MADS-Box Transcription Factor SsMADS Is Involved in Regulating Growth and Virulence in Sclerotinia sclerotiorum


Abstract
:1. Introduction
2. Results and Discussion
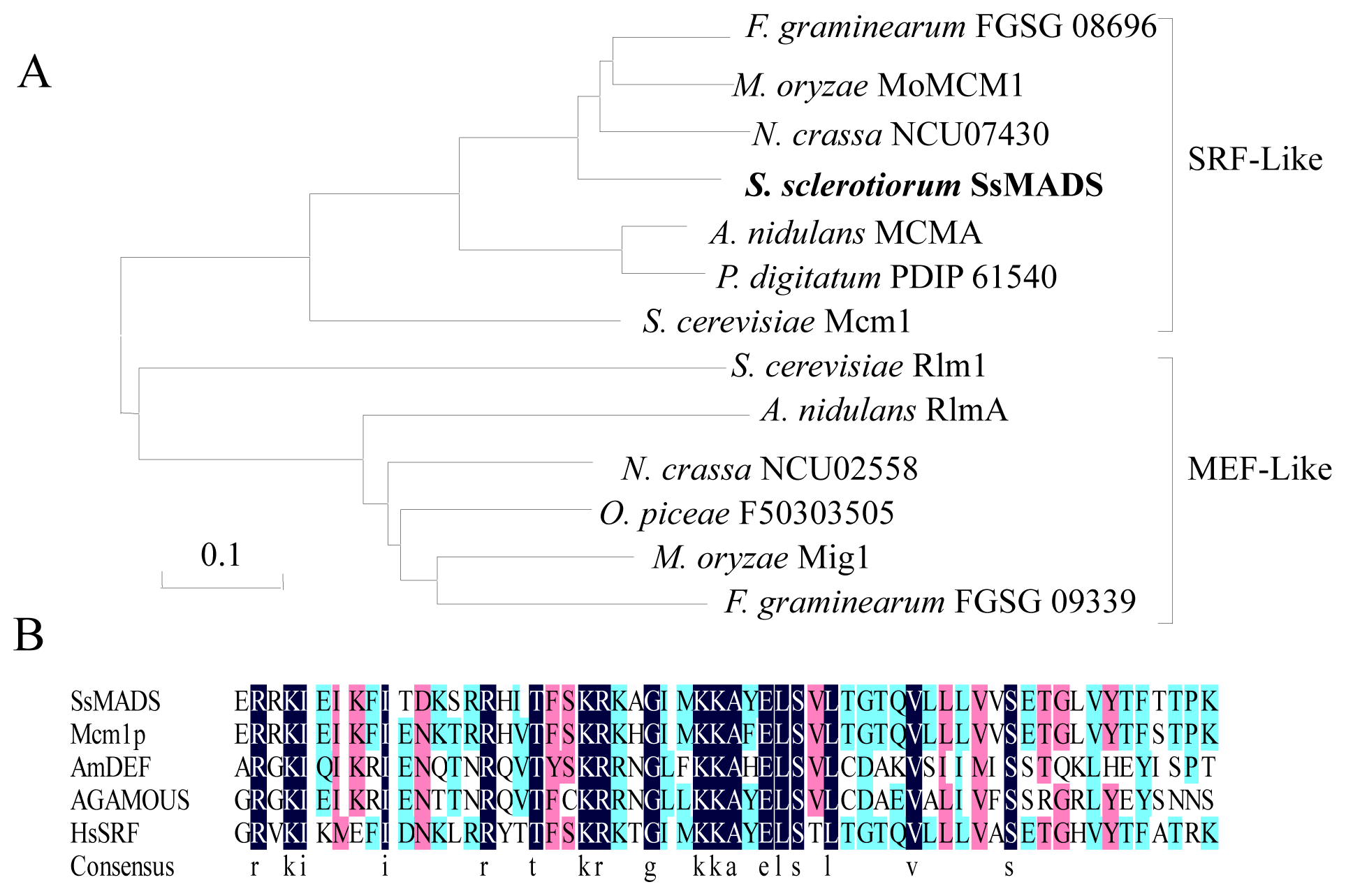
2.1. S. sclerotiorum SsMADS Is a SRF (Serum Response Factor)-MADS-Box Protein
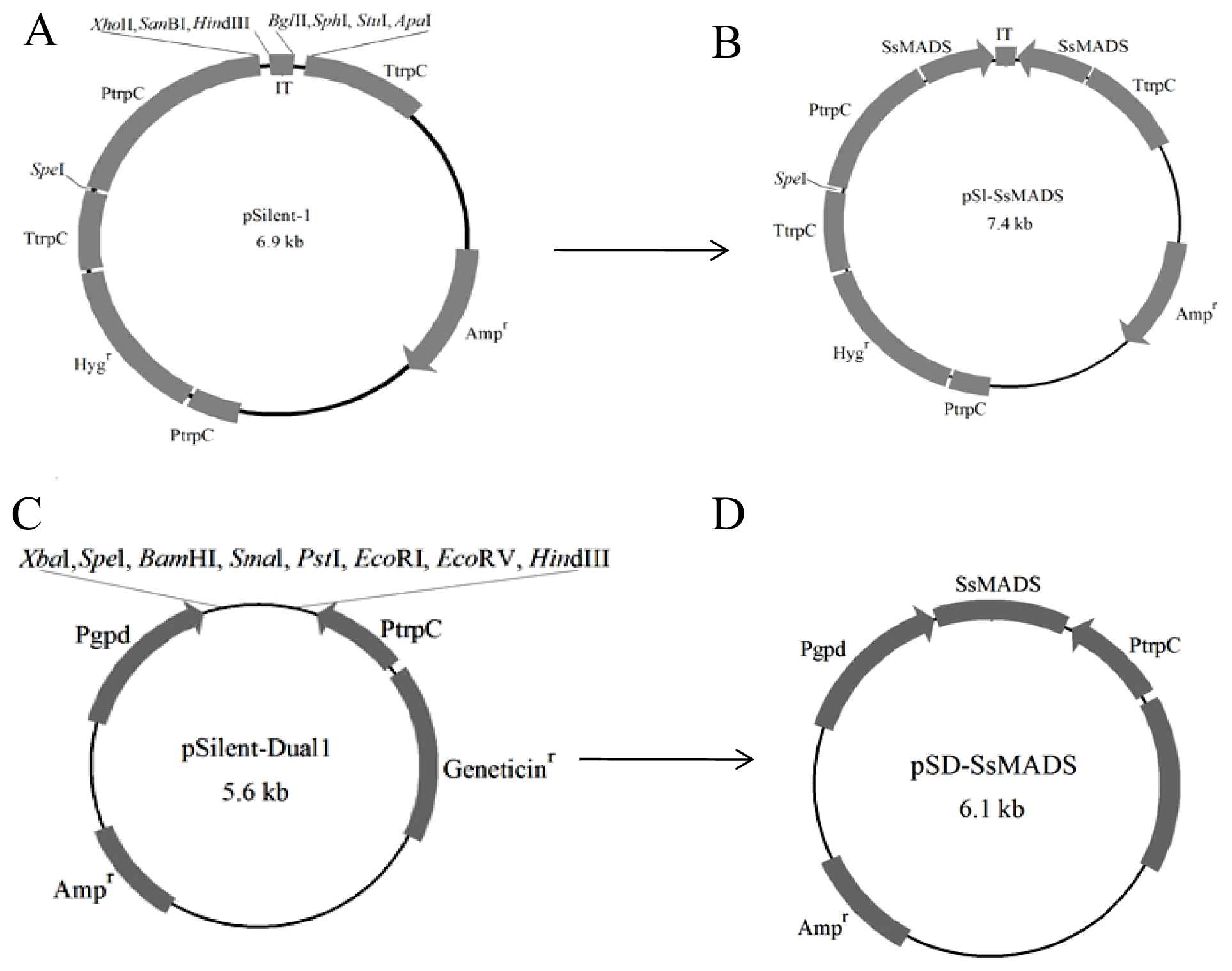
