Antioxidant Properties and PC12 Cell Protective Effects of a Novel Curcumin Analogue (2E,6E)-2,6-Bis(3,5- dimethoxybenzylidene)cyclohexanone (MCH)

Abstract
:1. Introduction
2. Results
2.1. Free Radical Scavenging Activities and Reducing Power of MCH
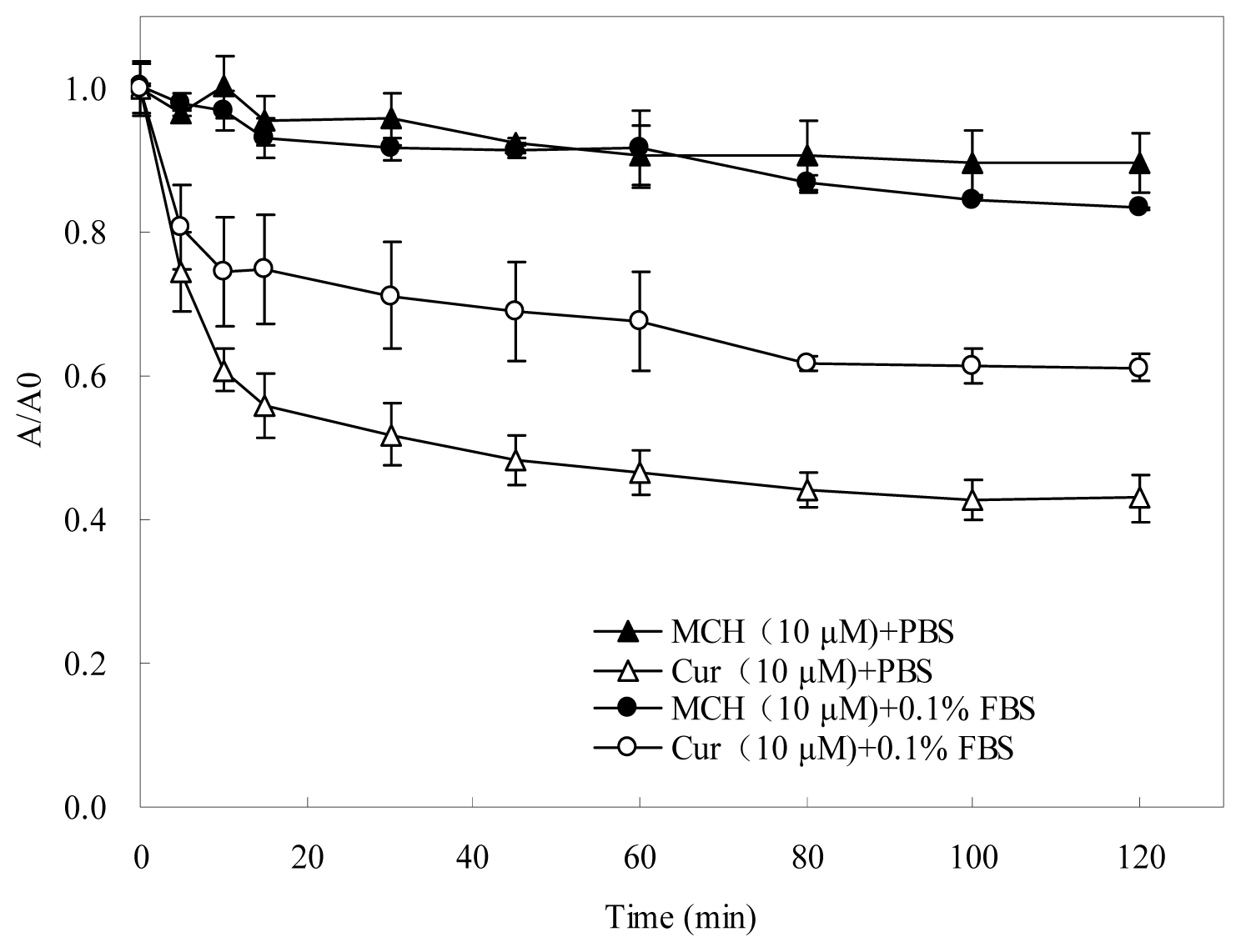
2.2. Stability of MCH and Curcumin
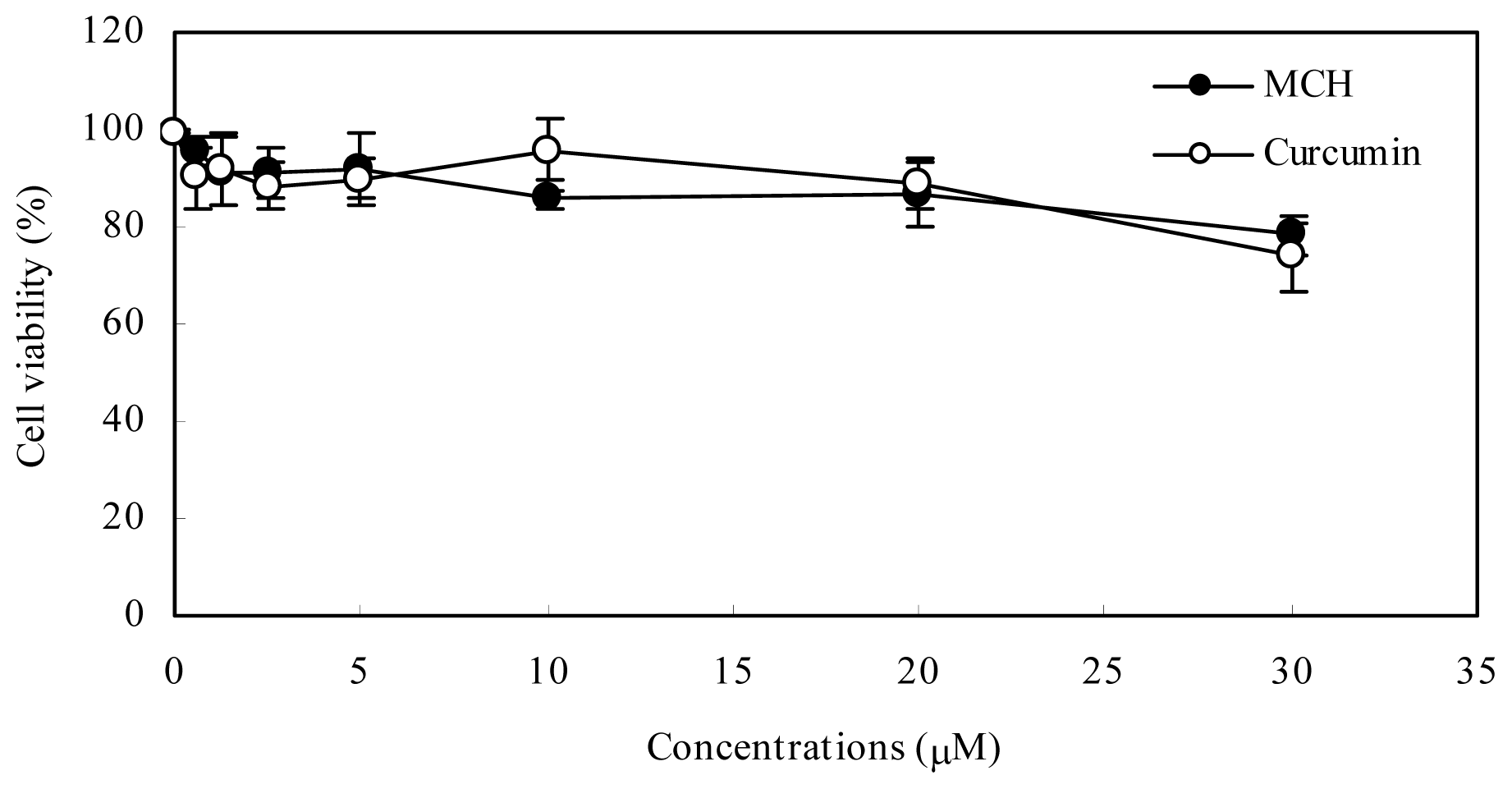
2.3. Cytotoxicity of MCH in PC12 Cells
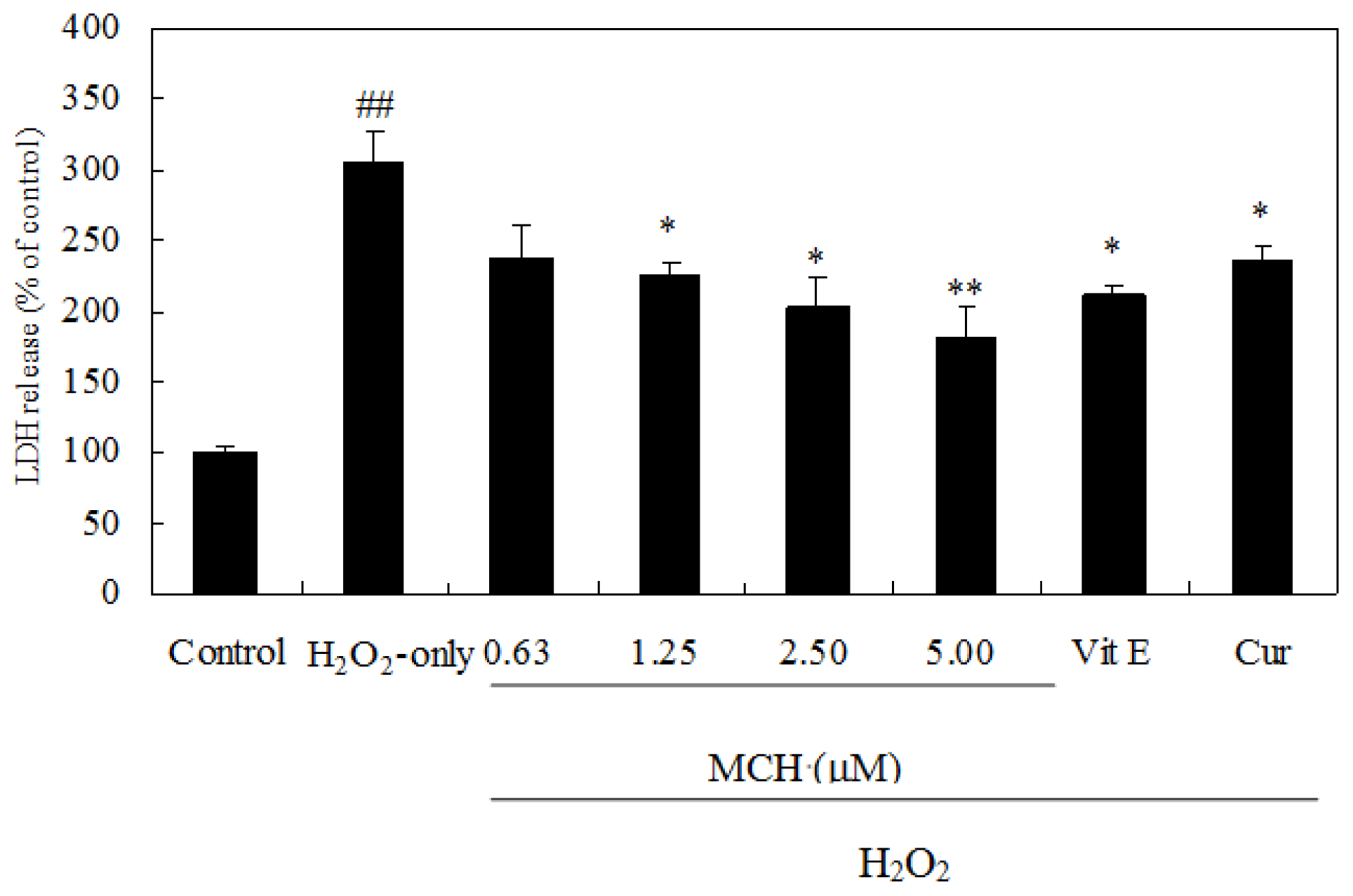
2.4. MCH Protects PC12 Cells against H2O2-Induced Cytotoxicity
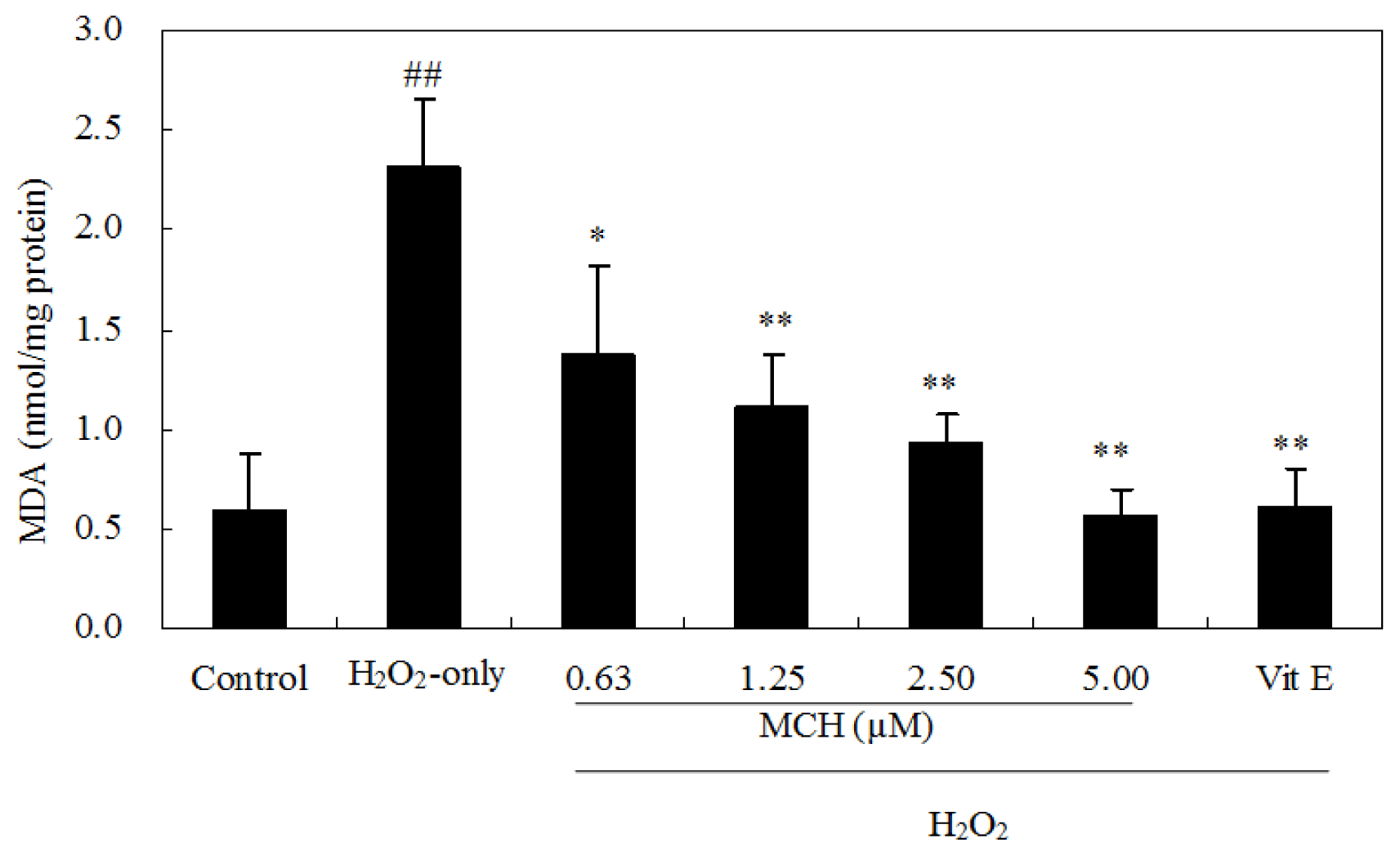
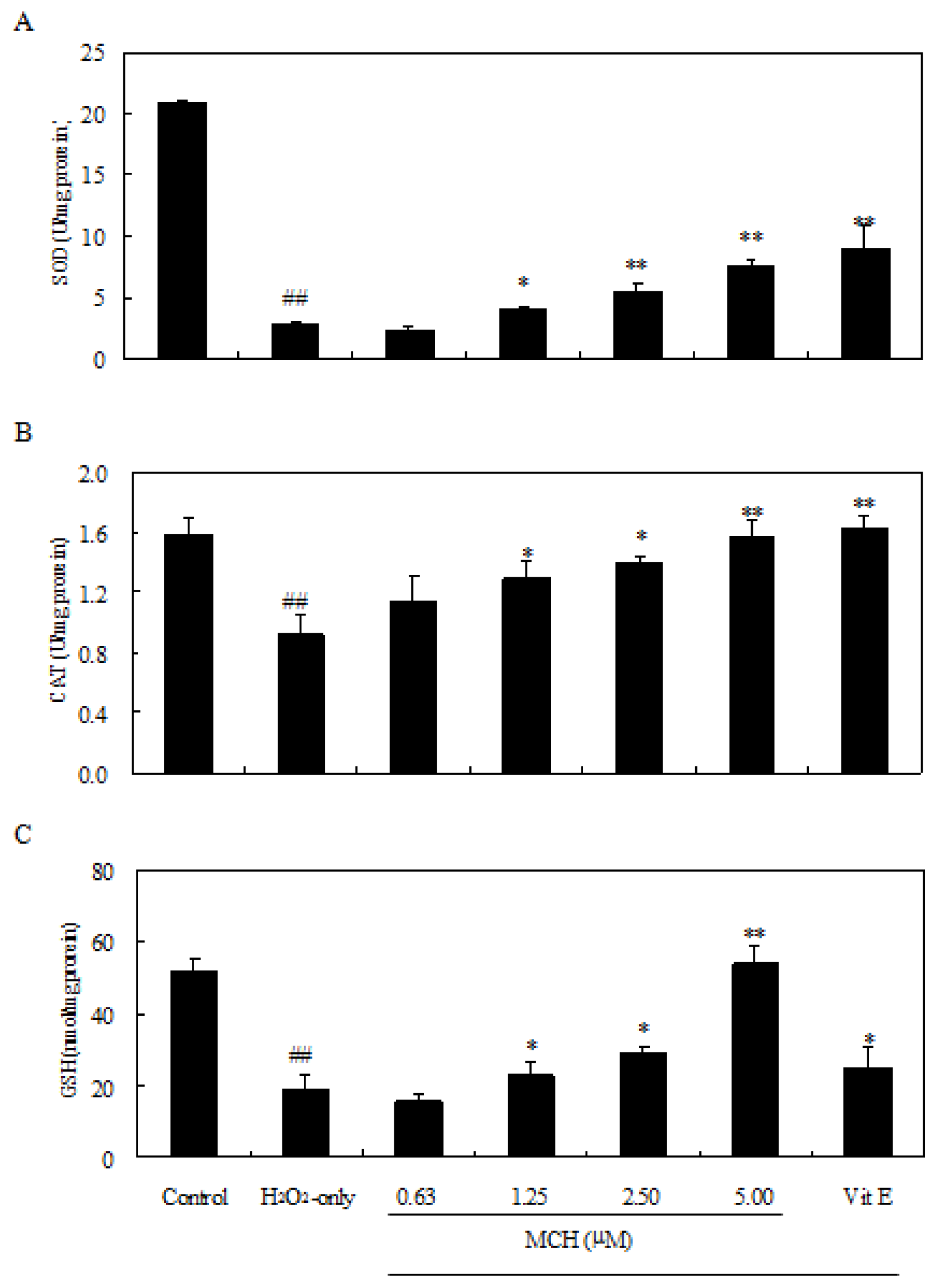
2.5. MCH Reduced Lipid Peroxidation and Rescued Loss of Antioxidant Enzyme Activities in H2O2-Treated PC12 Cells
2.6. MCH Prevents H2O2-Induced ROS Generation
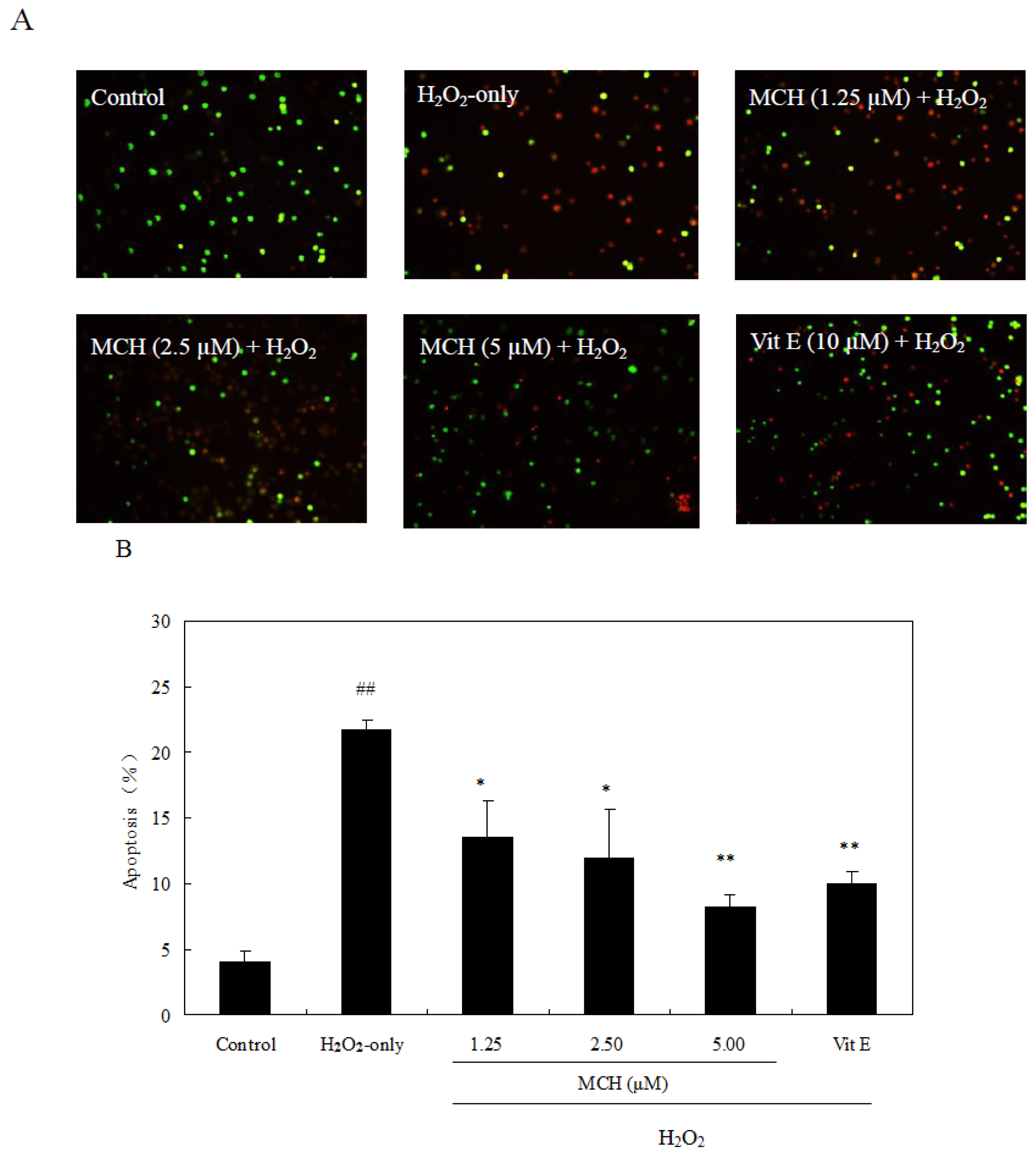
2.7. MCH Protected PC12 Cells against H2O2-Induced Apoptosis
2.8. MCH Prevented Loss of MMP in PC12 Cells
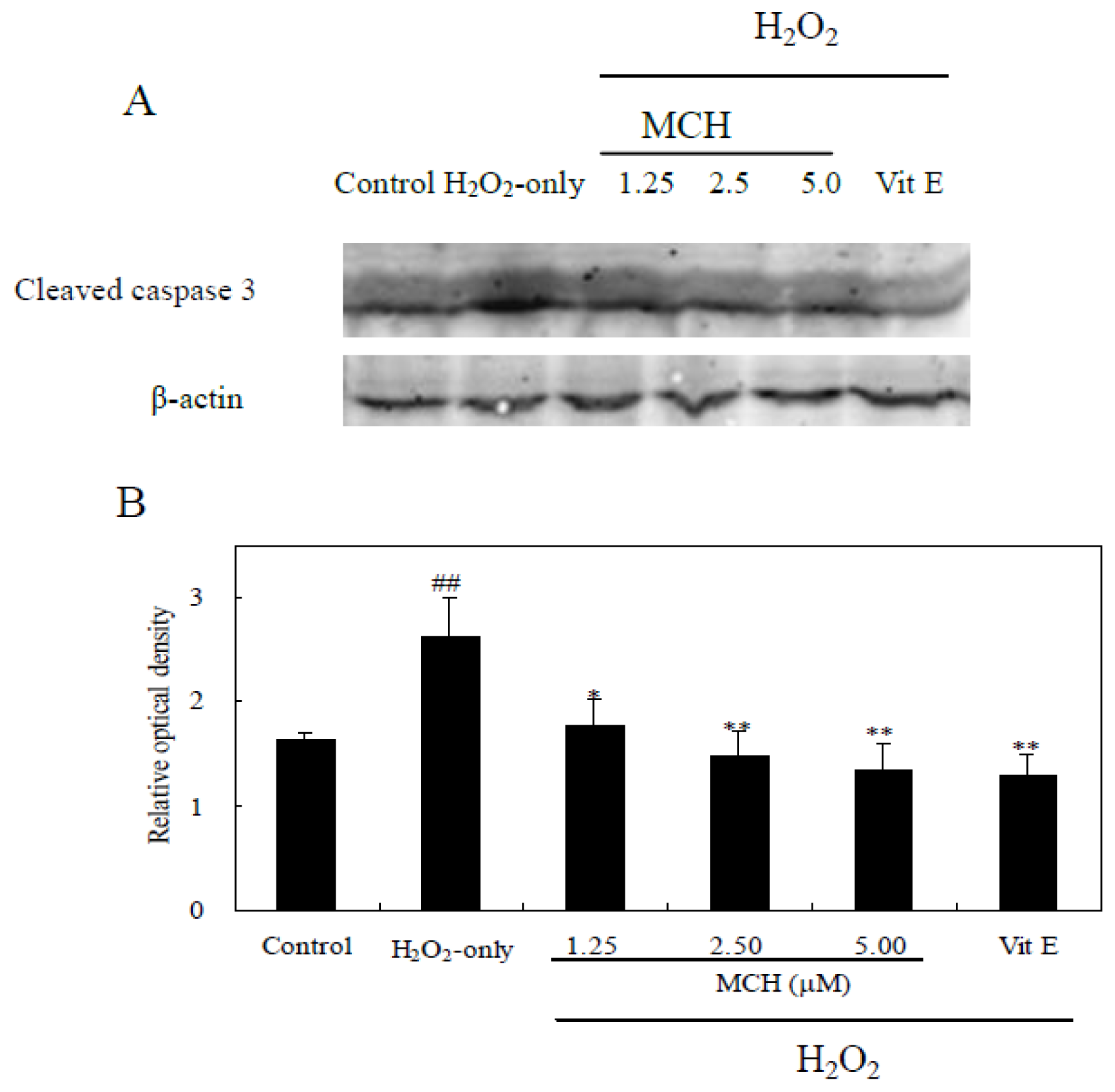
2.9. MCH Inhibited H2O2-Induced Expression of Cleaved Caspase-3 in PC12 Cells
2.10. MCH Promoted the Expression of Nrf2 in PC12 Cells
3. Discussion
4. Materials and Methods
4.1. Materials
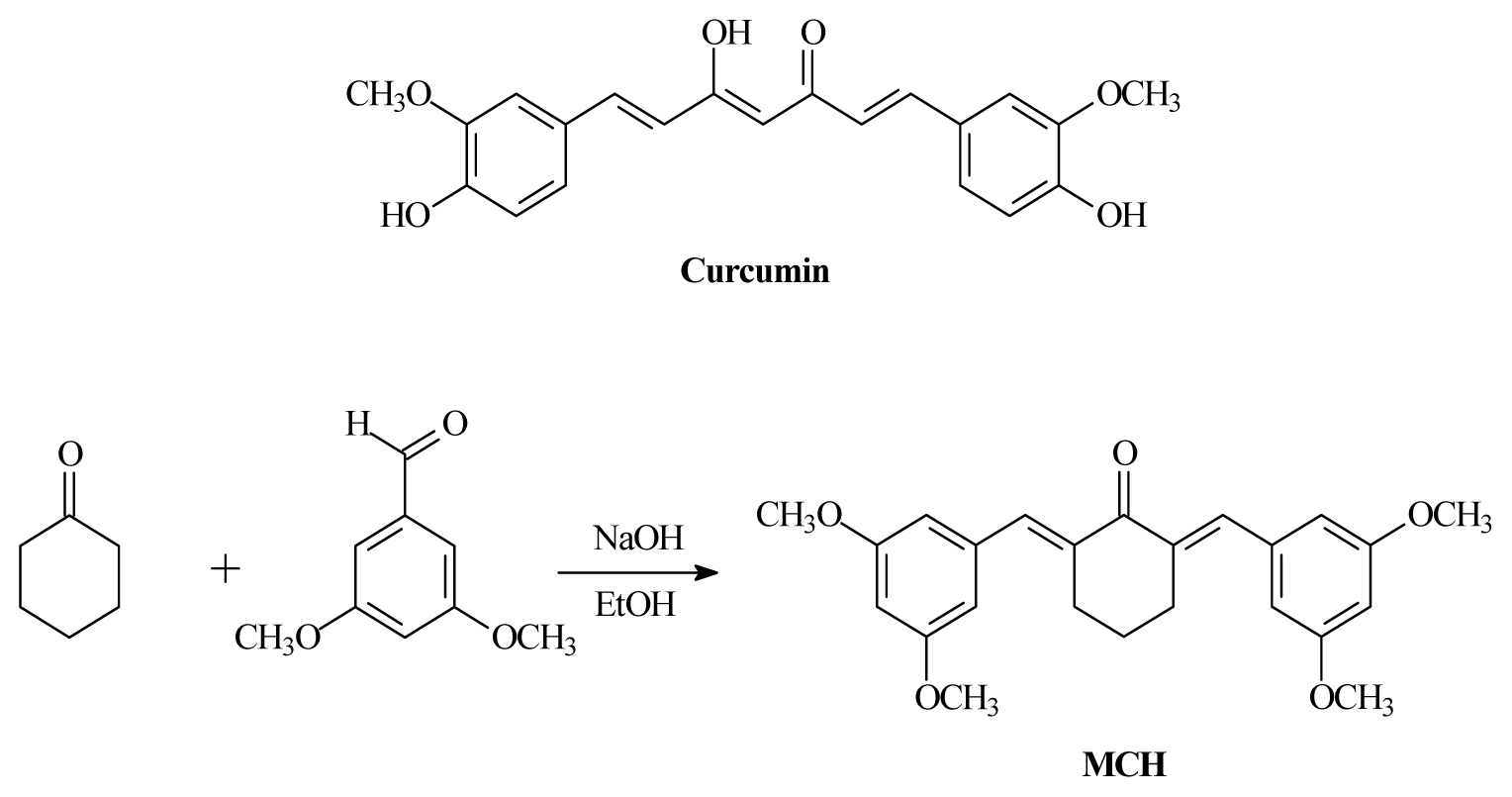
4.2. Synthesis of MCH
4.3. DPPH Radical Scavenging Assay
4.4. Superoxide Radical Scavenging Assay
4.5. Hydroxyl Radical Scavenging Assay
4.6. Stability Studies in Vitro
4.7. Culture of PC12 Cells
4.8. Measurement of Cell Viability
4.9. Lactate Dehydrogenase (LDH) Assay
4.10. Measurement of Intracellular ROS Accumulation
4.11. Assays for GSH Content and Antioxidant Enzymes
