Quantitative Trait Loci Mapping for Bacterial Blight Resistance in Rice Using Bulked Segregant Analysis

Abstract
:
1. Introduction
2. Results
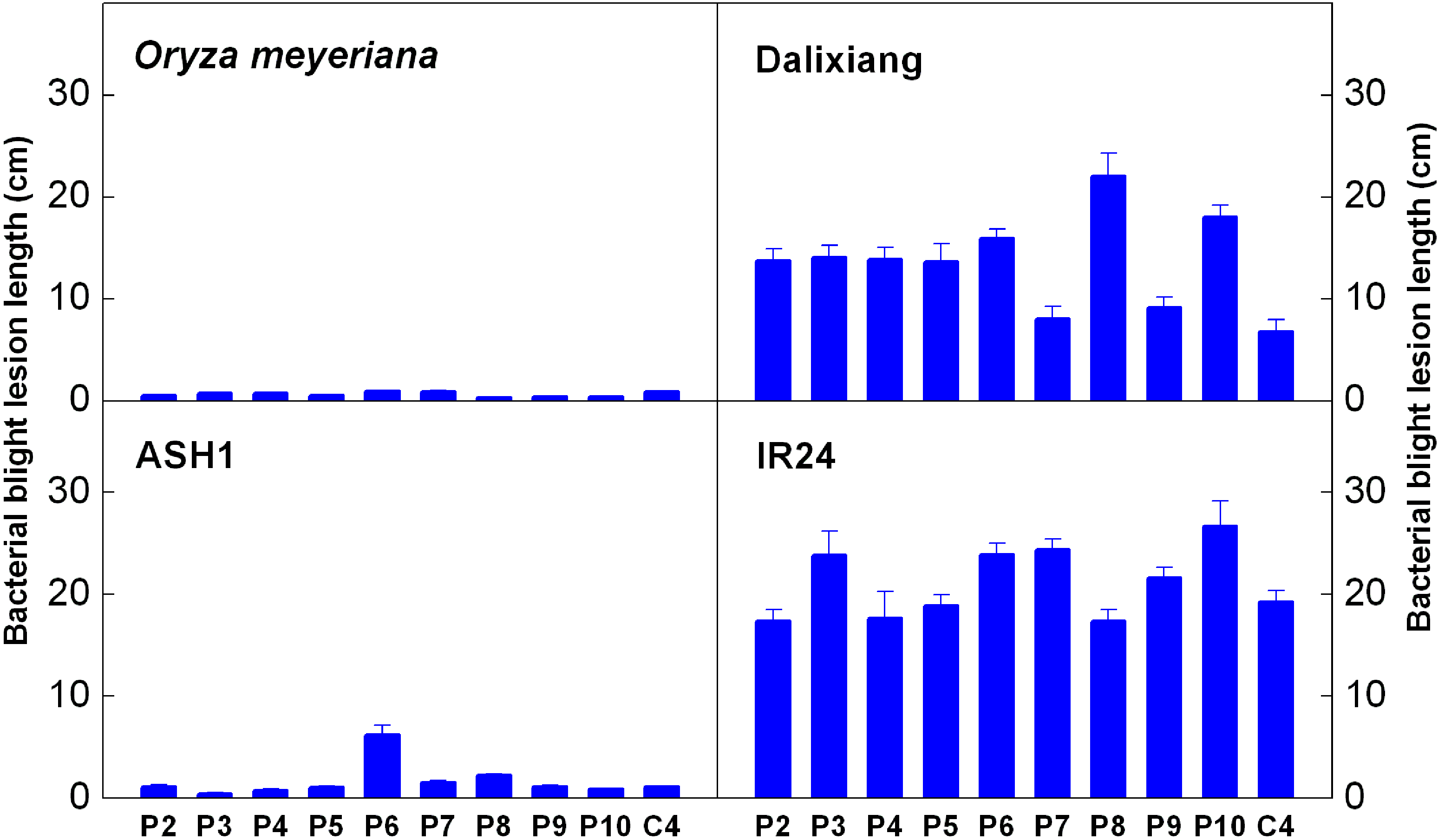
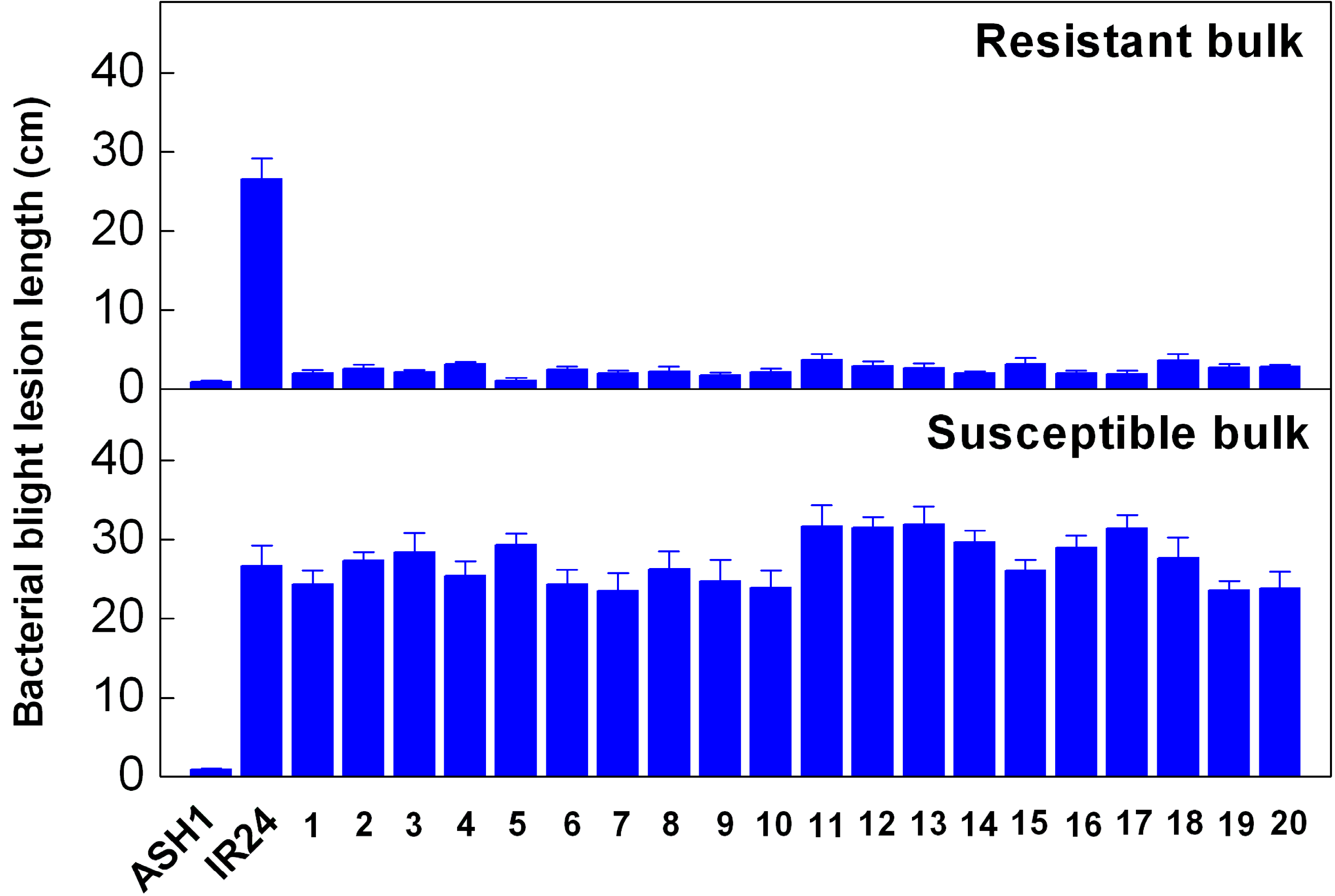
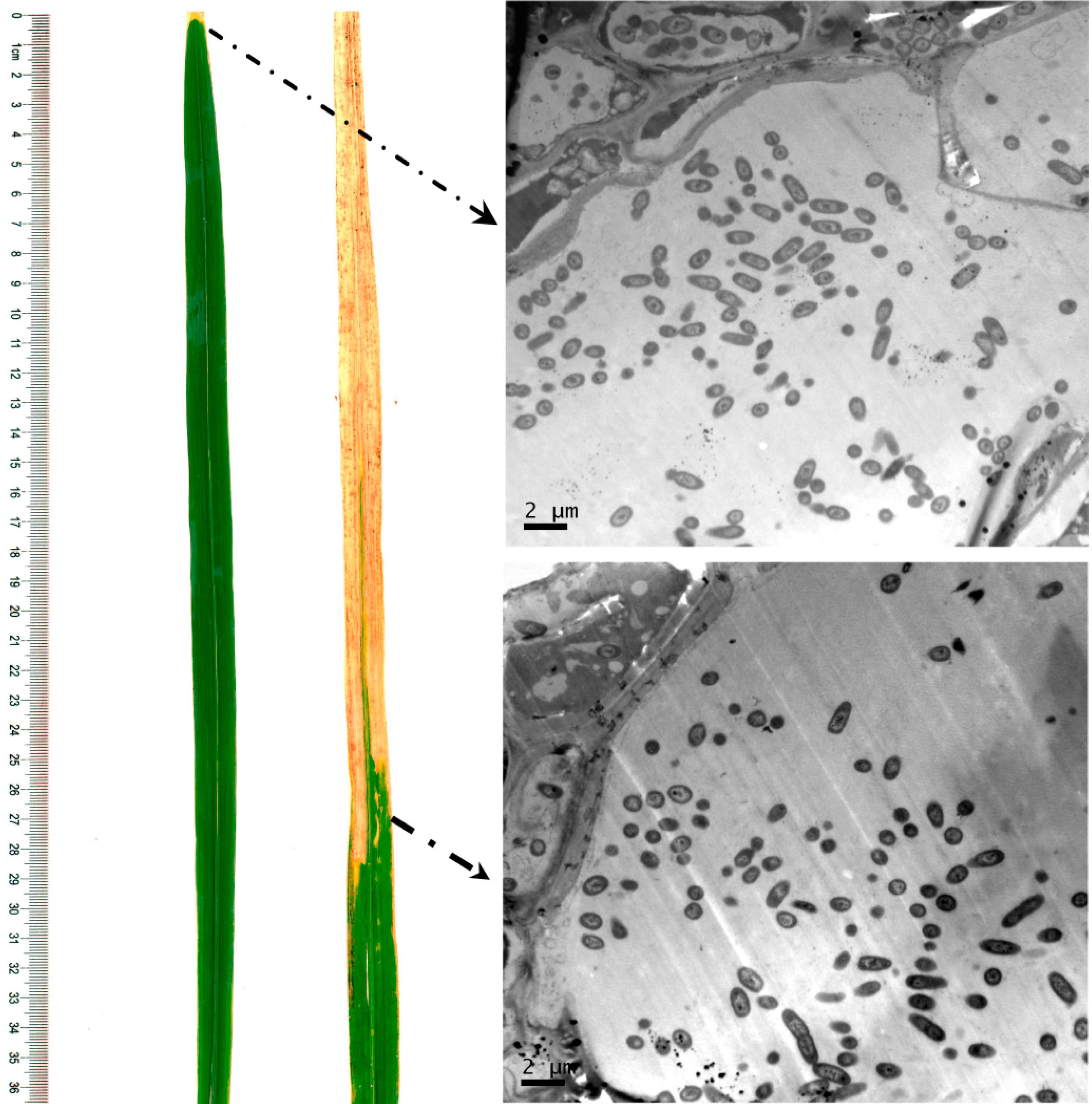
2.1. Resistance Analysis


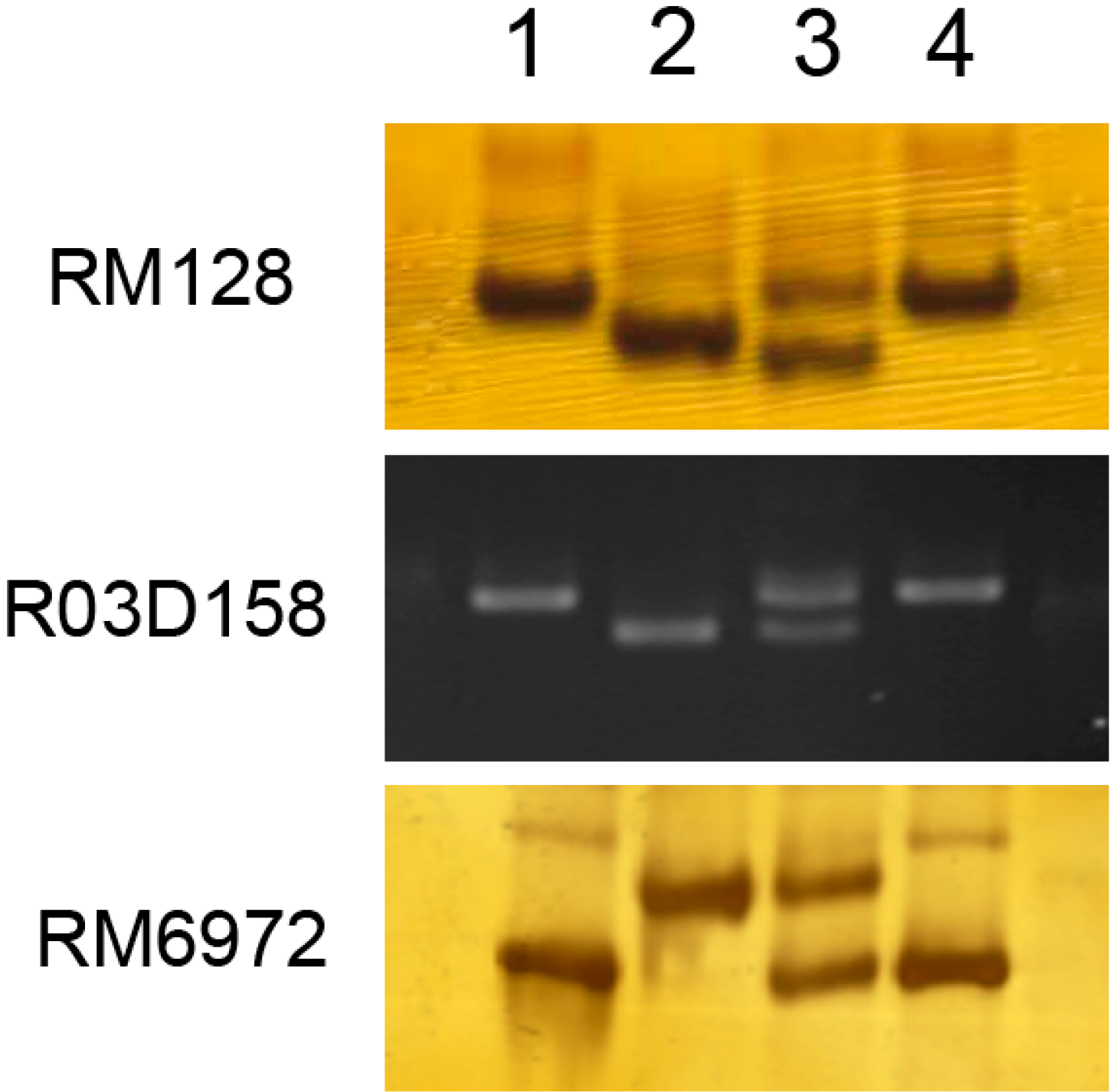
2.2. Markers Linked to Bacterial Blight (BB) Resistance



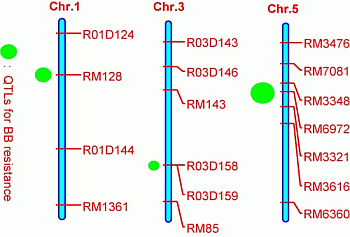
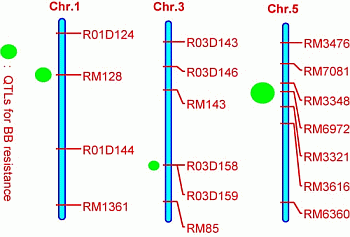
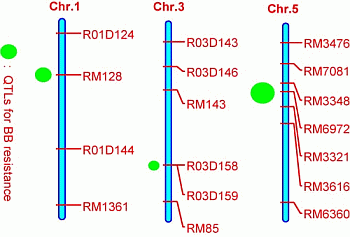
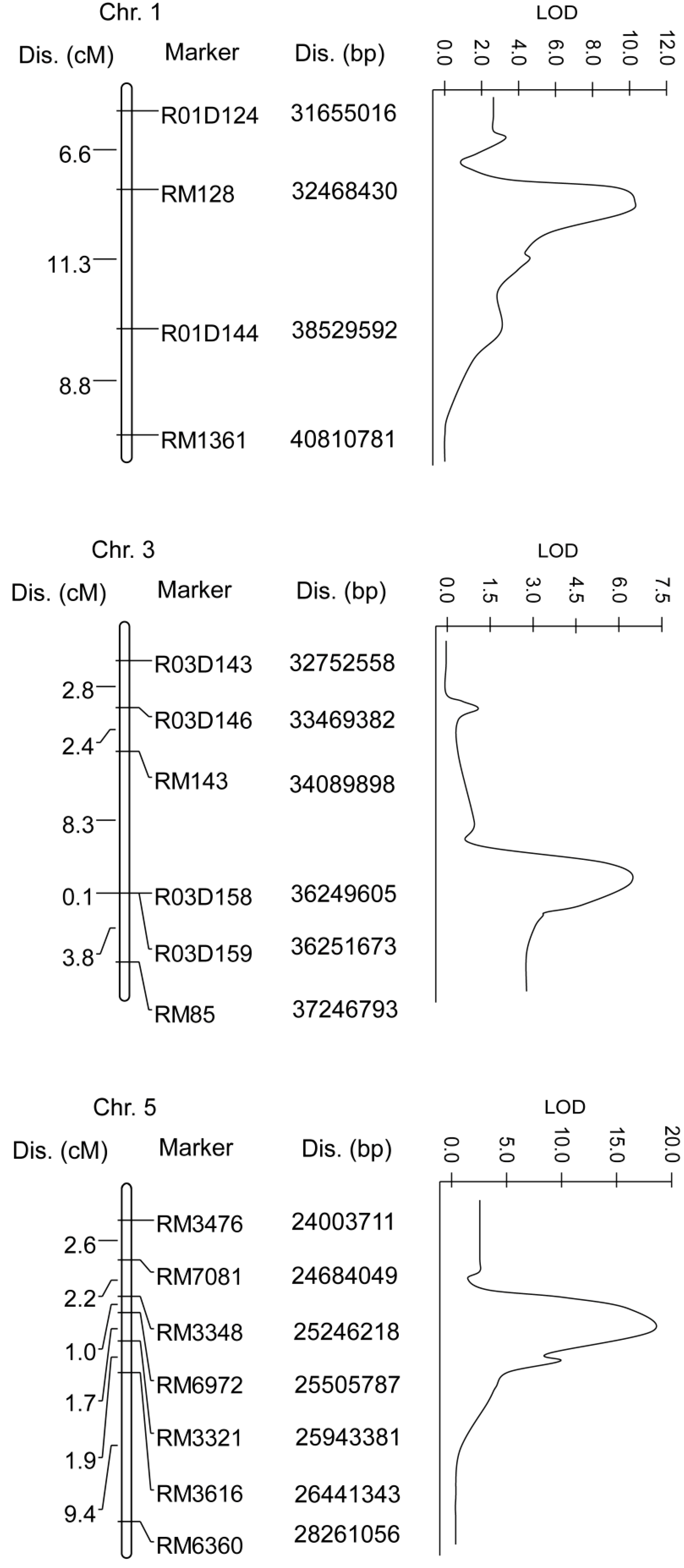
2.3. Quantitative Trait Loci (QTL) Mapping


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| QTL | Chromosome | Marker Interval | Nearest Marker | LOD | R2% | A |
|---|---|---|---|---|---|---|
| qBBR1 | 1 | RM128–R01D144 | RM128 | 10.1 | 21.5 | 4.89 |
| qBBR3 | 3 | R03D158–RM85 | R03D158 | 6.2 | 12.3 | 3.73 |
| qBBR5 | 5 | RM7081–RM3616 | RM6972 | 17.8 | 39.2 | 7.36 |
3. Discussion
4. Experimental Section
4.1. Plant Materials
4.2. Pathogen Inoculation and Resistance Identification
| Xoo Strain | Origin |
|---|---|
| P2 | Philippines |
| P3 | Philippines |
| P4 | Philippines |
| P5 | Philippines |
| P6 | Philippines |
| P7 | Philippines |
| P8 | Philippines |
| P9 | Philippines |
| P10 | Philippines |
| C4 | China |
4.3. Electron Microscopy
4.4. BSA Selection of Linkage Markers
4.5. Genetic Map Construction and QTL Mapping
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhang, Q. Genetics and Improvement of Resistance to Bacterial Blight in Rice, 1st ed.; Science Press: Beijing, China, 2007; pp. 1–4, 157–159. [Google Scholar]
