Interplay between Dioxin-Mediated Signaling and Circadian Clock: A Possible Determinant in Metabolic Homeostasis

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
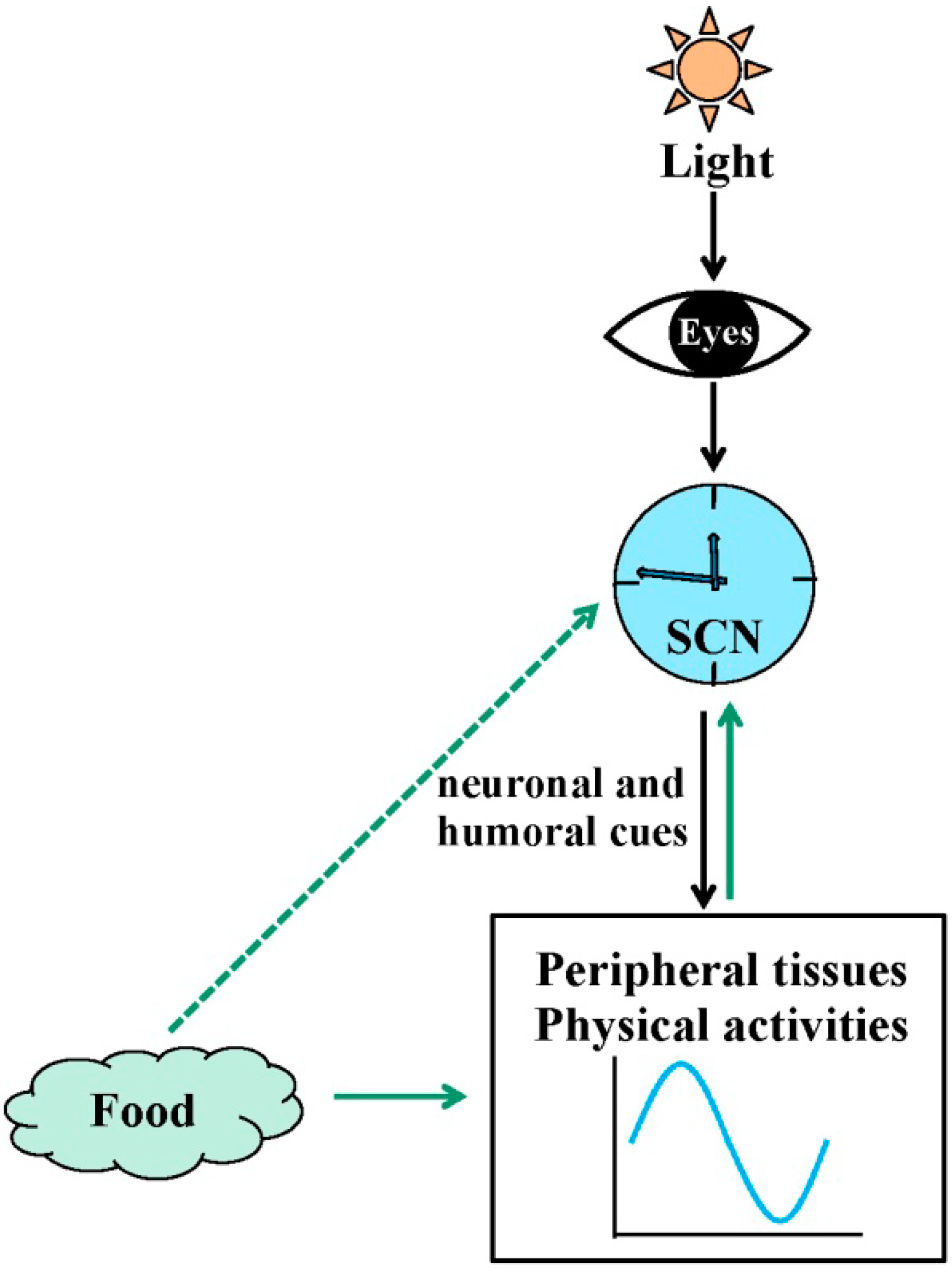
2. Circadian Rhythms in Physiology


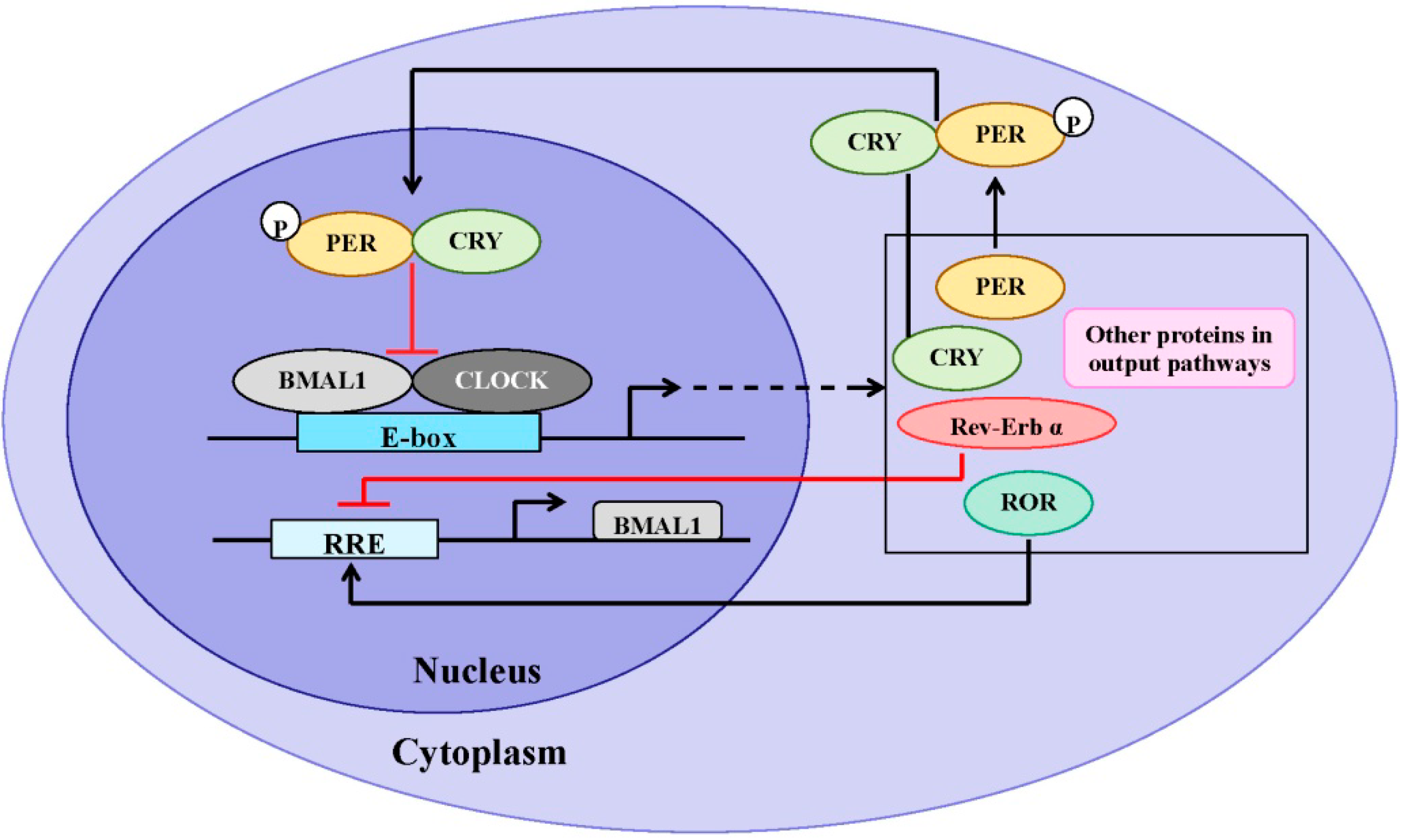
3. Similarities of Structure and Expression Pattern of Dioxin Receptor and Clock Genes
4. Effects of Circadian Clock on AhR Signaling
5. Effects of AhR Signaling on Circadian Rhythm
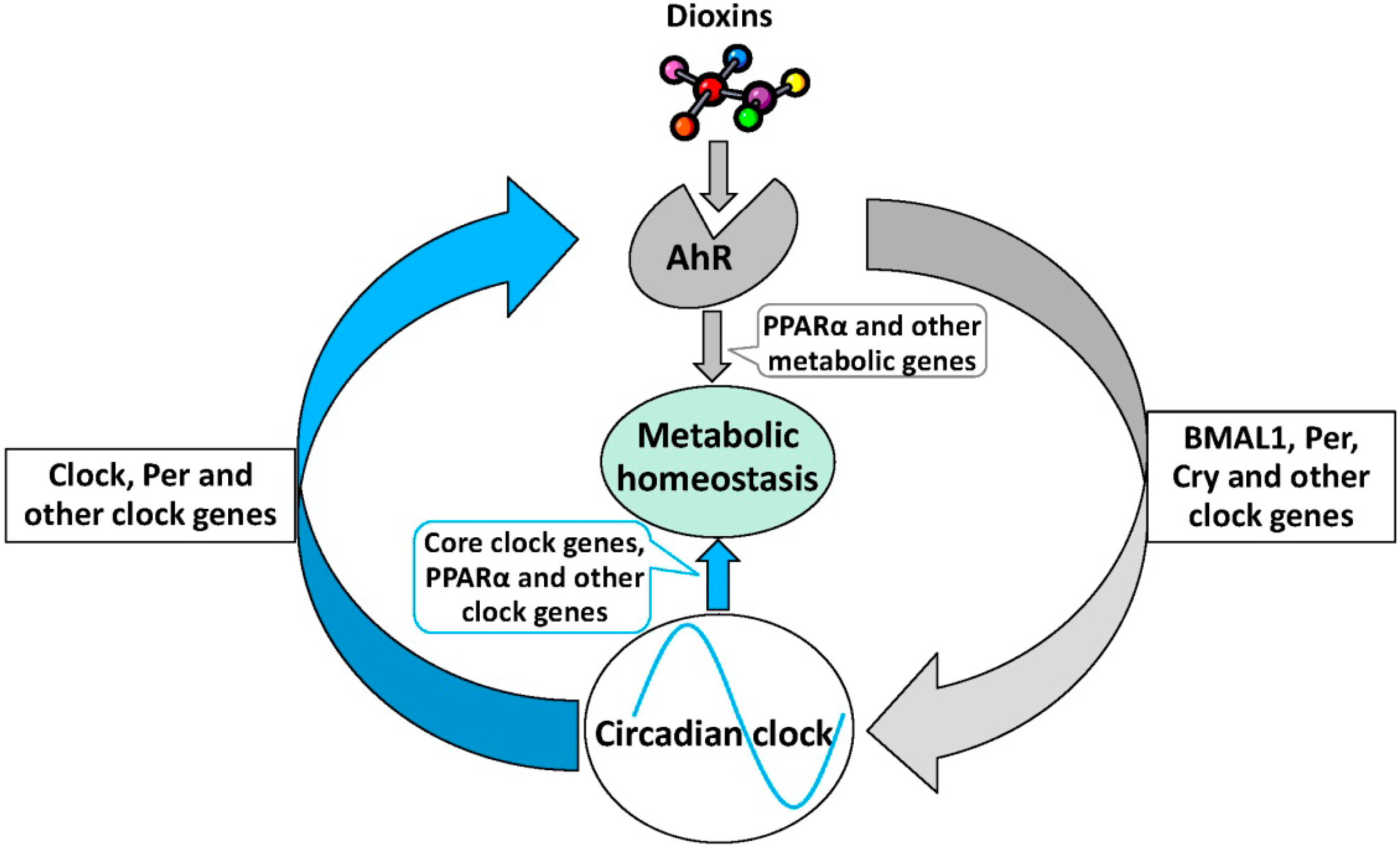
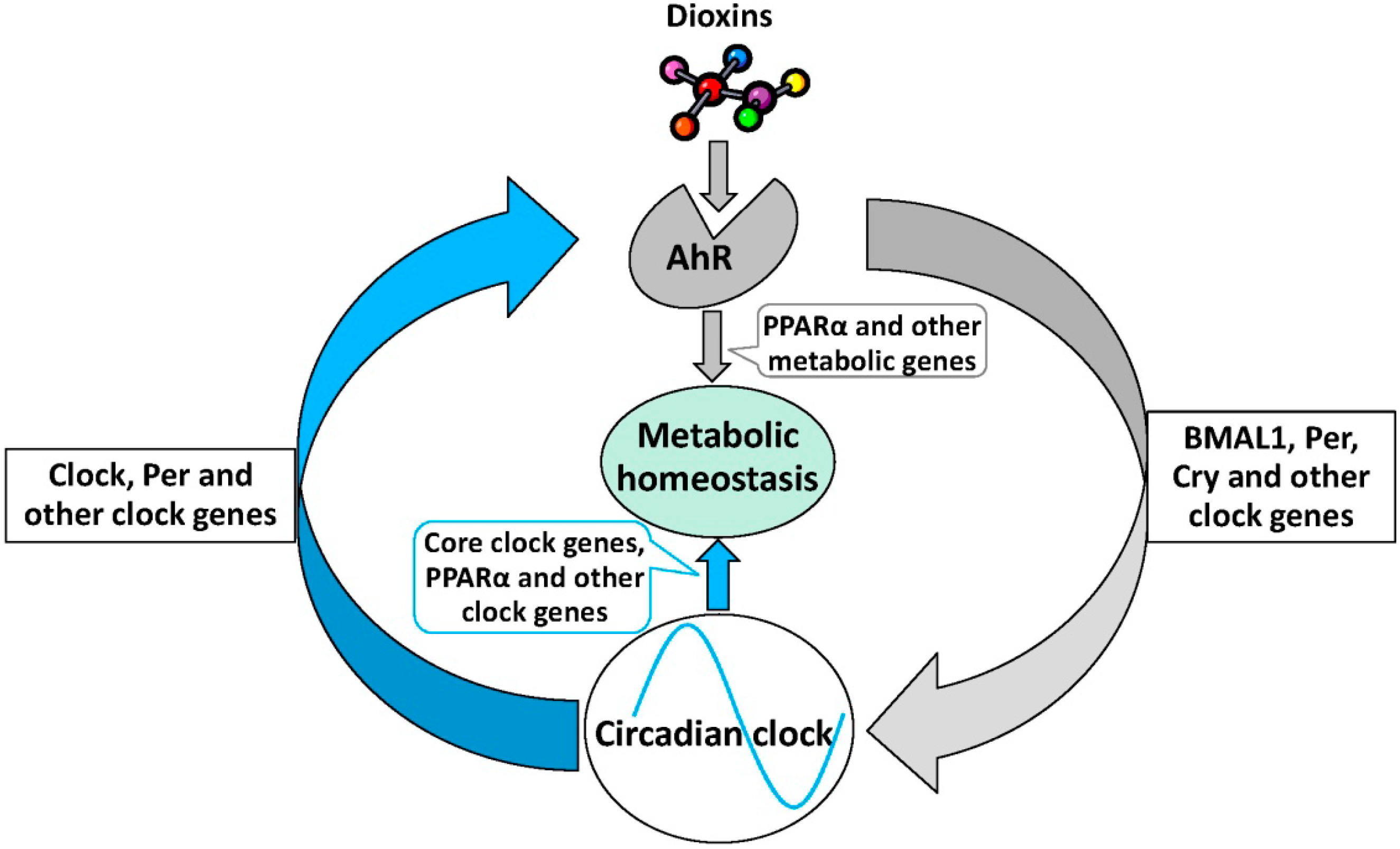
6. The Role of AhR Signaling in Metabolic Homeostasis
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Rey, G.; Reddy, A.B. Connecting cellular metabolism to circadian clocks. Trends Cell Biol. 2013, 23, 234–241. [Google Scholar] [CrossRef]
- Eckel-Mahan, K.; Sassone-Corsi, P. Metabolism and the circadian clock converge. Physiol. Rev. 2013, 93, 107–135. [Google Scholar] [CrossRef]
- Zelinski, E.L.; Deibel, S.H.; McDonald, R.J. The trouble with circadian clock dysfunction: Multiple deleterious effects on the brain and body. Neurosci. Biobehav. Rev. 2014, 40C, 80–101. [Google Scholar] [CrossRef]
- Sahar, S.; Sassone-Corsi, P. Metabolism and cancer: The circadian clock connection. Nat. Rev. Cancer 2009, 9, 886–896. [Google Scholar] [CrossRef]
- Diamanti-Kandarakis, E.; Bourguignon, J.P.; Giudice, L.C.; Hauser, R.; Prins, G.S.; Soto, A.M.; Zoeller, R.T.; Gore, A.C. Endocrine-disrupting chemicals: An Endocrine Society scientific statement. Endocr. Rev. 2009, 30, 293–342. [Google Scholar]
- Soto, A.M.; Sonnenschein, C. Environmental causes of cancer: Endocrine disruptors as carcinogens. Nat. Rev. Endocrinol. 2010, 6, 363–370. [Google Scholar] [CrossRef]
- Thayer, K.A.; Heindel, J.J.; Bucher, J.R.; Gallo, M.A. Role of environmental chemicals in diabetes and obesity: A National Toxicology Program workshop review. Environ. Health Perspect. 2012, 120, 779–789. [Google Scholar] [CrossRef]
- Gu, Y.Z.; Hogenesch, J.B.; Bradfield, C.A. The PAS superfamily: Sensors of environmental and developmental signals. Annu. Rev. Pharmacol. Toxicol. 2000, 40, 519–561. [Google Scholar] [CrossRef]