4.12. Measurement of Mitochondrial Membrane Potential (MMP)
4.13. Apoptosis Analysis by AO/EB and Annexin V-FITC/PI Staining
4.14. Analysis of Cleaved Caspase-3 Expression with Western Blot
4.15. Semi-Quantitative RT-PCR Analysis
4.16. Statistical Analysis
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Goel, A.; Kunnumakkara, A.B.; Aggarwal, B.B. Curcumin as “Curecumin”: From kitchen to clinic. Biochem. Pharmacol. 2008, 75, 787–809. [Google Scholar]
- Strimpakos, A.S.; Sharma, R.A. Curcumin: Preventive and therapeutic properties in laboratory studies and clinical trials. Antioxid. Redox Signal. 2008, 10, 511–549. [Google Scholar]
- Hamaguchi, T.; Ono, K.; Yamada, M. Curcumin and Alzheimer’s disease. CNS Neurosci. Ther. 2010, 16, 285–297. [Google Scholar]
- Anand, P.; Kunnumakkara, A.B.; Newman, R.A.; Aggarwal, B.B. Bioavailability of curcumin: Problems and promises. Mol. Pharm. 2007, 4, 807–818. [Google Scholar]
- Youssef, K.M.; El-Sherbeny, M.A.; El-Shafie, F.S.; Farag, H.A.; Al-Deeb, O.A.; Awadalla, S.A. Synthesis of curcumin analogues as potential antioxidant cancer chemopreventive agents. Arch. Pharm. 2004, 337, 42–54. [Google Scholar]
- Amolins, M.W.; Peterson, L.B.; Blagg, B.S.J. Synthesis and evaluation of electron-rich curcumin analogues. Bioorg. Med. Chem. 2009, 17, 360–367. [Google Scholar]
- Lee, K.H.; Chow, Y.L.; Sharmili, V.; Abas, F.; Alitheen, N.B.; Shaari, K.; Israf, D.A.; Lajis, N.H.; Syahida, A. BDMC33, A curcumin derivative suppresses inflammatory responses in macrophage-like cellular system: Role of inhibition in NF-κB and MAPK signaling pathways. Int. J. Mol. Sci. 2012, 13, 2985–3008. [Google Scholar]