- Guo, S.B.; Zhang, D.P.; Lin, X.H. Identification and mapping of a novel bacterial blight resistance gene Xa35(t) originated from Oryza minuta. Sci. Agric. Sin. 2010, 43, 2611–2618. [Google Scholar]
- Liu, Q.; Yuan, M.; Zhou, Y.; Li, X.; Xiao, J.; Wang, S. A paralog of the MtN3/saliva family recessively confers race-specific resistance to Xanthomonas oryzae in rice. Plant Cell Environ. 2011, 34, 1958–1969. [Google Scholar] [CrossRef]
- Tian, D.; Wang, J.; Zeng, X.; Gu, K.; Qiu, C.; Yang, X.; Zhou, Z.; Goh, M.; Luo, Y.; Murata-Hori, M.; et al. The rice TAL effector-dependent resistance protein XA10 triggers cell death and calcium depletion in the endoplasmic reticulum. Plant Cell 2014, 26, 497–515. [Google Scholar] [CrossRef]
- Song, W.Y.; Wang, G.L.; Chen, L.L.; Kim, H.S.; Pi, L.Y.; Holsten, T.; Gardner, J.; Wang, B.; Zhai, W.X.; Zhu, L.H.; et al. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science 1995, 270, 1804–1806. [Google Scholar]
- Yoshimura, S.; Yamanouchi, U.; Katayose, Y.; Toki, S.; Wang, Z.X.; Kono, I.; Kurata, N.; Yano, M.; Iwata, N.; Sasaki, T. Expression of Xa1, a bacterial blight-resistance gene in rice, is induced by bacterial inoculation. Proc. Natl. Acad. Sci. USA 1998, 95, 1663–1668. [Google Scholar] [CrossRef]
- Iyer, A.S.; McCouch, S.R. The rice bacterial blight resistance gene Xa5 encodes a novel form of disease resistance. Mol. Plant Microbe Interact. 2004, 17, 1348–1354. [Google Scholar] [CrossRef]