- McIntosh, B.E.; Hogenesch, J.B.; Bradfield, C.A. Mammalian Per–Arnt–Sim proteins in environmental adaptation. Annu. Rev. Physiol. 2010, 72, 625–645. [Google Scholar] [CrossRef]
- Denison, M.S.; Soshilov, A.A.; He, G.; DeGroot, D.E.; Zhao, B. Exactly the same but different: Promiscuity and diversity in the molecular mechanisms of action of the aryl hydrocarbon (dioxin) receptor. Toxicol. Sci. 2011, 124, 1–22. [Google Scholar] [CrossRef]
- Anderson, G.; Beischlag, T.V.; Vinciguerra, M.; Mazzoccoli, G. The circadian clock circuitry and the AHR signaling pathway in physiology and pathology. Biochem. Pharmacol. 2013, 85, 1405–1416. [Google Scholar] [CrossRef]
- Shimba, S.; Watabe, Y. Crosstalk between the AHR signaling pathway and circadian rhythm. Biochem. Pharmacol. 2009, 77, 560–565. [Google Scholar] [CrossRef]
- Qu, X.; Metz, R.P.; Porter, W.W.; Cassone, V.M.; Earnest, D.J. Disruption of clock gene expression alters responses of the aryl hydrocarbon receptor signaling pathway in the mouse mammary gland. Mol. Pharmacol. 2007, 72, 1349–1358. [Google Scholar] [CrossRef]
- Qu, X.; Metz, R.P.; Porter, W.W.; Neuendorff, N.; Earnest, B.J.; Earnest, D.J. The clock genes period 1 and period 2 mediate diurnal rhythms in dioxin-induced Cyp1A1 expression in the mouse mammary gland and liver. Toxicol. Lett. 2010, 196, 28–32. [Google Scholar] [CrossRef]
- Partch, C.L.; Green, C.B.; Takahashi, J.S. Molecular architecture of the mammalian circadian clock. Trends Cell Biol. 2014, 24, 90–99. [Google Scholar] [CrossRef]
- Dibner, C.; Schibler, U.; Albrecht, U. The mammalian circadian timing system: Organization and coordination of central and peripheral clocks. Annu. Rev. Physiol. 2010, 72, 517–549. [Google Scholar]
- Boulos, Z.; Terman, M. Food availability and daily biological rhythms. Neurosci. Biobehav. Rev. 1980, 4, 119–131. [Google Scholar] [CrossRef]
- Mistlberger, R.E. Circadian food-anticipatory activity: Formal models and physiological mechanisms. Neurosci. Biobehav. Rev. 1994, 18, 171–195. [Google Scholar] [CrossRef]
- Stephan, F.K. The “other” circadian system: Food as a Zeitgeber. J. Biol. Rhythm. 2002, 17, 284–292. [Google Scholar]