- Lee, K.H.; Abas, F.; Alitheen, N.B.; Shaari, K.; Lajis, N.H.; Ahmad, S. A curcumin derivative 26-bis(25-dimethoxybenzylidene)-cyclohexanone (BDMC33) attenuates prostaglandin E2 synthesis via selective suppression of cyclooxygenase-2 in IFN-γ/LPS-stimulated macrophages. Molecules 2011, 1, 9728–9738. [Google Scholar]
- Bhullar, K.S.; Jha, A.; Youssef, D.; Rupasinghe, H.P. Curcumin and its carbocyclic analogs: Structure-activity in relation to antioxidant and selected biological properties. Molecules 2013, 10, 5389–5404. [Google Scholar]
- Wang, Y.J.; Pan, M.H.; Cheng, A.L.; Lin, L.I.; Ho, Y.S.; Hsieh, C.Y.; Lin, J.K. Stability of curcumin in buffer solutions and characterization of its degradation products. J. Pharm. Biomed. Anal. 1997, 15, 1867–1876. [Google Scholar]
- Liang, G.; Li, X.; Chen, L.; Yang, S.L.; Wu, X.D.; Studer, E.; Gurley, E.; Hylemon, P.B.; Ye, F.Q.; Li, Y.R.; et al. Synthesis and anti-inflammatory activities of mono-carbonyl analogues of curcumin. Bioorg. Med. Chem. Lett. 2008, 18, 1525–1529. [Google Scholar]
- Sardjiman, S.S.; Reksohadiprodjo, M.S.; Hakim, L.; van der Goot, H.; Timmerman, H. l5-Diphenyl-14-pentadiene-3-ones and cyclic analogues as antioxidative agents Synthesis and structure-activity relationship. Eur. J. Med. Chem. 1997, 32, 625–630. [Google Scholar]