2.2. The RNA Silencing Vectors pS1-SsMADS and pSD-SsMADS Constructed
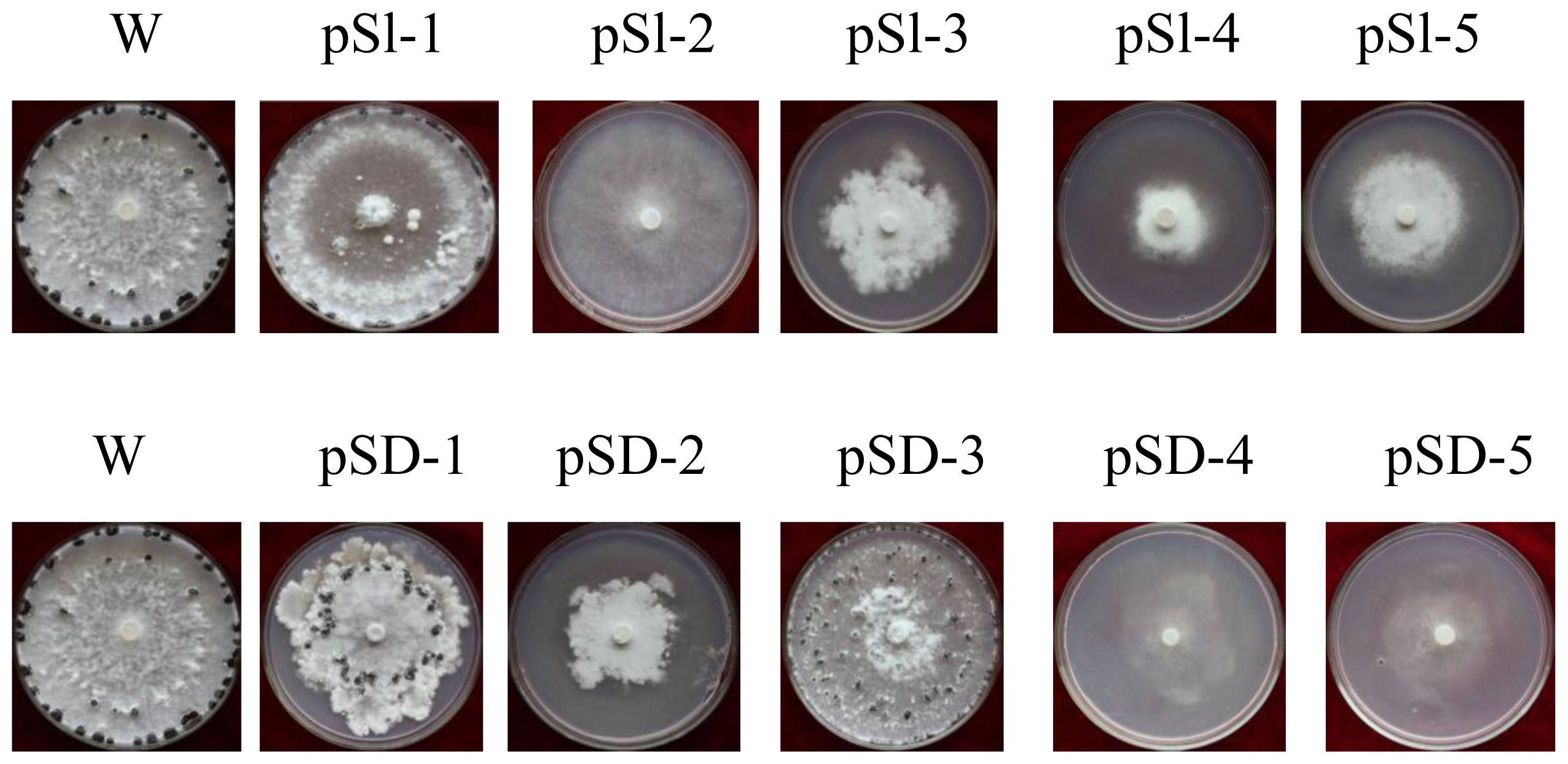
2.3. The Acquisition of RNAi-Positive Transformants
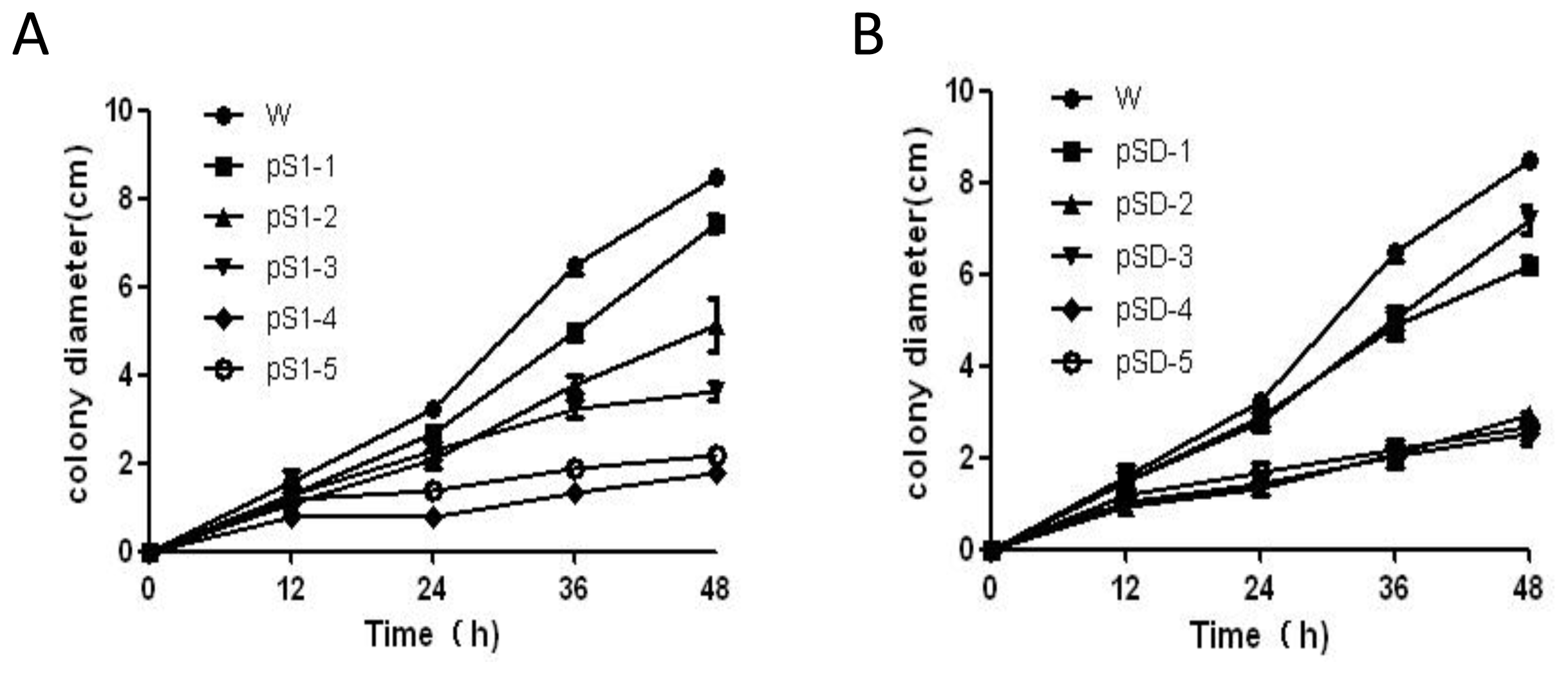
2.4. Silencing of SsMADS Reduced the Aerial Hyphal Growth Rate
2.5. SsMADS Essential for Full Pathogenicity
2.6. Discussion
3. Experimental Section
3.1. Strains and Plasmids
3.2. Cloning of the SsMADS Gene
3.3. Construction of RNAi Reconstructed Plasmids
3.4. Preparation of S. sclerotiorum Protoplasts
3.5. Transformation of Reconstructed Plasmids into S. sclerotiorum
3.6. Screening and Analysis for Phenotypic Transformants
3.7. Detection of Growth Rates
3.8. Real-Time RT-PCR Analysis of SsMADS Gene Expression
3.9. Pathogenicity Tests
4. Conclusions
Acknowledgments
Conflicts of Interest
- Author ContributionsX.Q. and B.Y. performed cloning gene, analyzed data and wrote the paper; J.L., X.Z., and G.L. performed construction plasmids; D.Z., L.L., X.W., and L.W. performed transformation; J.C. and W.M. performed analysis transformants; H.P. and Y.Z. conceived and designed the experiments.