- Sun, X.; Cao, Y.; Yang, Z.; Xu, C.; Li, X.; Wang, S.; Zhang, Q. Xa26, a gene conferring resistance to Xanthomonas oryzae pv. oryzae in rice, encodes an LRR receptor kinase-like protein. Plant J. 2004, 37, 517–527. [Google Scholar] [CrossRef]
- Gu, K.; Yang, B.; Tian, D.; Wu, L.; Wang, D.; Sreekala, C.; Yang, F.; Chu, Z.; Wang, G.L.; White, F.F.; Yin, Z. R gene expression induced by a type-III effector triggers disease in rice. Nature 2005, 435, 1122–1125. [Google Scholar] [CrossRef]
- Chu, Z.; Yuan, M.; Yao, J.; Ge, X.; Yuan, B.; Xu, C.; Li, X.; Fu, B.; Li, Z.; Bennetzen, J.L.; et al. Promoter mutations of an essential gene for pollen development result in disease resistance in rice. Genes Dev. 2006, 20, 1250–1255. [Google Scholar] [CrossRef]
- Khush, G.S.; Mackill, D.J.; Sidhu, G.S. Breeding rice for resistance to bacterial blight. In Bacterial Blight of Rice, 1st ed.; International Rice Research Institute: Manila, Philippines, 1989; Volume 1; pp. 207–217. [Google Scholar]
- Huang, N.; Angeles, E.R.; Domingo, J.; Magpantay, G.; Singh, S.; Zhang, G.; Kumaravadivel, N.; Bennett, J.; Khush, G.S. Pyramiding of bacterial blight resistance genes in rice: Marker-assisted selection using RFLP and PCR. Theor. Appl. Genet. 1997, 95, 313–320. [Google Scholar] [CrossRef]
- Lee, S.W.; Choi, S.H.; Han, S.S.; Lee, D.G.; Lee, B.Y. Distribution of Xanthomonas oryzae pv. oryzae strains virulent to Xa21 in Korea. Phytopathology 1999, 89, 928–933. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, C.L.; Zhao, K.J.; Zhao, Y.L.; Caslana, V.C.; Zhu, X.D.; Li, D.Y.; Jiang, Q.X. The effectiveness of advanced rice lines with new resistance gene Xa23 to rice bacterial blight. Rice Genet. Newsl. 2001, 18, 71–73. [Google Scholar]