- Ringman, J.M.; Frautschy, S.A.; Cole, G.M.; Masterman, D.L.; Cummings, J.L. A potential role of the curry spice curcumin in Alzheimer’s disease. Curr. Alzheimer Res. 2005, 2, 131–136. [Google Scholar]
- Kim, D.S.; Park, S.Y.; Kim, J.K. Curcuminoids from Curcuma longa L (Zingiberaceae) that protect PC12 rat pheochromocytoma and normal human umbilical vein endothelial cells from βA(1–42) insult. Neurosci. Lett. 2001, 303, 57–61. [Google Scholar]
- Xu, Y.-Y.; Cao, Y.; Ma, H.K.; Li, H.-Q.; Ao, G.-Z. Design synthesis and molecular docking of αβ-unsaturated cyclohexanone analogous of curcumin as potent EGFR inhibitors with antiproliferative activity. Bioorg. Med. Chem. 2013, 21, 388–394. [Google Scholar]
- Chipuk, J.E.; Bouchier-Hayes, L.; Green, D.R. Mitochondrial outer membrane permeabilization during apoptosis: The innocent bystander scenario. Cell Death Differ. 2006, 13, 1396–1402. [Google Scholar]
- Wang, X. The expanding role of mitochondria in apoptosis. Genes Dev. 2001, 15, 2922–2933. [Google Scholar]
- Yang, C.; Zhang, X.; Fan, H.; Liu, Y. Curcumin upregulates transcription factor Nrf2 HO-1 expression and protects rat brains against focal ischemia. Brain Res. 2009, 1282, 133–141. [Google Scholar]
- Barclay, L.R.C.; Vinqvist, M.R.; Mukai, K.; Goto, H.; Hashimoto, Y.; Tokunaga, A.; Uno, H. On the antioxidant mechanism of curcumin: Classical methods are needed to determine antioxidant mechanism and activity. Org. Lett. 2000, 2, 2841–2843. [Google Scholar]
- Chance, B.; Sies, H.; Boveris, A. Hydroperoxide metabolism in mammalian organs. Physiol. Rev. 1979, 59, 527–605. [Google Scholar]