References
- Boland, G.J.; Hall, R. Index of plant hosts of Sclerotinia sclerotiorum. Can. J. Plant Pathol 1994, 16, 93–108. [Google Scholar]
- Dong, T.; Hu, Z.; Deng, L.; Wang, Y.; Zhu, M.; Zhang, J.; Chen, G. A tomato MADS-box transcription factor, SlMADS1, acts as a negative regulator of fruit ripening. Plant Physiol 2013, 163, 1026–1036. [Google Scholar]
- Kofuji, R.; Sumikawa, N.; Yamasaki, M.; Kondo, K.; Ueda, K.; Ito, M.; Hasebe, M. Evolution and divergence of the MADS-box gene family based on genome-wide expression analyses. Mol. Biol. Evol 2003, 20, 1963–1977. [Google Scholar]
- Messenguy, F.; Dubois, E. Role of MADS box proteins and their cofactors in combinatorial control of gene expression and cell development. Gene 2003, 316, 1–21. [Google Scholar]
- Shore, P.; Sharrocks, A.D. The MADS-box family of transcription factors. Eur. J. Biochem 1995, 229, 1–13. [Google Scholar]
- Parenicova, L.; de Folter, S.; Kieffer, M.; Horner, D.S.; Favalli, C.; Busscher, J.; Cook, H.E.; Ingram, R.M.; Kater, M.M.; Davies, B.; et al. Molecular and phylogenetic analyses of the complete MADS-box transcription factor family in Arabidopsis: New openings to the MADS world. Plant Cell 2003, 15, 1538–1551. [Google Scholar]
- Liu, Y.; Cui, S.; Wu, F.; Yan, S.; Lin, X.; Du, X.; Chong, K.; Schilling, S.; Theissen, G.; Meng, Z. Functional conservation of MIKC-Type MADS box genes in Arabidopsis and rice pollen maturation. Plant Cell 2013, 25, 1288–1303. [Google Scholar]
- Perilleux, C.; Pieltain, A.; Jacquemin, G.; Bouche, F.; Detry, N.; D’Aloia, M.; Thiry, L.; Aljochim, P.; Delansnay, M.; Mathieu, A.S.; et al. A root chicory MADS box sequence and the Arabidopsis flowering repressor FLC share common features that suggest conserved function in vernalization and de-vernalization responses. Plant J 2013, 75, 390–402. [Google Scholar]
- Uddenberg, D.; Reimegard, J.; Clapham, D.; Almqvist, C.; von Arnold, S.; Emanuelsson, O.; Sundstrom, J.F. Early cone setting in Picea abies acrocona is associated with increased transcriptional activity of a MADS box transcription factor. Plant Physiol 2013, 161, 813–823. [Google Scholar]
- Treisman, R.; Ammerer, G. The SRF and MCM1 transcription factors. Curr. Opin. Genet. Dev 1992, 2, 221–226. [Google Scholar]
- Wang, D.; Chang, P.S.; Wang, Z.; Sutherland, L.; Richardson, J.A.; Small, E.; Krieg, P.A.; Olson, E.N. Activation of cardiac gene expression by myocardin, a transcriptional cofactor for serum response factor. Cell 2001, 105, 851–862. [Google Scholar]
- Puri, P.L.; Wu, Z.; Zhang, P.; Wood, L.D.; Bhakta, K.S.; Han, J.; Feramisco, J.R.; Karin, M.; Wang, J.Y. Induction of terminal differentiation by constitutive activation of p38 MAP kinase in human rhabdomyosarcoma cells. Genes Dev 2000, 14, 574–584. [Google Scholar]
- Sclerotinia sclerotiorum MADS Box Transcription Factor Gene, Complete CDS. Available online: http://www.ncbi.nlm.nih.gov/nucleotide/%20226938410?report=genbank&log$=nucltop&blast_rank=1&rid=gd6eafn5014 accessed on 22 April, 2009.