- Peng, S.Q.; Wei, Z.S.; Mao, C.X.; Huang, H.Q.; Xiao, F.H.; Luo, K. Identification of multiresistance of O. meyeriana, O. officinalis and O. sativa f. spantanea growing in Yunan province. Acta Phytopathol. Sin. 1982, 12, 58–60. [Google Scholar]
- Zhang, Q.; Wang, C.; Shi, A.; Bai, J.; Ling, S.; Li, D.; Chen, C.; Pang, H. Evaluation of resistance to bacterial blight (Xanthomonas oryzae pv. oryzae) in wild rice species. Sci. Agric. Sin. 1994, 27, 1–9. [Google Scholar]
- Yan, C.Q.; Qian, K.X.; Yan, Q.S.; Zhang, X.Q.; Xue, G.P.; Huangfu, W.G.; Wu, Y.F.; Zhao, Y.Z.; Xue, Z.Y.; Huang, J.; et al. Use of asymmetric somatic hybridization for transfer of the bacterial resistance trait from Oryza meyeriana L. to O. sativa L. ssp. japonica. Plant Cell Rep. 2004, 22, 569–575. [Google Scholar] [CrossRef]
- Cheng, Z.Q.; Yan, H.J.; Geng, X.S.; Yin, F.Y.; Sun, Y.D.; Zhang, C.; Huang, X.Q. Identification of Oryza granulata for resistance to Xanthomonas oryzae pv. oryzae and observation of leaf tissue. Acta Phytopathol. Sin. 2008, 38, 582–591. [Google Scholar]
- Aggarwarl, R.K.; Brar, D.S.; Khush, G.S. Two new genomes in the Oryza complex identified on the basis of molecular divergence analysis using total genomic DNA hybridization. Mol. Gen. Genet. 1997, 254, 1–12. [Google Scholar] [CrossRef]
- Kinoshita, T.; Mori, K. In vitro techniques for genomic alteration in rice plants. Euphytica 2001, 120, 367–372. [Google Scholar] [CrossRef]
