The Involvement of miR-23a/APAF1 Regulation Axis in Colorectal Cancer

Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
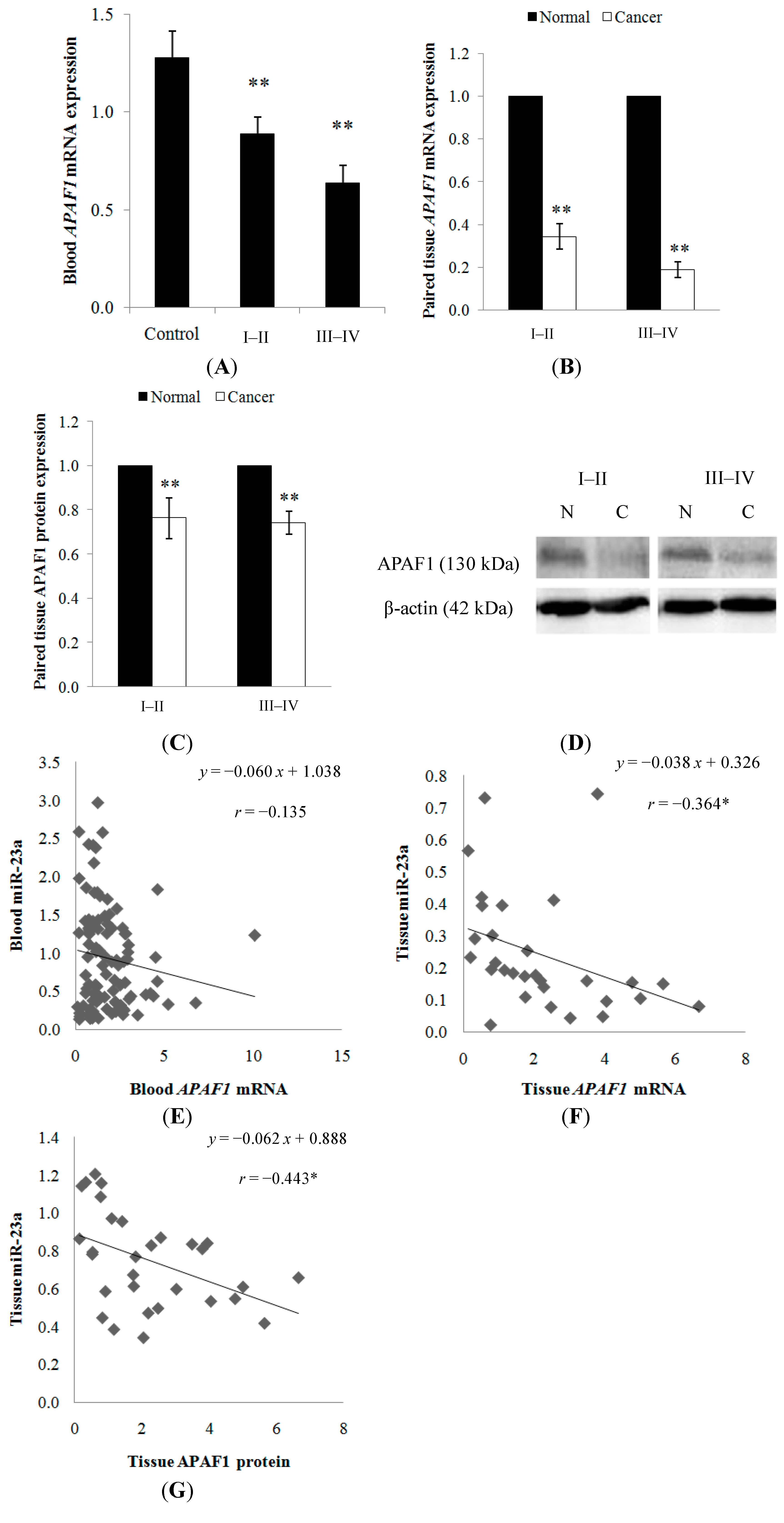
2.1.1. miR-23a Up-Regulation in Clinical CRC Samples

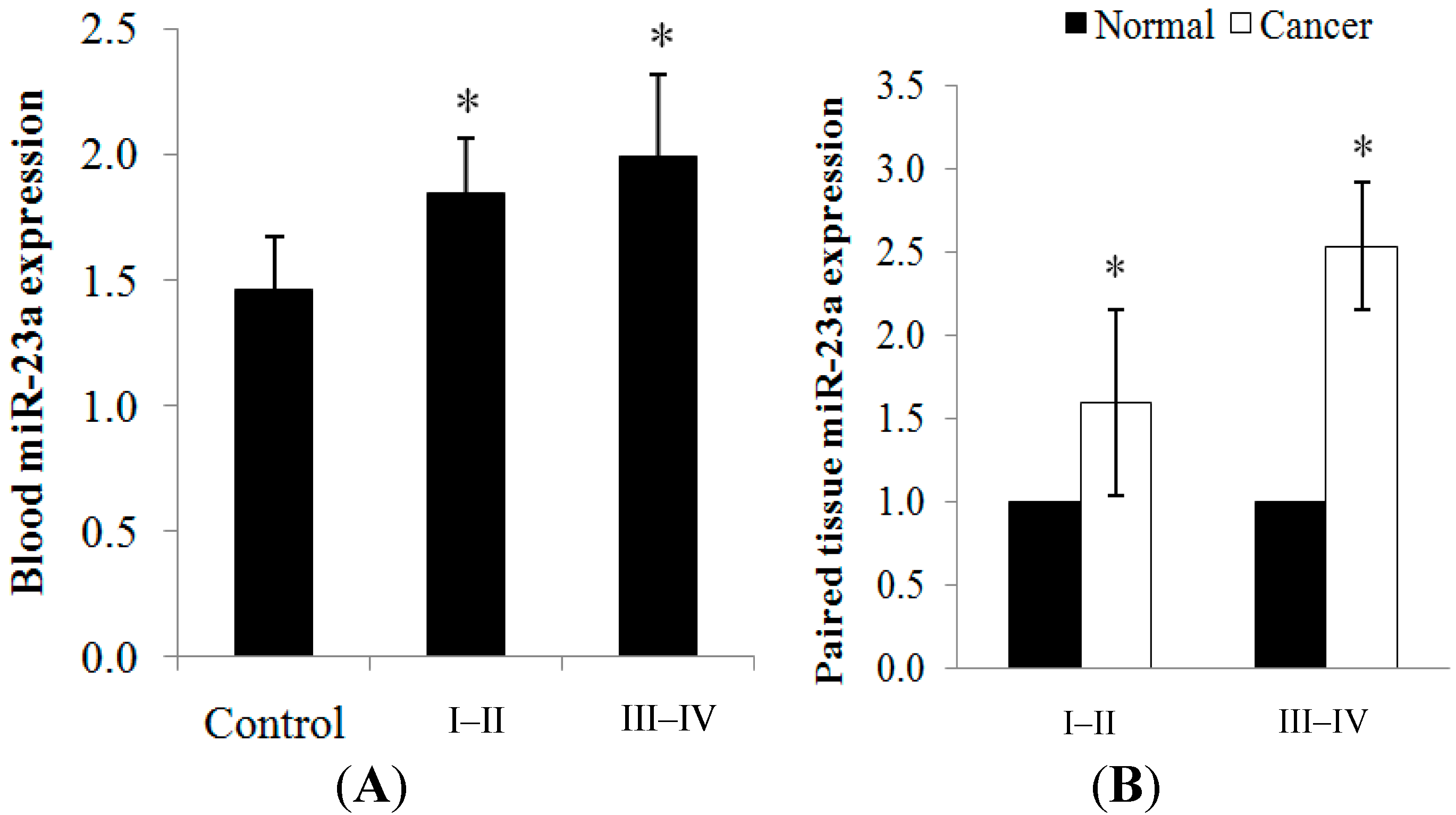{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Healthy Control (n = 32), n (%) | CRC Blood (n = 70), n (%) | Subset of Paired CRC Tissue (n = 30), n (%) | |
|---|---|---|---|---|
| Age (years) | 58.0 ± 11.1 | 65.1 ± 9.6 | 65.2 ± 9.8 | |
| Gender | Male | 19 (59.4%) | 54 (77.1%) | 19 (63.3%) |
| Female | 13 (40.6%) | 16 (22.9%) | 11 (36.7%) | |
| Ethnicity | Asian | 32 (100%) | 70 (100.0%) | 30 (100%) |
| TNM stage | I–II | 39 (55.7%) | 10 (33.3%) | |
| III–IV | 31 (44.3%) | 20 (66.7%) | ||
| Tumor location | Colon | 42 (60.0%) | 30 (100%) | |
| Rectum | 28 (40.0%) | 0 | ||
| Tumor grading (adenocarcinoma) | G1 | 25 (35.7%) | 13 (43.3%) | |
| G2 | 34 (48.6%) | 13 (43.3%) | ||
| G3 | 11 (15.7%) | 4 (13.3%) | ||

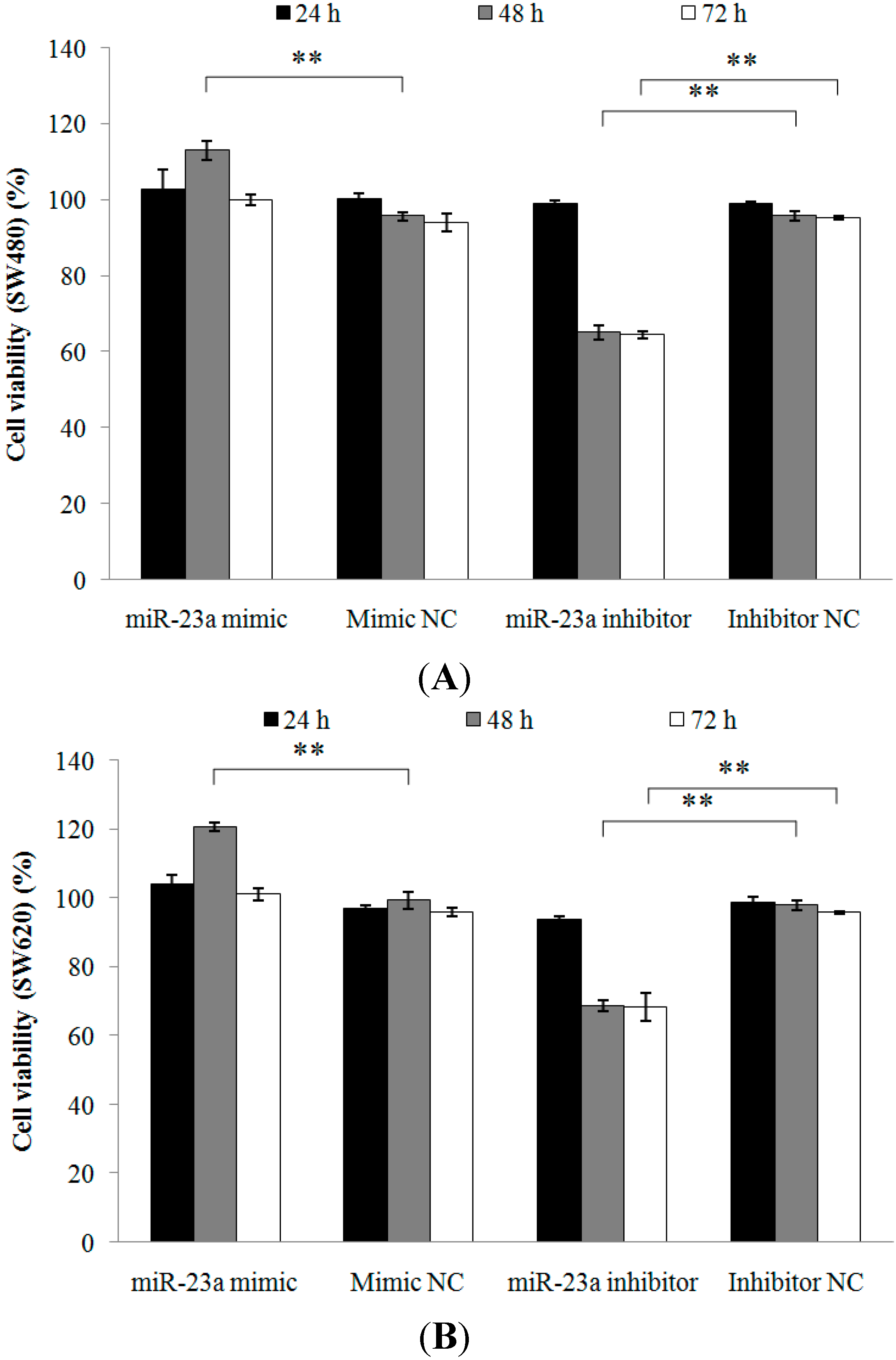
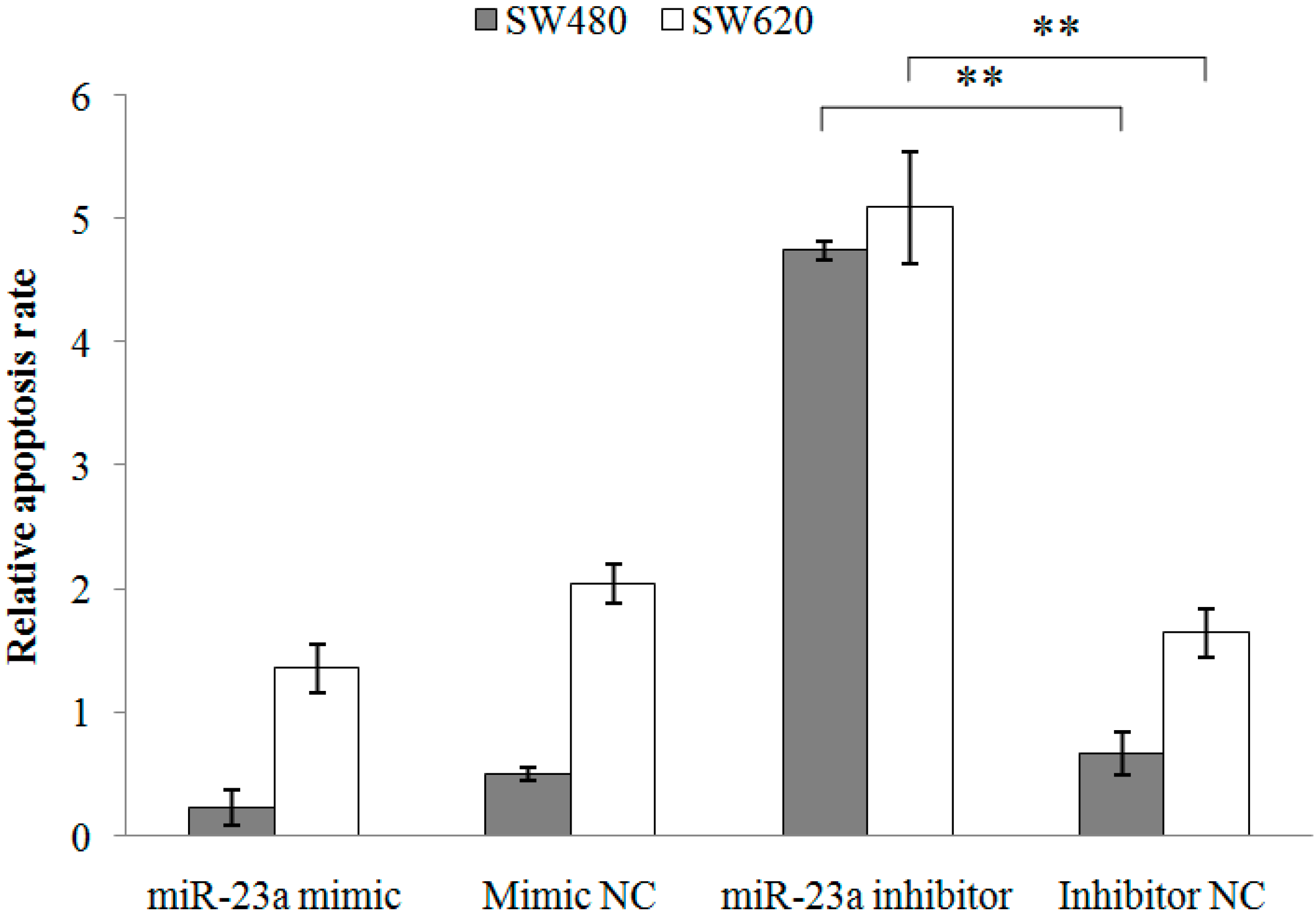
2.1.2. Inhibition of miR-23a in CRC Cell Lines Reduces Cell Viability, Promotes Cell Apoptosis and Increases Caspase Activation

| Treatment | SW480 | SW620 | ||||
|---|---|---|---|---|---|---|
| Live (%) | Dead (%) | Apoptotic (%) | Live (%) | Dead (%) | Apoptotic (%) | |