- Tusi, S.K.; Ansari, N.; Amini, M.; Amirabad, A.D.; Shafiee, A.; Khodagholi, F. Attenuation of NF-κB and activation of Nrf2 signaling by 124-triazine derivatives protects neuron-like PC12 cells against apoptosis. Apoptosis 2010, 15, 738–751. [Google Scholar]
- Armstrong, J.S.; Steinauer, K.K.; Hornung, B.; Irish, J.M.; Lecane, P.; Birrell, G.W.; Peehl, D.M.; Knox, S.J. Role of glutathione depletion and reactive oxygen species generation in apoptotic signaling in a human B lymphoma cell line. Cell Death Differ. 2002, 9, 252–263. [Google Scholar]
- Yang, C.F.; Shen, H.M.; Ong, C.N. Intracellular thiol depletion causes mitochondrial permeability transition in ebselen-induced apoptosis. Arch. Biochem. Biophys. 2000, 380, 319–330. [Google Scholar]
- Ishii, T.; Itoh, K.; Takahashi, S.; Sato, H.; Yanagawa, T.; Katoh, Y.; Bannai, S.; Yamamoto, M. Transcription factor Nrf2 coordinately regulates a group of oxidative stress-inducible genes in macrophages. J. Biol. Chem. 2000, 275, 16023–16029. [Google Scholar]
- Pugazhenthi, S.; Akhov, L.; Selvaraj, G.; Wang, M.; Alam, J. Regulation of heme oxygenase-1 expression by demethoxy curcuminoids through Nrf2 by a PI3-kinase/Akt-mediated pathway in mouse beta-cells. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E645–E655. [Google Scholar]
- Aziz, M.T.A.; Ibrashy, I.N.E.; Mikhailidis, D.P.; Rezq, A.M.; Wassef, M.A.A.; Fouad, H.H.; Ahmed, H.H.; Sabry, D.A.; Shawky, H.M.; Hussein, R.E. Signaling mechanisms of a water soluble curcumin derivative in experimental type 1 diabetes with cardiomyopathy. Diabetol. Metab. Syndr. 2013, 5, 1–13. [Google Scholar]