- Michelmore, R.W.; Paran, I.; Kesseli, R.V. Identification of markers linked to disease-resistance genes by bulked segregant analysis: A rapid method to detect markers in specific genomic regions by using segregating populations. Proc. Natl. Acad. Sci. USA 1991, 88, 9828–9832. [Google Scholar] [CrossRef]
- Zhang, Q.F.; Shen, B.Z.; Dai, X.K.; Mei, M.H.; Saghai Maroof, M.A.; Li, Z.B. Using bulked extremes and recessive class to map genes for photoperiod-sensitive genic male sterility in rice. Proc. Natl. Acad. Sci. USA 1994, 91, 8675–8679. [Google Scholar] [CrossRef]
- Wang, G.L.; Ruan, D.L.; Song, W.Y.; Sideris, S.; Chen, L.L.; Pi, L.Y.; Zhang, S.; Zhang, Z.; Fauquet, C.; Gaut, B.S.; et al. Xa21D encodes a receptor-like molecule with a leucine-rich repeat domain that determines race-specific recognition and is subject to adaptive evolution. Plant Cell 1998, 10, 765–779. [Google Scholar] [CrossRef]
- Gu, K.; Tian, D.; Yang, F.; Wu, L.; Sreekala, C.; Wang, D.; Wang, G.L.; Yin, Z. High-resolution genetic mapping of Xa27(t), a new bacterial blight resistance gene in rice, Oryza sativa L. Theor. Appl. Genet. 2004, 108, 800–807. [Google Scholar] [CrossRef]
- Wang, C.L.; Chen, L.T.; Zeng, C.Z.; Zhang, Q.Y.; Liu, P.Q.; Liu, Y.G.; Fan, Y.L.; Zhang, Q.; Zhao, K.J. Chromosome walking for fine mapping of Xa23 gene locus by using genomic libraries. Chin. J. Rice Sci. 2006, 20, 355–360. [Google Scholar]
- Tan, G.X.; Ren, X.; Weng, Q.M.; Shi, Z.Y.; Zhu, L.L.; He, G.C. Mapping of a new resistance gene to bacterial blight in rice line introgressed from Oryza officinalis. Acta Genet. Sin. 2004, 31, 724–729. [Google Scholar]