- MADS Box Transcription Factor [Sclerotinia sclerotiorum]. Available online: http://www.ncbi.nlm.nih.gov/protein/226938411 accessed on 22 April, 2009.
- Nakayashiki, H.; Hanada, S.; Nguyen, B.Q.; Kadotani, N.; Tosa, Y.; Mayama, S. RNA silencing as a tool for exploring gene function in ascomycete fungi. Fungal Genet. Biol 2005, 42, 275–283. [Google Scholar]
- Nguyen, Q.B.; Kadotani, N.; Kasahara, S.; Tosa, Y.; Mayama, S.; Nakayashiki, H. Systematic functional analysis of calcium-signalling proteins in the genome of the rice-blast fungus, Magnaporthe oryzae, using a high-throughput RNA-silencing system. Mol. Microbiol 2008, 68, 1348–1365. [Google Scholar]
- Alvarez-Buylla, E.R.; Pelaz, S.; Liljegren, S.J.; Gold, S.E.; Burgeff, C.; Ditta, G.S.; de Pouplana, L.R.; Martinez-Castilla, L.; Yanofsky, M.F. An ancestral MADS-box gene duplication occurred before the divergence of plants and animals. Proc. Natl. Acad. Sci. USA 2000, 97, 5328–5333. [Google Scholar]
- Elble, R.; Tye, B.K. Chromosome loss, hyperrecombination, and cell cycle arrest in a yeast mcm1 mutant. Mol. Biol. Cell 1992, 3, 971–980. [Google Scholar]
- McInerny, C.J.; Partridge, J.F.; Mikesell, G.E.; Creemer, D.P.; Breeden, L.L. A novel Mcm1-dependent element in the SWI4, CLN3, CDC6, and CDC47 promoters activates M/G1-specific transcription. Genes Dev 1997, 11, 1277–1288. [Google Scholar]
- Nolting, N.; Poggeler, S. A MADS box protein interacts with a mating-type protein and is required for fruiting body development in the homothallic ascomycete Sordaria macrospora. Eukaryot. Cell 2006, 5, 1043–1056. [Google Scholar]
- Nolting, N.; Poggeler, S. A STE12 homologue of the homothallic ascomycete Sordaria macrospora interacts with the MADS box protein MCM1 and is required for ascosporogenesis. Mol. Microbiol 2006, 62, 853–868. [Google Scholar]
- Ortiz, C.S.; Shim, W.B. The role of MADS-box transcription factors in secondary metabolism and sexual development in the maize pathogen Fusarium verticillioides. Microbiology 2013, 159, 2259–2268. [Google Scholar]
- Zhou, X.; Liu, W.; Wang, C.; Xu, Q.; Wang, Y.; Ding, S.; Xu, J.R. A MADS-box transcription factor MoMcm1 is required for male fertility, microconidium production and virulence in Magnaporthe oryzae. Mol. Microbiol 2011, 80, 33–53. [Google Scholar]
- Park, G.; Xue, C.; Zheng, L.; Lam, S.; Xu, J.R. MST12 regulates infectious growth but not appressorium formation in the rice blast fungus Magnaporthe grisea. Mol. Plant Microbe Interact 2002, 15, 183–192. [Google Scholar]