- Jeong, G.S.; Oh, G.S.; Pae, H.O.; Jeong, S.O.; Kim, Y.C.; Shin, M.K.; Seo, B.Y.; Han, S.Y.; Lee, H.S.; Jeong, J.G.; et al. Comparative effects of curcuminoids on endothelial heme oxygenase-1 expression: Ortho-methoxy groups are essential to enhance heme oxygenase activity and protection. Exp. Mol. Med. 2006, 38, 393–400. [Google Scholar]
- Dinkova-Kostova, A.T.; Massiah, M.A.; Bozak, R.E.; Hicks, R.J.; Talalay, P. Potency of Michael reaction acceptors as inducers of enzymes that protect against carcinogenesis depends on their reactivity with sulfhydryl groups. Proc. Natl. Acad. Sci. USA 2001, 98, 3404–3409. [Google Scholar]
- Simon, H.U.; Haj-Yehia, A.; Levi-Schaffer, F. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis 2000, 5, 415–418. [Google Scholar]
- Stridh, H.; Kimland, M.; Jones, D.P.; Orrenius, S.; Hampton, M.B. Cytochrome c release and caspase activation in hydrogen peroxide- and tributyltin-induced apoptosis. FEBS Lett. 1998, 429, 351–355. [Google Scholar]
- Gao, L.W.; Wang, J.W. Antioxidant potential and DNA damage protecting activity of aqueous extract from Armillaria mellea. J. Food Biochem. 2012, 36, 139–148. [Google Scholar]
- Sun, C.; Wang, J.W.; Fang, L.; Gao, X.D.; Tan, R.X. Free radical scavenging and antioxidant activities of EPS2 an exopolysaccharide produced by a marine filamentous fungus Keissleriella sp YS 4108. Life Sci. 2004, 75, 1063–1073. [Google Scholar]