- Liu, J.M.; Cheng, Z.Q.; Yang, M.Z.; Wu, C.J.; Wang, L.X.; Sun, Y.D.; Huang, X.Q. Cloning and sequence analysis of disease resistance gene analogues from three wild rice species in Yunnan. Sci. Agric. Sin. 2003, 36, 273–280. [Google Scholar]
- Yang, Z.; Sun, X.; Wang, S.; Zhang, Q. Genetic and physical mapping of a new gene for bacterial blight resistance in rice. Theor. Appl. Genet. 2003, 106, 1467–1472. [Google Scholar]
- Xiang, Y.; Cao, Y.L.; Xu, C.G.; Li, X.H.; Wang, S.P. Xa3, conferring resistance for rice bacterial blight and encoding a receptor kinase-like protein, is the same as Xa26. Theor. Appl. Genet. 2006, 113, 1347–1355. [Google Scholar] [CrossRef]
- Blair, M.W.; Garris, A.J.; Iyer, A.S.; Chapman, B.; Kresovich, S.; McCouch, S.R. High resolution genetic mapping and candidate gene identification at the Xa5 locus for bacterial blight resistance in rice (Oryza sativa L.). Theor. Appl. Genet. 2003, 107, 62–73. [Google Scholar]
- Zhang, G.; Angeles, E.R.; Abenes, M.L.P.; Khush, G.S.; Huang, N. RAPD and RFLP mapping of the bacterial blight resistance gene Xa-13 in rice. Theor. Appl. Genet. 1996, 93, 65–70. [Google Scholar] [CrossRef]
- Ronald, P.C.; Albano, B.; Tabien, R.; Abenes, L.; Wu, K.S.; McCouch, S.; Tanksley, S.D. Genetic and physical analysis of the rice bacterial blight disease resistance locus, Xa21. Mol. Gen. Genet. 1992, 236, 113–120. [Google Scholar]
- Chen, H.; Wang, S.; Zhang, Q. New gene for bacterial blight resistance in rice located on chromosome 12 identified from Minghui 63, an elite restorer line. Phytopathology 2002, 92, 750–754. [Google Scholar] [CrossRef]
