.; Sotomayor, P.; Montecinos, V.P.; Huss, W.J.; Onate, S.A.; Smith, G.J. Androgen receptor is causally involved in the homeostasis of the human prostate endothelial cell. Endocrinology 2008, 149, 2959–2969. [Google Scholar]
- Compagno, D.; Merle, C.; Morin, A.; Gilbert, C.; Mathieu, J.R.; Bozec, A.; Mauduit, C.; Benahmed, M.; Cabon, F. SIRNA-directed in vivo silencing of androgen receptor inhibits the growth of castration-resistant prostate carcinomas. PLoS One 2007, 2, e1006. [Google Scholar]
- Cheng, L.; Zhang, S.; Sweeney, C.J.; Kao, C.; Gardner, T.A.; Eble, J.N. Androgen withdrawal inhibits tumor growth and is associated with decrease in angiogenesis and VEGF expression in androgen-independent CWR22Rv1 human prostate cancer model. Anticancer Res 2004, 24, 2135–2140. [Google Scholar]
- Gustavsson, H.; Welen, K.; Damber, J.E. Transition of an androgen-dependent human prostate cancer cell line into an androgen-independent subline is associated with increased angiogenesis. Prostate 2005, 62, 364–373. [Google Scholar]
- Culig, Z.; Hoffmann, J.; Erdel, M.; Eder, I.E.; Hobisch, A.; Hittmair, A.; Bartsch, G.; Utermann, G.; Schneider, M.R.; Parczyk, K.; et al. Switch from antagonist to agonist of the androgen receptor blocker bicalutamide is associated with prostate tumour progression in a new model system. Br. J. Cancer 1999, 81, 242–251. [Google Scholar]
- Hobisch, A.; Hoffmann, J.; Lambrinidis, L.; Eder, I.E.; Bartsch, G.; Klocker, H.; Culig, Z. Antagonist/agonist balance of the nonsteroidal antiandrogen bicalutamide (Casodex) in a new prostate cancer model. Urol. Int 2000, 65, 73–79. [Google Scholar]
- Hensley, H.H.; Hannoun-Levi, J.M.; Hachem, P.; Mu, Z.; Stoyanova, R.; Khor, L.Y.; Agrawal, S.; Pollack, A. PKA knockdown enhances cell killing in response to radiation and androgen deprivation. Int. J. Cancer 2011, 128, 962–973. [Google Scholar]

© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Eder, I.E.; Egger, M.; Neuwirt, H.; Seifarth, C.; Maddalo, D.; Desiniotis, A.; Schäfer, G.; Puhr, M.; Bektic, J.; Cato, A.C.B.; et al. Enhanced Inhibition of Prostate Tumor Growth by Dual Targeting the Androgen Receptor and the Regulatory Subunit Type Iα of Protein Kinase A in Vivo. Int. J. Mol. Sci. 2013, 14, 11942-11962. https://doi.org/10.3390/ijms140611942
Eder IE, Egger M, Neuwirt H, Seifarth C, Maddalo D, Desiniotis A, Schäfer G, Puhr M, Bektic J, Cato ACB, et al. Enhanced Inhibition of Prostate Tumor Growth by Dual Targeting the Androgen Receptor and the Regulatory Subunit Type Iα of Protein Kinase A in Vivo. International Journal of Molecular Sciences. 2013; 14(6):11942-11962. https://doi.org/10.3390/ijms140611942
Chicago/Turabian StyleEder, Iris E., Martina Egger, Hannes Neuwirt, Christof Seifarth, Danilo Maddalo, Andreas Desiniotis, Georg Schäfer, Martin Puhr, Jasmin Bektic, Andrew C. B. Cato, and et al. 2013. "Enhanced Inhibition of Prostate Tumor Growth by Dual Targeting the Androgen Receptor and the Regulatory Subunit Type Iα of Protein Kinase A in Vivo" International Journal of Molecular Sciences 14, no. 6: 11942-11962. https://doi.org/10.3390/ijms140611942