- Fang, X.B.; Fang, L.; Gou, S.H.; Cheng, L. Design and synthesis of dimethylaminomethyl-substituted curcumin derivatives/analogues: Potent antitumor and antioxidant activity improved stability and aqueous solubility compared with curcumin. Bioorg. Med. Chem. Lett. 2013, 23, 1297–1301. [Google Scholar]
- Takahashi, M.; Shibata, M.; Niki, E. Estimation of lipid peroxidation of live cells using a fluorescent probe diphenyl-1-pyrenylphosphine. Free Radic. Biol. Med. 2001, 31, 164–174. [Google Scholar]
- Cohen, G.; Dembiec, D.; Marcus, J. Measurement of catalase in tissue extracts. Anal. Biochem. 1970, 34, 30–38. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar]
- Chen, W.; Zhao, Z.; Li, L.; Wu, B.; Chen, S.-F.; Zhou, H.; Wang, Y.; Li, Y.-Q. Hispolon induces apoptosis in human gastric cancer cells through a ROS-mediated mitochondrial pathway. Free Radic. Biol. Med. 2008, 45, 60–72. [Google Scholar]
- Walker, J.M. The Bicinchoninic Acid (BCA) Assay for Protein Quantitation. In The Protein Protocols Handbook; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 1994; Volume 32, pp. 5–8. [Google Scholar]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar]
- Li, H.Y.; Wu, S.Y.; Shi, N.A. Transcription factor Nrf2 activation by deltamethrin in PC12 cells: Involvement of ROS. Toxicol. Lett. 2007, 171, 87–98. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | EC50 (μM) (b) | ||
|---|---|---|---|
| DPPH | Superoxide anion | Hydroxyl radical | |
| MCH | 466.79 ± 30.50 ** | 64.78 ± 7.72 ** | 94.03 ± 1.12 ## |
| Curcumin | 14.24 ± 0.60 | 88.09 ± 5.97 ** | 58.84 ± 3.14 |