- Brown, S.A.; Zumbrunn, G.; Fleury-Olela, F.; Preitner, N.; Schibler, U. Rhythms of mammalian body temperature can sustain peripheral circadian clocks. Curr. Biol. 2002, 12, 1574–1583. [Google Scholar] [CrossRef]
- Mendoza, J. Circadian clocks: Setting time by food. J. Neuroendocrinol. 2007, 19, 127–137. [Google Scholar] [CrossRef]
- Langner, R.; Rensing, L. Circadian rhythm of oxygen consumption in rat liver suspension culture: Changes of pattern. Z. Naturforschung Teil B 1972, 27, 1117–1118. [Google Scholar]
- Yoo, S.H.; Yamazaki, S.; Lowrey, P.L.; Shimomura, K.; Ko, C.H.; Buhr, E.D.; Siepka, S.M.; Hong, H.K.; Oh, W.J.; Yoo, O.J.; et al. PERIOD2::LUCIFERASE real-time reporting of circadian dynamics reveals persistent circadian oscillations in mouse peripheral tissues. Proc. Natl. Acad. Sci. USA 2004, 101, 5339–5346. [Google Scholar] [CrossRef]
- Richards, J.; Gumz, M.L. Mechanism of the circadian clock in physiology. Am. J. Physiol. 2013, 304, R1053–R1064. [Google Scholar] [CrossRef]
- Gekakis, N.; Staknis, D.; Nguyen, H.B.; Davis, F.C.; Wilsbacher, L.D.; King, D.P.; Takahashi, J.S.; Weitz, C.J. Role of the CLOCK protein in the mammalian circadian mechanism. Science 1998, 280, 1564–1569. [Google Scholar] [CrossRef]
- Zhang, E.E.; Kay, S.A. Clocks not winding down: Unravelling circadian networks. Nat. Rev. Mol. Cell Biol. 2010, 11, 764–776. [Google Scholar] [CrossRef]
- Ripperger, J.A.; Shearman, L.P.; Reppert, S.M.; Schibler, U. CLOCK, an essential pacemaker component, controls expression of the circadian transcription factor DBP. Genes Dev. 2000, 14, 679–689. [Google Scholar]
- Jin, X.; Shearman, L.P.; Weaver, D.R.; Zylka, M.J.; de Vries, G.J.; Reppert, S.M. A molecular mechanism regulating rhythmic output from the suprachiasmatic circadian clock. Cell 1999, 96, 57–68. [Google Scholar] [CrossRef]
- Cheng, M.Y.; Bullock, C.M.; Li, C.; Lee, A.G.; Bermak, J.C.; Belluzzi, J.; Weaver, D.R.; Leslie, F.M.; Zhou, Q.Y. Prokineticin 2 transmits the behavioural circadian rhythm of the suprachiasmatic nucleus. Nature 2002, 417, 405–410. [Google Scholar] [CrossRef]
- Oishi, K.; Shirai, H.; Ishida, N. CLOCK is involved in the circadian transactivation of peroxisome-proliferator-activated receptor alpha (PPARα) in mice. Biochem. J. 2005, 386, 575–581. [Google Scholar] [CrossRef]