| NTC | 84.33 ± 0.33 | 10.33 ± 0.33 | 5.33 ± 0.33 | 84.00 ± 2.00 | 12.33 ± 1.76 | 3.67 ± 0.67 |
| Mock | 87.00 ± 1.53 | 11.00 ± 0.00 | 2.00 ± 1.53 | 81.67 ± 0.67 | 16.00 ± 0.58 | 2.00 ± 0.00 |
| miR-23a mimic | 85.33 ± 1.20 | 13.00 ± 1.00 | 1.33 ± 0.88 | 85.00 ± 1.15 | 10.00 ± 0.00 | 5.00 ± 1.15 |
| Mimic NC | 83.67 ± 0.88 | 13.00 ± 0.58 | 2.67 ± 0.33 | 78.33 ± 0.88 | 14.00 ± 0.58 | 7.33 ± 0.88 |
| miR-23a inhibitor | 65.67 ±1.45 | 9.00 ± 1.00 | 25.33 ± 1.86 | 68.00 ± 1.73 | 13.33 ± 0.67 | 18.33 ± 2.60 |
| Inhibitor NC | 85.00 ± 1.53 | 11.67 ± 0.67 | 3.67 ± 1.20 | 78.67 ± 1.20 | 15.00 ± 0.58 | 6.00 ± 1.15 |
| Doxorubicin | 54.33 ± 4.67 | 23.67 ± 0.88 | 22.33 ± 5.33 | 48.33 ± 4.84 | 27.00 ± 2.08 | 24.33 ± 2.91 |


2.1.3. Functional Interaction between miR-23a and 3'-UTR of Apoptotic Peptidase Activating Factor 1 (APAF1) mRNA
| APAF1 3'-UTR (nt 1474–1481) | 5' ...AAGAUUUUUCUAAGAAAUGUGAA... |
|---|---|
| | | | | | | | | |
| hsa-miR-23a | 3' CCUUUAGGGACCGUUACACUA |
| Mutated APAF1 3'-UTR (nt 1474−1481) | 5' ...AAGAUUUUUCUAAGAAUUCUGCA... |
| | † | † | | † | |
| hsa-miR-23a | 3' CCUUUAGGGACCGUUACACUA |

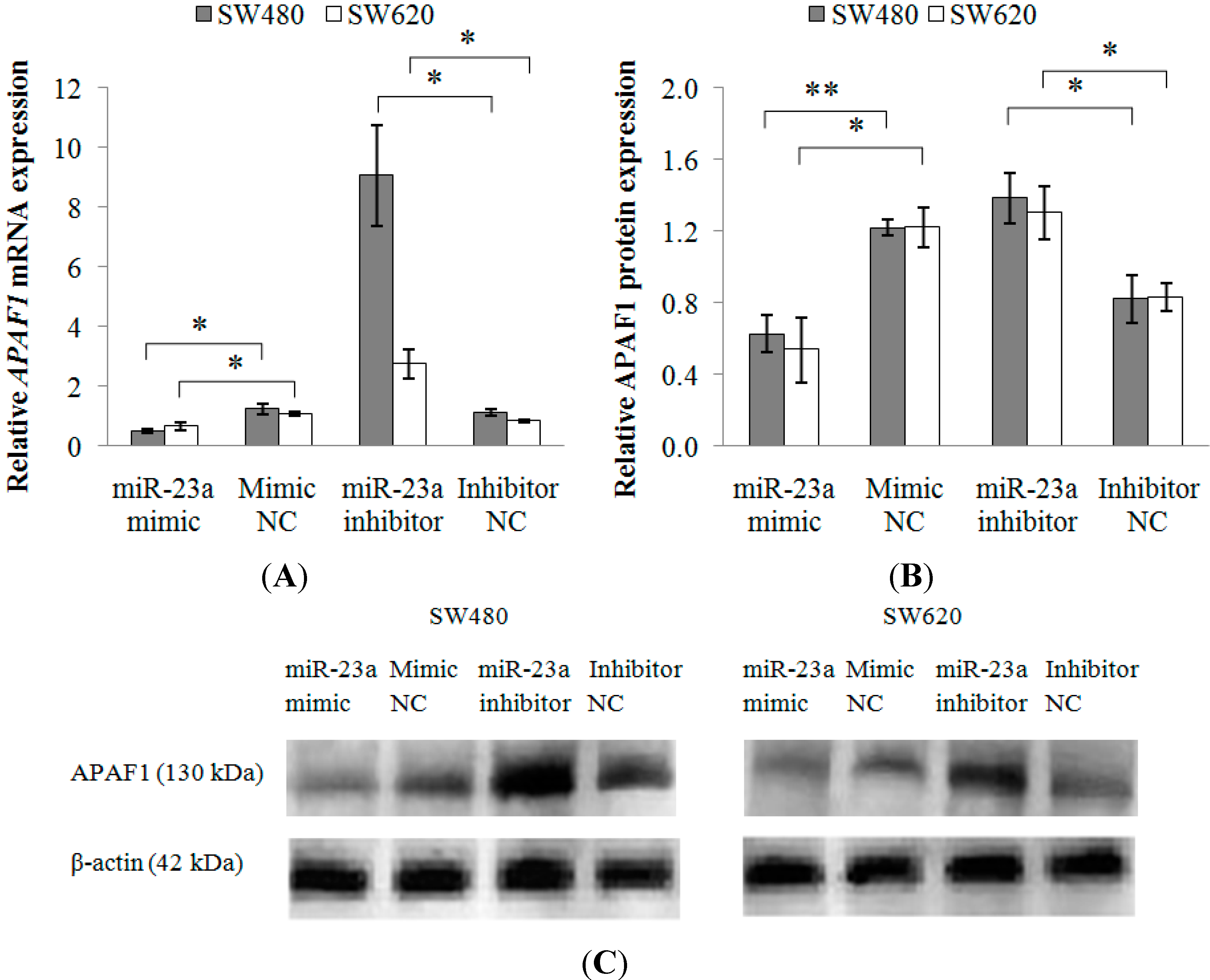
2.1.4. Modulation of APAF1 Expression in SW480 and SW620 Cell Lines
| Treatment | Relative miR-23a Expression | |
|---|---|---|
| SW480 | SW620 | |
| miR-23a mimic | 285.17 ± 50.11 | 234.59 ± 8.98 |