- Yang, Y.; Yu, C.L.; Wang, X.M.; Yan, C.Q.; Cheng, Y.; Chen, J.P. Inoculation with Xanthomonas oryzae pv. oryzae induces thylakoid membrane association of Rubisco activase in Oryza meyeriana. J. Plant Physiol. 2011, 168, 1701–1704. [Google Scholar] [CrossRef]
- Yang, Y.; Xie, L.; Yan, C.Q.; Wang, X.M.; Yu, C.L.; Cheng, X.Y.; Cheng, Y.; Chen, J.P. Xylem secondary cell-wall thickening involved in defense responses of Oryza meyeriana to Xanthomonas oryzae pv. oryzae. Acta Phytopathol. Sin. 2012, 42, 505–514. [Google Scholar]
- Kauffman, H.E.; Reddy, A.P.K.; Hsieh, S.P.Y.; Merca, S.D. An improved technique for evaluating resistance of rice varieties to Xanthomonas oryzae. Plant Dis. Rep. 1973, 57, 537–541. [Google Scholar]
- Sanchez, A.C.; Ilag, L.L.; Yang, D.; Brar, D.S.; Ausubel, F.; Khush, G.S.; Yano, M.; Sasaki, T.; Li, Z.; Huang, N. Genetic and physical mapping of Xa13, a recessive bacterial blight resistance gene in rice. Theor. Appl. Genet. 1999, 98, 1022–1028. [Google Scholar] [CrossRef]
- Hong, J.; Jiang, D.A.; Weng, X.Y.; Wang, W.B.; Hu, D.W. Leaf anatomy, chloroplast ultrastructure, and cellular localisation of ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBPCO) and RuBPCO activase in Amaranthus tricolor L. Photosynthetica 2005, 43, 519–528. [Google Scholar] [CrossRef]
- Wu, S.B.; Wirthensohn, M.G.; Hunt, P.; Gibson, J.P.; Sedgley, M. High resolution melting analysis of almond SNPs derived from ESTs. Theor. Appl. Genet. 2008, 118, 1–14. [Google Scholar] [CrossRef]