- Canaple, L.; Rambaud, J.; Dkhissi-Benyahya, O.; Rayet, B.; Tan, N.S.; Michalik, L.; Delaunay, F.; Wahli, W.; Laudet, V. Reciprocal regulation of brain and muscle Arnt-like protein 1 and peroxisome proliferator-activated receptor α defines a novel positive feedback loop in the rodent liver circadian clock. Mol. Endocrinol. 2006, 20, 1715–1727. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.K.; Yeager, R.L.; Klaassen, C.D. Circadian expression profiles of drug-processing genes and transcription factors in mouse liver. Drug Metab. Dispos. 2009, 37, 106–115. [Google Scholar] [CrossRef]
- Antoch, M.P.; Song, E.J.; Chang, A.M.; Vitaterna, M.H.; Zhao, Y.; Wilsbacher, L.D.; Sangoram, A.M.; King, D.P.; Pinto, L.H.; Takahashi, J.S. Functional identification of the mouse circadian Clock gene by transgenic BAC rescue. Cell 1997, 89, 655–667. [Google Scholar] [CrossRef]
- Ikeda, M.; Nomura, M. cDNA cloning and tissue-specific expression of a novel basic helix-loop-helix/PAS protein (BMAL1) and identification of alternatively spliced variants with alternative translation initiation site usage. Biochem. Biophys. Res. Commun. 1997, 233, 258–264. [Google Scholar] [CrossRef]
- Reddy, P.; Jacquier, A.C.; Abovich, N.; Petersen, G.; Rosbash, M. The period clock locus of D. melanogaster codes for a proteoglycan. Cell 1986, 46, 53–61. [Google Scholar] [CrossRef]
- Hoffman, E.C.; Reyes, H.; Chu, F.F.; Sander, F.; Conley, L.H.; Brooks, B.A.; Hankinson, O. Cloning of a factor required for activity of the Ah (dioxin) receptor. Science 1991, 252, 954–958. [Google Scholar]
- Nambu, J.R.; Lewis, J.O.; Wharton, K.A., Jr.; Crews, S.T. The Drosophila single-minded gene encodes a helix-loop-helix protein that acts as a master regulator of CNS midline development. Cell 1991, 67, 1157–1167. [Google Scholar] [CrossRef]
- Itoh, S.; Kamataki, T. Human Ah receptor cDNA: Analysis for highly conserved sequences. Nucleic Acids Res. 1993, 21, 3578. [Google Scholar] [CrossRef]
- Takahata, S.; Sogawa, K.; Kobayashi, A.; Ema, M.; Mimura, J.; Ozaki, N.; Fujii-Kuriyama, Y. Transcriptionally active heterodimer formation of an Arnt-like PAS protein, Arnt3, with HIF-1a, HLF, and Clock. Biochem. Biophys. Res. Commun. 1998, 248, 789–794. [Google Scholar] [CrossRef]