| miR-23a inhibitor | 0.03 ± 0.003 | 0.04 ± 0.005 |

2.1.5. APAF1 Down-Regulation in Clinical CRC Samples

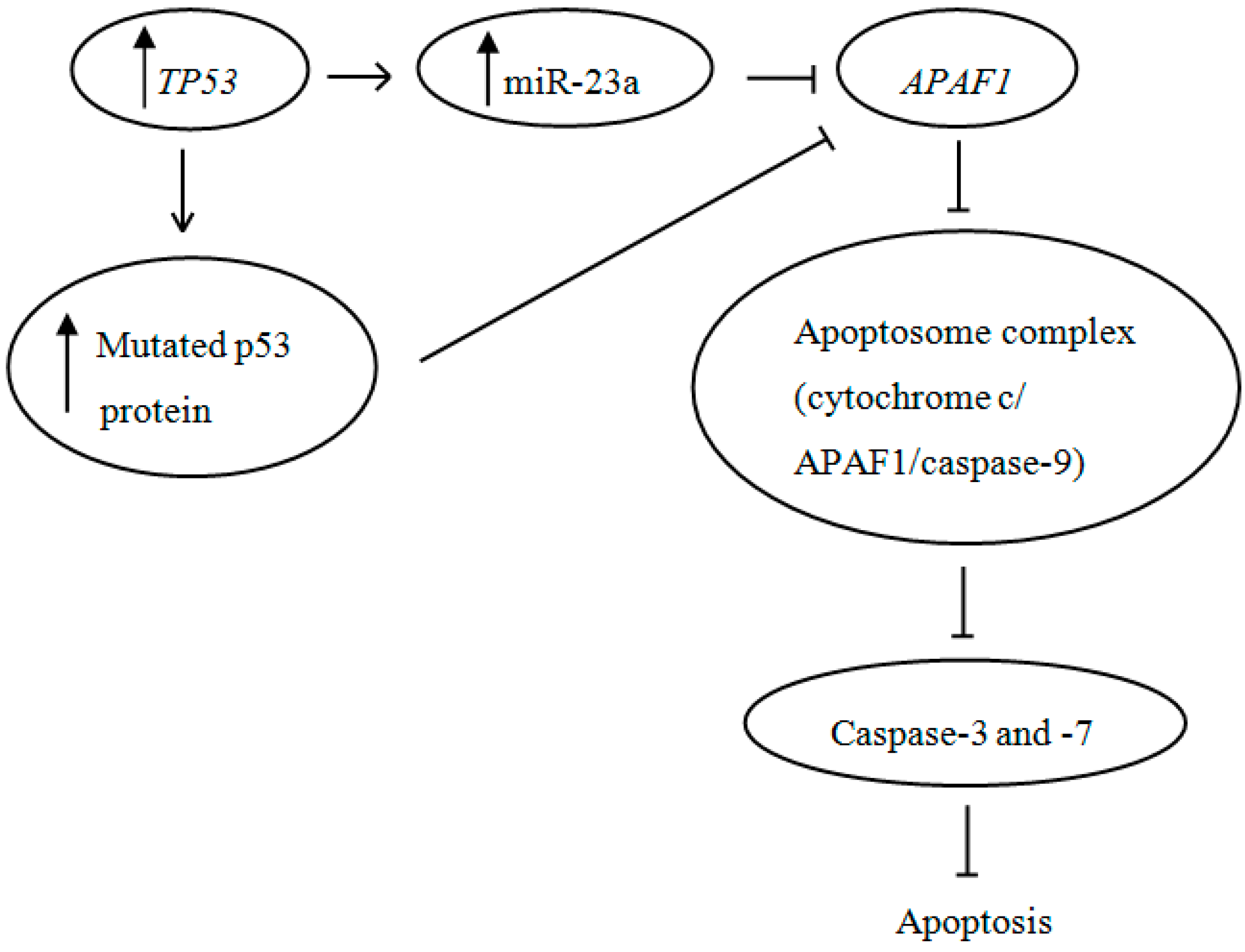
2.2. Discussion

3. Experimental Section
3.1. Clinical Sample Collection and Preservation
3.2. Cell Culture
3.3. miRNA Target Prediction
3.4. Total RNA Extraction
3.5. Reverse Transcription and Quantitative Real-Time PCR (RT-qPCR)
3.6. Transfection
3.7. Total Protein Extraction and Western Blot
3.8. Luciferase Reporter Construct
3.9. Luciferase Assay
3.10. Cell Viability Assay
3.11. Apoptosis Assay and Image-Based Cytometry
3.12. Caspase-Glo 3/7 Assay
3.13. Statistical Analysis
4. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bray, F.; Jemal, A.; Grey, N.; Ferlay, J.; Forman, D. Global cancer transitions according to the human development index (2008–2030): A population-based study. Lancet Oncol. 2012, 13, 790–801. [Google Scholar]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar]
- Chan, S.P.; Slack, F.J. MicroRNA-mediated silencing inside p-bodies. RNA Biol. 2006, 3, 97–100. [Google Scholar]
- Liu, X.; Fortin, K.; Mourelatos, Z. MicroRNAs: Biogenesis and molecular functions. Brain Pathol. 2008, 18, 113–121. [Google Scholar]
- Wu, L.; Fan, J.; Belasco, J.G. MicroRNAs direct rapid deadenylation of mRNA. Proc. Natl. Acad. Sci. USA 2006, 103, 4034–4039. [Google Scholar]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar]