- Lander, E.S.; Green, P.; Abrahamson, J.; Barlow, A.; Daly, M.J.; Lincoln, S.E.; Newberg, L.A. MAPMAKER: An interactive computer package for constructing primary genetic linkage maps of experimental and natural populations. Genomics 1987, 1, 174–181. [Google Scholar] [CrossRef]
- Wang, S.; Basten, C.J.; Zeng, Z.B. Windows QTL Cartographer 2.5; Department of Statistics, North Carolina State University: Raleigh, NC, USA, 2007. [Google Scholar]
- Churchill, G.A.; Doerge, R.W. Empirical threshold values for quantitative trait mapping. Genetics 1994, 138, 963–971. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Han, X.; Yang, Y.; Wang, X.; Zhou, J.; Zhang, W.; Yu, C.; Cheng, C.; Cheng, Y.; Yan, C.; Chen, J. Quantitative Trait Loci Mapping for Bacterial Blight Resistance in Rice Using Bulked Segregant Analysis. Int. J. Mol. Sci. 2014, 15, 11847-11861. https://doi.org/10.3390/ijms150711847
Han X, Yang Y, Wang X, Zhou J, Zhang W, Yu C, Cheng C, Cheng Y, Yan C, Chen J. Quantitative Trait Loci Mapping for Bacterial Blight Resistance in Rice Using Bulked Segregant Analysis. International Journal of Molecular Sciences. 2014; 15(7):11847-11861. https://doi.org/10.3390/ijms150711847
Chicago/Turabian StyleHan, Xueying, Yong Yang, Xuming Wang, Jie Zhou, Wenhao Zhang, Chulang Yu, Chen Cheng, Ye Cheng, Chengqi Yan, and Jianping Chen. 2014. "Quantitative Trait Loci Mapping for Bacterial Blight Resistance in Rice Using Bulked Segregant Analysis" International Journal of Molecular Sciences 15, no. 7: 11847-11861. https://doi.org/10.3390/ijms150711847