- Xu, C.X.; Krager, S.L.; Liao, D.F.; Tischkau, S.A. Disruption of CLOCK-BMAL1 transcriptional activity is responsible for aryl hydrocarbon receptor-mediated regulation of Period1 gene. Toxicol. Sci. 2010, 115, 98–108. [Google Scholar] [CrossRef]
- Hogenesch, J.B.; Gu, Y.Z.; Jain, S.; Bradfield, C.A. The basic-helix-loop-helix-PAS orphan MOP3 forms transcriptionally active complexes with circadian and hypoxia factors. Proc. Natl. Acad. Sci. USA 1998, 95, 5474–5479. [Google Scholar] [CrossRef]
- Abbott, B.D.; Birnbaum, L.S.; Perdew, G.H. Developmental expression of two members of a new class of transcription factors: I. Expression of aryl hydrocarbon receptor in the C57BL/6N mouse embryo. Dev. Dyn. 1995, 204, 133–143. [Google Scholar] [CrossRef]
- Li, W.; Donat, S.; Dohr, O.; Unfried, K.; Abel, J. Ah receptor in different tissues of C57BL/6J and DBA/2J mice: Use of competitive polymerase chain reaction to measure Ah-receptor mRNA expression. Arch. Biochem. Biophys. 1994, 315, 279–284. [Google Scholar] [CrossRef]
- Richardson, V.M.; Santostefano, M.J.; Birnbaum, L.S. Daily cycle of bHLH-PAS proteins, Ah receptor and Arnt, in multiple tissues of female Sprague-Dawley rats. Biochem. Biophys. Res. Commun. 1998, 252, 225–231. [Google Scholar]
- Huang, P.; Ceccatelli, S.; Rannug, A. A study on diurnal mRNA expression of CYP1A1, AHR, ARNT, and PER2 in rat pituitary and liver. Environ. Toxicol. Pharmacol. 2002, 11, 119–126. [Google Scholar] [CrossRef]
- Mukai, M.; Lin, T.M.; Peterson, R.E.; Cooke, P.S.; Tischkau, S.A. Behavioral rhythmicity of mice lacking AhR and attenuation of light-induced phase shift by 2,3,7,8-tetrachlorodibenzo-p-dioxin. J. Biol. Rhythm. 2008, 23, 200–210. [Google Scholar] [CrossRef]
- Qu, X.; Metz, R.P.; Porter, W.W.; Cassone, V.M.; Earnest, D.J. Disruption of period gene expression alters the inductive effects of dioxin on the AhR signaling pathway in the mouse liver. Toxicol. Appl. Pharmacol. 2009, 234, 370–377. [Google Scholar] [CrossRef]