- Lynam-Lennon, N.; Maher, S.G.; Reynolds, J.V. The roles of microRNA in cancer and apoptosis. Biol. Rev. Camb. Philos. Soc. 2009, 84, 55–71. [Google Scholar]
- Tsuchida, A.; Ohno, S.; Wu, W.; Borjigin, N.; Fujita, K.; Aoki, T.; Ueda, S.; Takanashi, M.; Kuroda, M. miR-92 is a key oncogenic component of the miR-17-92 cluster in colon cancer. Cancer Sci. 2011, 102, 2264–2271. [Google Scholar]
- Asangani, I.A.; Rasheed, S.A.; Nikolova, D.A.; Leupold, J.H.; Colburn, N.H.; Post, S.; Allgayer, H. MicroRNA-21 (miR-21) post-transcriptionally downregulates tumor suppressor Pdcd4 and stimulates invasion, intravasation and metastasis in colorectal cancer. Oncogene 2008, 27, 2128–2136. [Google Scholar]
- Yamamichi, N.; Shimomura, R.; Inada, K.; Sakurai, K.; Haraguchi, T.; Ozaki, Y.; Fujita, S.; Mizutani, T.; Furukawa, C.; Fujishiro, M.; et al. Locked nucleic acid in situ hybridization analysis of miR-21 expression during colorectal cancer development. Clin. Cancer Res. 2009, 15, 4009–4016. [Google Scholar]
- Chang, T.C.; Wentzel, E.A.; Kent, O.A.; Ramachandran, K.; Mullendore, M.; Lee, K.H.; Feldmann, G.; Yamakuchi, M.; Ferlito, M.; Lowenstein, C.J.; et al. Transactivation of miR-34a by p53 broadly influences gene expression and promotes apoptosis. Mol. Cell 2007, 26, 745–752. [Google Scholar]