| Vitamin C | 13.00 ± 0.37 | 100.93 ± 4.10 | n.d. (c) |
| Treatment | ROS (percentage of the control %) | MMP (percentage of the control %) |
|---|---|---|
| Control (0.2% DMSO) | 100 ± 7.35 | 100 ± 7.58 |
| H2O2-only (150 μM) | 426.09 ± 36.20 ## | 48.32 ± 1.14 ## |
| MCH (0.63 μM) + H2O2 | 294.78 ± 57.32 ** | 74.84 ± 5.05 ** |
| MCH (1.25 μM) + H2O2 | 269.18 ± 1.17 ** | 74.01 ± 4.72 ** |
| MCH (2.50 μM) + H2O2 | 248.70 ± 22.55 ** | 79.78 ± 1.55 ** |
| MCH (5.00 μM) + H2O2 | 231.59 ± 12.90 ** | 82.01 ± 5.53 ** |
| Vit E (10.00 μM) + H2O2 | 241.16 ± 4.06 ** | 86.64 ± 3.95 ** |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ao, G.-Z.; Chu, X.-J.; Ji, Y.-Y.; Wang, J.-W. Antioxidant Properties and PC12 Cell Protective Effects of a Novel Curcumin Analogue (2E,6E)-2,6-Bis(3,5- dimethoxybenzylidene)cyclohexanone (MCH). Int. J. Mol. Sci. 2014, 15, 3970-3988. https://doi.org/10.3390/ijms15033970
Ao G-Z, Chu X-J, Ji Y-Y, Wang J-W. Antioxidant Properties and PC12 Cell Protective Effects of a Novel Curcumin Analogue (2E,6E)-2,6-Bis(3,5- dimethoxybenzylidene)cyclohexanone (MCH). International Journal of Molecular Sciences. 2014; 15(3):3970-3988. https://doi.org/10.3390/ijms15033970
Chicago/Turabian StyleAo, Gui-Zhen, Xiao-Jing Chu, Yuan-Yuan Ji, and Jian-Wen Wang. 2014. "Antioxidant Properties and PC12 Cell Protective Effects of a Novel Curcumin Analogue (2E,6E)-2,6-Bis(3,5- dimethoxybenzylidene)cyclohexanone (MCH)" International Journal of Molecular Sciences 15, no. 3: 3970-3988. https://doi.org/10.3390/ijms15033970