- Tanimura, N.; Kusunose, N.; Matsunaga, N.; Koyanagi, S.; Ohdo, S. Aryl hydrocarbon receptor-mediated Cyp1a1 expression is modulated in a CLOCK-dependent circadian manner. Toxicology 2011, 290, 203–207. [Google Scholar] [CrossRef]
- Garrison, P.M.; Denison, M.S. Analysis of the murine AhR gene promoter. J. Biochem. Mol. Toxicol. 2000, 14, 1–10. [Google Scholar]
- Xu, C.X.; Wang, C.; Krager, S.L.; Bottum, K.M.; Tischkau, S.A. Aryl hydrocarbon receptor activation attenuates Per1 gene induction and influences circadian clock resetting. Toxicol. Sci. 2013, 132, 368–378. [Google Scholar] [CrossRef]
- Mukai, M.; Tischkau, S.A. Effects of tryptophan photoproducts in the circadian timing system: Searching for a physiological role for aryl hydrocarbon receptor. Toxicol. Sci. 2007, 95, 172–181. [Google Scholar]
- Solak, K.A.; Wijnolts, F.M.; Pralong, F.P.; Blaauboer, B.J.; van den Berg, M.; Westerink, R.H.; van Duursen, M.B. In vitro neuroendocrine effects of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) in the AhR-expressing hypothalamic rat GnV-3 cell line. Toxicology 2013, 311, 124–134. [Google Scholar] [CrossRef]
- Sato, S.; Shirakawa, H.; Tomita, S.; Ohsaki, Y.; Haketa, K.; Tooi, O.; Santo, N.; Tohkin, M.; Furukawa, Y.; Gonzalez, F.J.; et al. Low-dose dioxins alter gene expression related to cholesterol biosynthesis, lipogenesis, and glucose metabolism through the aryl hydrocarbon receptor-mediated pathway in mouse liver. Toxicol. Appl. Pharmacol. 2008, 229, 10–19. [Google Scholar]
- Tischkau, S.A.; Jaeger, C.D.; Krager, S.L. Circadian clock disruption in the mouse ovary in response to 2,3,7,8-tetrachlorodibenzo-p-dioxin. Toxicol. Lett. 2011, 201, 116–122. [Google Scholar] [CrossRef]
- Garrett, R.W.; Gasiewicz, T.A. The aryl hydrocarbon receptor agonist 2,3,7,8-tetrachlorodibenzo-p-dioxin alters the circadian rhythms, quiescence, and expression of clock genes in murine hematopoietic stem and progenitor cells. Mol. Pharmacol. 2006, 69, 2076–2083. [Google Scholar] [CrossRef]
- Malik, A.I.; Rowan-Carroll, A.; Williams, A.; Lemieux, C.L.; Long, A.S.; Arlt, V.M.; Phillips, D.H.; White, P.A.; Yauk, C.L. Hepatic genotoxicity and toxicogenomic responses in MutaMouse males treated with dibenz [a,h] anthracene. Mutagenesis 2013, 28, 543–554. [Google Scholar] [CrossRef]