- Yong, F.L.; Law, C.W.; Wang, C.W. Potentiality of a triple microRNA classifier: miR-193a-3p, miR-23a and miR-338-5p for early detection of colorectal cancer. BMC Cancer 2013, 13, 280. [Google Scholar]
- Krek, A.; Grün, D.; Poy, M.N.; Wolf, R.; Rosenberg, L.; Epstein, E.J.; MacMenamin, P.; da Piedade, I.; Gunsalus, K.C.; Stoffel, M.; et al. Combinatorial microRNA target predictions. Nat. Genet. 2005, 37, 495–500. [Google Scholar]
- Leibovitz, A.; Stinson, J.C.; McCombs, W.B.I.; McCoy, C.E.; Mazur, K.C.; Mabry, N.D. Classification of human colorectal adenocarcinoma cell lines. Cancer Res. 1976, 36, 4562–4569. [Google Scholar]
- Doench, J.G.; Sharp, P.A. Specificity of microRNA target selection in translational repression. Genes Dev. 2004, 18, 504–511. [Google Scholar]
- Huerta, S.; Heinzerling, J.H.; Anguiano-Hernandez, Y.M.; Huerta-Yepez, S.; Lin, J.; Chen, D.; Bonavida, B.; Livingston, E.H. Modification of gene products involved in resistance to apoptosis in metastatic colon cancer cells: Roles of Fas, Apaf-1, NFkappaB, IAPs, Smac/DIABLO, and AIF. J. Surg. Res. 2007, 142, 184–194. [Google Scholar]
- Hewitt, R.E.; McMarlin, A.; Kleiner, D.; Wersto, R.; Martin, P.; Tsokos, M.; Stamp, G.W.; Stetler-Stevenson, W.G. Validation of a model of colon cancer progression. J. Pathol. 2000, 192, 446–454. [Google Scholar]