- Mehrabi, R.; Ding, S.; Xu, J.R. MADS-box transcription factor mig1 is required for infectious growth in Magnaporthe grisea. Eukaryot. Cell 2008, 7, 791–799. [Google Scholar]
- Duncan, K.E.; Howard, R.J. Biology of maize kernel infection by Fusarium verticillioides. Mol. Plant Microbe Interact 2010, 23, 6–16. [Google Scholar]
- Rollins, J.A. The Sclerotinia sclerotiorum pac1 gene is required for sclerotial development and virulence. Mol. Plant Microbe Interact 2003, 16, 785–795. [Google Scholar]
- Zhang, Y.; Li, G.; He, D.; Yu, B.; Yokoyama, K.; Wang, L. Efficient insertional mutagenesis system for the dimorphic pathogenic fungus Sporothrix schenckii using Agrobacterium tumefaciens. J. Microbiol. Methods 2011, 84, 418–422. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′–3′) |
|---|---|
| FP1 | CGAGCTCATGGCCGATATCACAGATCAACACGAC |
| RP2 | CAAGCTTTGATTGATGTGCTTGCGGTTGTGGC |
| pSl-L-1 | CCGCTCGAGGCTCGTGGAATTAAACGCGCAAG (XhoI) |
| pSl-L-2 | CCCAAGCTTGAGCCATATAATTTTGGTACGC (HindIII) |
| pSl-R-1 | GGGGTACCGCTCGTGGAATTAAACGCGCAAG (KpnI) |
| pSl-R-2 | GAAGATCTGAGCCATATAATTTTGGTACGC (BglII) |
| SsSrf-1 | CCCTCTAGAGCTCGTGGAATTAAACGCGCAAG (XbaI) |
| SsSrf-2 | CCCTCTAGAGAGCCATATAATTTTGGTACGC (XbaI) |
| hyg-F | CGACAGCGTCTCCGACCTGA |
| hyg-R | CGCCCAAGCTGCATCATCGAA |
| gen-F | TGTCCGGTGCCCTGAATGAACT |
| gen-R | GCCGCCAAGCTCTTCAGCAATAT |
| kmads11 | GCGTCGCCACATCACATTC |
| kmads12 | TTTCCCTTCCGCCTTTGTG |
| ActF | CCCAGCGTTCTACGTCT |
| ActR | CATGTCAACACGAGCAATG |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Qu, X.; Yu, B.; Liu, J.; Zhang, X.; Li, G.; Zhang, D.; Li, L.; Wang, X.; Wang, L.; Chen, J.; et al. MADS-Box Transcription Factor SsMADS Is Involved in Regulating Growth and Virulence in Sclerotinia sclerotiorum. Int. J. Mol. Sci. 2014, 15, 8049-8062. https://doi.org/10.3390/ijms15058049
Qu X, Yu B, Liu J, Zhang X, Li G, Zhang D, Li L, Wang X, Wang L, Chen J, et al. MADS-Box Transcription Factor SsMADS Is Involved in Regulating Growth and Virulence in Sclerotinia sclerotiorum. International Journal of Molecular Sciences. 2014; 15(5):8049-8062. https://doi.org/10.3390/ijms15058049
Chicago/Turabian StyleQu, Xiaoyan, Baodong Yu, Jinliang Liu, Xianghui Zhang, Guihua Li, Dongjing Zhang, Le Li, Xueliang Wang, Lu Wang, Jingyuan Chen, and et al. 2014. "MADS-Box Transcription Factor SsMADS Is Involved in Regulating Growth and Virulence in Sclerotinia sclerotiorum" International Journal of Molecular Sciences 15, no. 5: 8049-8062. https://doi.org/10.3390/ijms15058049