- Pendergast, J.S.; Yamazaki, S. The mammalian circadian system is resistant to dioxin. J. Biol. Rhythm. 2012, 27, 156–163. [Google Scholar] [CrossRef]
- Wang, C.; Xu, C.X.; Krager, S.L.; Bottum, K.M.; Liao, D.F.; Tischkau, S.A. Aryl hydrocarbon receptor deficiency enhances insulin sensitivity and reduces PPAR-α pathway activity in mice. Environ. Health Perspect. 2011, 119, 1739–1744. [Google Scholar]
- Kelling, C.K.; Christian, B.J.; Inhorn, S.L.; Peterson, R.E. Hypophagia-induced weight loss in mice, rats, and guinea pigs treated with 2,3,7,8-tetrachlorodibenzo-p-dioxin. Fundam. Appl. Toxicol. 1985, 5, 700–712. [Google Scholar] [CrossRef]
- Seefeld, M.D.; Corbett, S.W.; Keesey, R.E.; Peterson, R.E. Characterization of the wasting syndrome in rats treated with 2,3,7,8-tetrachlorodibenzo-p-dioxin. Toxicol. Appl. Pharmacol. 1984, 73, 311–322. [Google Scholar] [CrossRef]
- Jones, M.K.; Weisenburger, W.P.; Sipes, I.G.; Russell, D.H. Circadian alterations in prolactin, corticosterone, and thyroid hormone levels and down-regulation of prolactin receptor activity by 2,3,7,8-tetrachlorodibenzo-p-dioxin. Toxicol. Appl. Pharmacol. 1987, 87, 337–350. [Google Scholar] [CrossRef]
- Pohjanvirta, R.; Tuomisto, J.; Linden, J.; Laitinen, J. TCDD reduces serum melatonin levels in Long-Evans rats. Pharmacol. Toxicol. 1989, 65, 239–240. [Google Scholar] [CrossRef]
- Yellon, S.M.; Singh, D.; Garrett, T.M.; Fagoaga, O.R.; Nehlsen-Cannarella, S.L. Reproductive, neuroendocrine, and immune consequences of acute exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin in the Siberian hamster. Biol. Reprod. 2000, 63, 538–543. [Google Scholar] [CrossRef]
- Gentner, N.J.; Weber, L.P. Intranasal benzo [a] pyrene alters circadian blood pressure patterns and causes lung inflammation in rats. Arch. Toxicol. 2011, 85, 337–346. [Google Scholar] [CrossRef]
- Dugo, M.A.; Han, F.; Tchounwou, P.B. Persistent polar depletion of stratospheric ozone and emergent mechanisms of ultraviolet radiation-mediated health dysregulation. Rev. Environ. Health 2012, 27, 103–116. [Google Scholar]