- Li, B.; Sun, M.; Gao, F.; Liu, W.; Yang, Y.; Liu, H.; Cheng, Y.; Liu, C.; Cai, J. Up-regulated expression of miR-23a/b targeted the pro-apoptotic Fas in radiation-induced thymic lymphoma. Cell Physiol. Biochem. 2013, 32, 1729–1740. [Google Scholar]
- Guo, Z.; Zhou, B.; Liu, W.; Xu, Y.; Wu, D.; Yin, Z.; Permatasari, F.; Luo, D. miR-23a regulates DNA damage repair and apoptosis in UVB-irradiated HaCaT cells. J. Dermatol. Sci. 2013, 69, 68–76. [Google Scholar]
- Paik, S.S.; Jang, K.S.; Song, Y.S.; Jang, S.H.; Min, K.W.; Han, H.X.; Na, W.; Lee, K.H.; Choi, D.; Jang, S.J. Reduced expression of Apaf-1 in colorectal adenocarcinoma correlates with tumor progression and aggressive phenotype. Ann. Surg. Oncol. 2007, 14, 3453–3459. [Google Scholar]
- Zlobec, I.; Minoo, P.; Baker, K.; Haegert, D.; Khetani, K.; Tornillo, L.; Terracciano, L.; Jass, J.R.; Lugli, A. Loss of APAF-1 expression is associated with tumour progression and adverse prognosis in colorectal cancer. Eur. J. Cancer 2007, 43, 1101–1107. [Google Scholar]
- Luo, X.; Burwinkel, B.; Tao, S.; Brenner, H. MicroRNA signatures: Novel biomarker for colorectal cancer? Cancer Epidemiol. Biomark. Prev. 2011, 20, 1272–1286. [Google Scholar]
- Volinia, S.; Calin, G.A.; Liu, C.G.; Ambs, S.; Cimmino, A.; Petrocca, F.; Visone, R.; Lorio, M.; Roldo, C.; Ferracin, M.; et al. A microRNA expression signature of human solid tumors defines cancer gene targets. Proc. Natl. Acad. Sci. USA 2006, 103, 2257–2261. [Google Scholar]