- Ma, Q. Influence of light on aryl hydrocarbon receptor signaling and consequences in drug metabolism, physiology and disease. Expert Opin. Drug Metab. Toxicol. 2011, 7, 1267–1293. [Google Scholar] [CrossRef]
- Arisawa, K.; Takeda, H.; Mikasa, H. Background exposure to PCDDs/PCDFs/PCBs and its potential health effects: A review of epidemiologic studies. J. Med. Investig. 2005, 52, 10–21. [Google Scholar] [CrossRef]
- Bertazzi, P.A.; Bernucci, I.; Brambilla, G.; Consonni, D.; Pesatori, A.C. The Seveso studies on early and long-term effects of dioxin exposure: A review. Environ. Health Perspect. 1998, 106, 625–633. [Google Scholar]
- Larsen, J.C. Risk assessments of polychlorinated dibenzo-p-dioxins, polychlorinated dibenzofurans, and dioxin-like polychlorinated biphenyls in food. Mol. Nutr. Food Res. 2006, 50, 885–896. [Google Scholar]
- Henriksen, G.L.; Ketchum, N.S.; Michalek, J.E.; Swaby, J.A. Serum dioxin and diabetes mellitus in veterans of Operation Ranch Hand. Epidemiology 1997, 8, 252–258. [Google Scholar] [CrossRef]
- Cui, J.; Li, S. Inhibitors and prodrugs targeting CYP1: A novel approach in cancer prevention and therapy. Curr. Med. Chem. 2014, 21, 519–552. [Google Scholar] [CrossRef]
- Korkina, L.G.; Pastore, S.; Dellambra, E.; de Luca, C. New molecular and cellular targets for chemoprevention and treatment of skin tumors by plant polyphenols: A critical review. Curr. Med. Chem. 2013, 20, 852–868. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, C.; Zhang, Z.-M.; Xu, C.-X.; Tischkau, S.A. Interplay between Dioxin-Mediated Signaling and Circadian Clock: A Possible Determinant in Metabolic Homeostasis. Int. J. Mol. Sci. 2014, 15, 11700-11712. https://doi.org/10.3390/ijms150711700
Wang C, Zhang Z-M, Xu C-X, Tischkau SA. Interplay between Dioxin-Mediated Signaling and Circadian Clock: A Possible Determinant in Metabolic Homeostasis. International Journal of Molecular Sciences. 2014; 15(7):11700-11712. https://doi.org/10.3390/ijms150711700
Chicago/Turabian StyleWang, Chun, Zhi-Ming Zhang, Can-Xin Xu, and Shelley A. Tischkau. 2014. "Interplay between Dioxin-Mediated Signaling and Circadian Clock: A Possible Determinant in Metabolic Homeostasis" International Journal of Molecular Sciences 15, no. 7: 11700-11712. https://doi.org/10.3390/ijms150711700