- Meltzer, P.S. Cancer genomics: Small RNAs with big impacts. Nature 2005, 435, 745–746. [Google Scholar]
- Wang, N.; Zhu, M.; Tsao, S.W.; Man, K.; Zhang, Z.; Feng, Y. miR-23a-mediated inhibition of topoisomerase 1 expression potentiates cell response to etoposide in human hepatocellular carcinoma. Mol. Cancer 2013, 12, 119. [Google Scholar]
- Rodrigues, N.R.; Rowan, A.; Smith, M.E.; Kerr, I.B.; Bodmer, W.F.; Gannon, J.V.; Lane, D.P. p53 mutations in colorectal cancer. Proc. Natl. Acad. Sci. USA 1990, 87, 7555–7559. [Google Scholar]
- Violette, S.; Poulain, L.; Dussaulx, E.; Pepin, D.; Faussat, A.M.; Chambaz, J.; Lacorte, J.M.; Staedel, C.; Lesuffleur, T. Resistance of colon cancer cells to long-term 5-fluorouracil exposure is correlated to the relative level of bcl-2 and bcl-xl in addition to bax and p53 status. Int. J. Cancer 2002, 98, 498–504. [Google Scholar]
- Lacopetta, B. TP53 mutation in colorectal cancer. Hum. Mutat. 2003, 21, 271–276. [Google Scholar]
- Robles, A.I.; Bemmels, N.A.; Foraker, A.B.; Harris, C.C. APAF-1 is a transcriptional target of p53 in DNA damage-induced apoptosis. Cancer Res. 2001, 61, 6660–6664. [Google Scholar]
- Campioni, M.; Santini, D.; Tonini, G.; Murace, R.; Dragonetti, E.; Spugnini, E.P.; Baldi, A. Role of Apaf-1, a key regulator of apoptosis, in melanoma progression and chemoresistance. Exp. Dermatol. 2005, 14, 811–818. [Google Scholar]
- Cain, K.; Bratton, S.B.; Cohen, G.M. The apaf-1 apoptosome: A large caspase-activating complex. Biochimie 2002, 84, 203–214. [Google Scholar]
- Dweep, H.; Sticht, C.; Pandey, P.; Gretz, N. miRWalk-database: Prediction of possible miRNA binding sites by “walking” the genes of 3 genomes. J. Biomed. Inform. 2011, 44, 839–847. [Google Scholar]
- Yuan, J.S.; Reed, A.; Chen, F.; Stewart, C.N., Jr. Statistical analysis of real-time PCR data. BMC. Bioinform. 2006, 7, 85. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yong, F.L.; Wang, C.W.; Roslani, A.C.; Law, C.W. The Involvement of miR-23a/APAF1 Regulation Axis in Colorectal Cancer. Int. J. Mol. Sci. 2014, 15, 11713-11729. https://doi.org/10.3390/ijms150711713
Yong FL, Wang CW, Roslani AC, Law CW. The Involvement of miR-23a/APAF1 Regulation Axis in Colorectal Cancer. International Journal of Molecular Sciences. 2014; 15(7):11713-11729. https://doi.org/10.3390/ijms150711713
Chicago/Turabian StyleYong, Fung Lin, Chee Woon Wang, April Camilla Roslani, and Chee Wei Law. 2014. "The Involvement of miR-23a/APAF1 Regulation Axis in Colorectal Cancer" International Journal of Molecular Sciences 15, no. 7: 11713-11729. https://doi.org/10.3390/ijms150711713