
Simultaneous Analysis of SEPT9 Promoter Methylation Status, Micronuclei Frequency, and Folate-Related Gene Polymorphisms: The Potential for a Novel Blood-Based Colorectal Cancer Biomarker


Abstract
:
1. Introduction
2. Results and Discussion




{kind=link}
{kind=link}
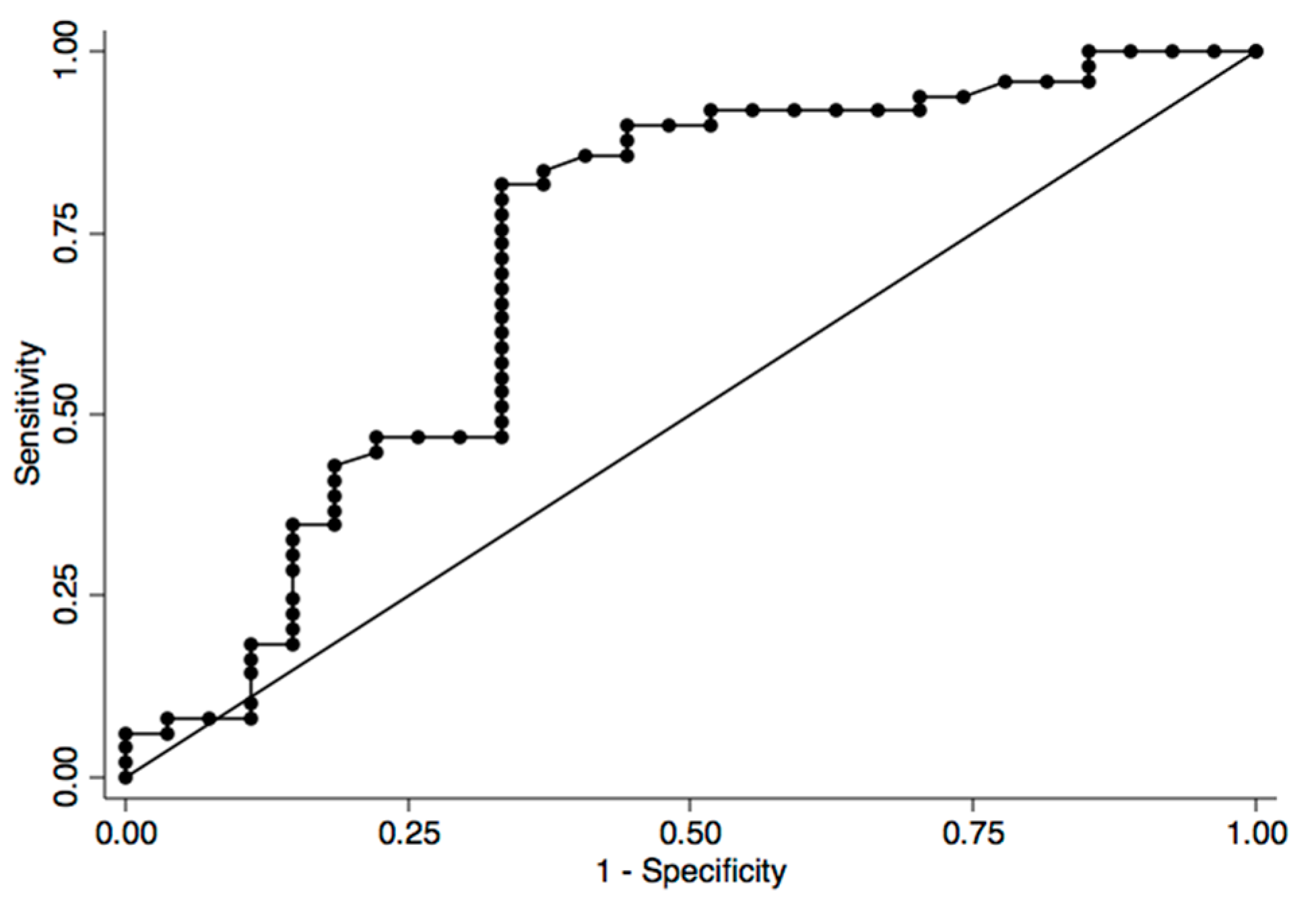{kind=link}
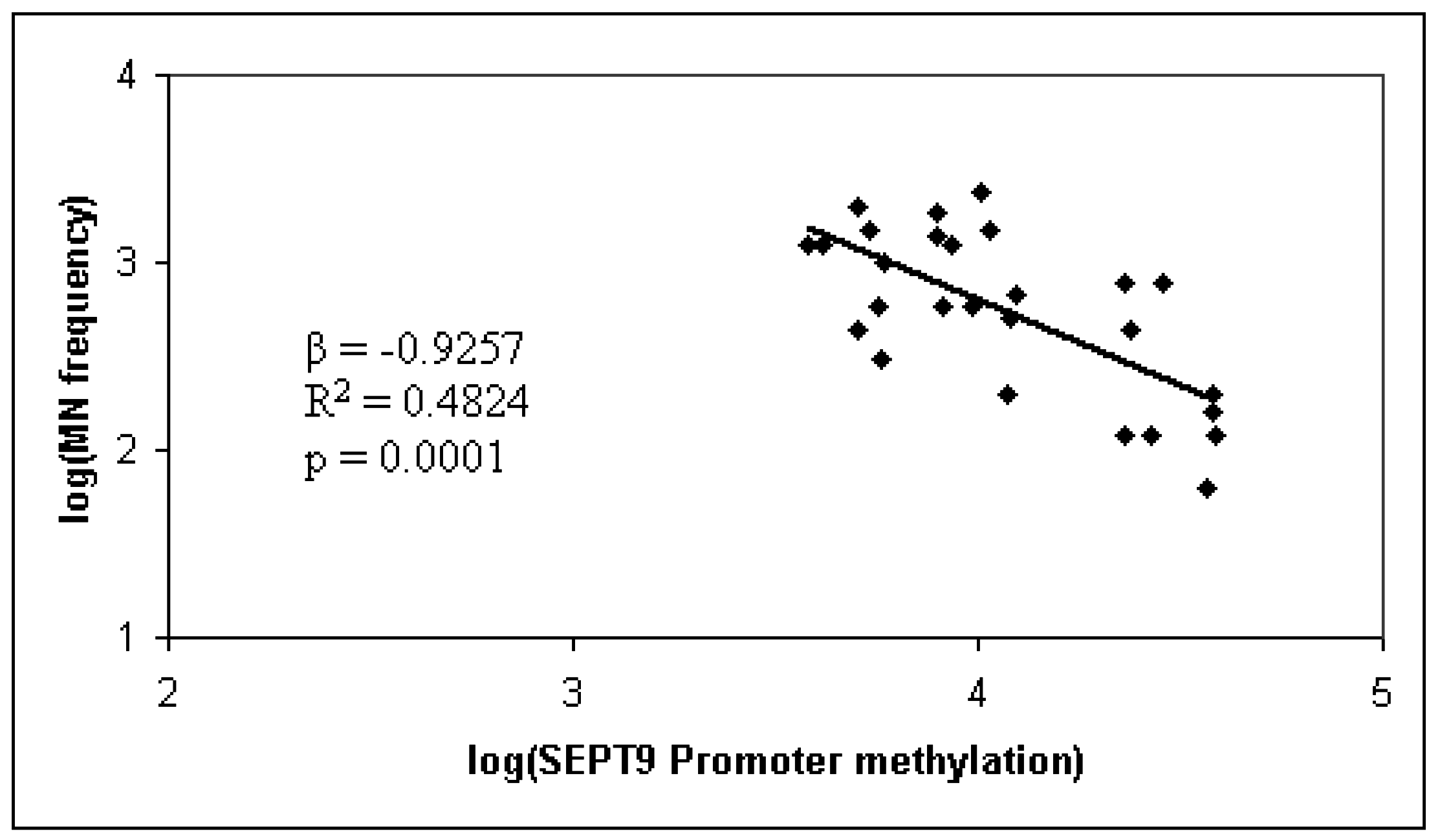{kind=link}
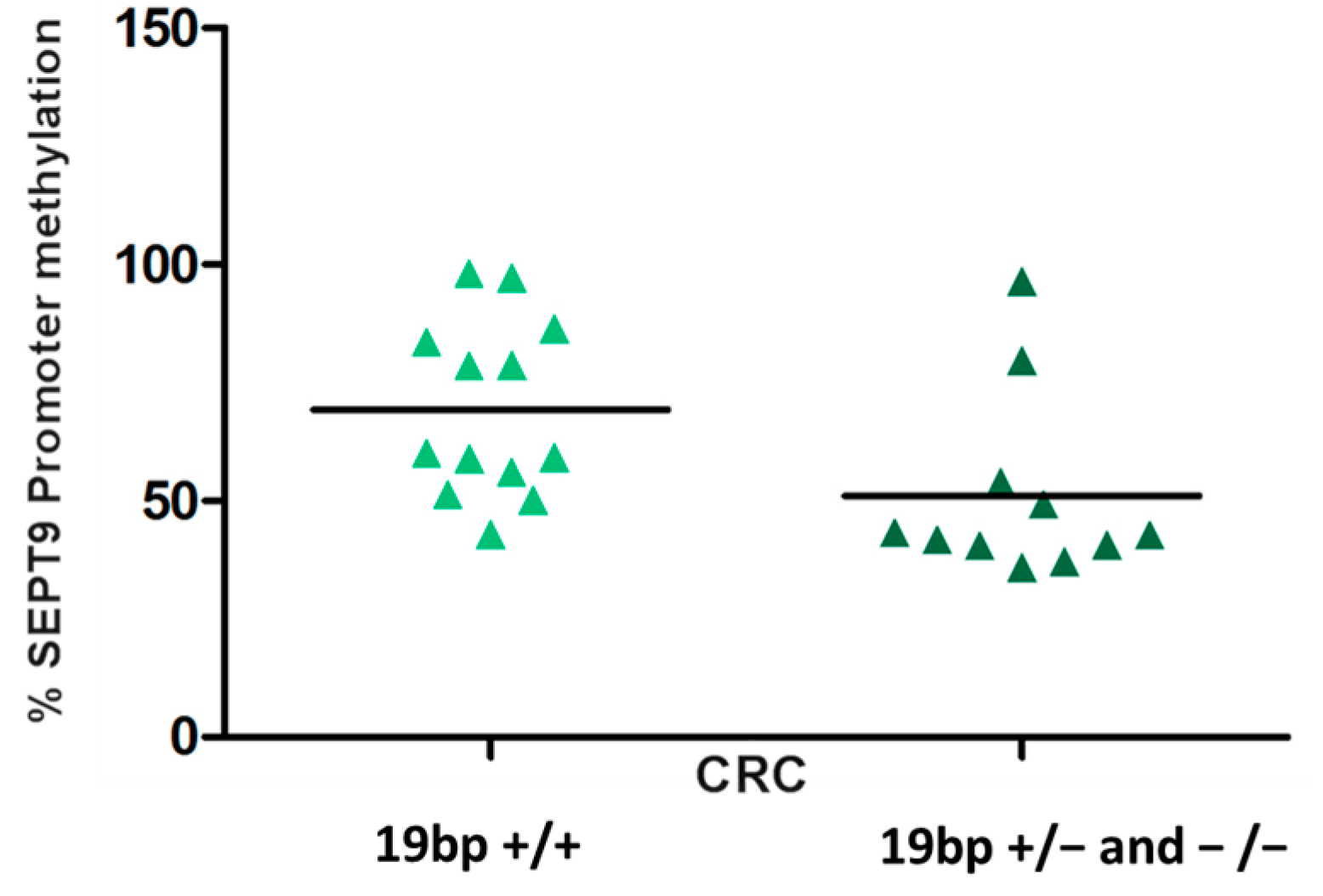{kind=link}
| Genotype | CRC n (%) | Controls n (%) | OR (95% CI) | p |
|---|---|---|---|---|
| RFC rs1051266, p.His27Arg | ||||
| AA | 8 (33.3) | 9 (34.6) | 1.00 | 1.000 |
| AG | 12 (50.0) | 13 (50.0) | 1.04 (0.26–4.25) | |
| GG | 4 (16.7) | 4 (15.4) | 1.11 (0.15–8.32) | |
| AG/GG | 16 (66.7) | 17 (65.4) | 1.06 (0.28–4.03) | |
| FOLR rs20710110, −20G>A | ||||
| AA | 23 (95.8) | 22 (84.6) | 1.00 | 0.351 |
| AG | 1 (4.2) | 4 (15.4) | 0.25 (0.01–2.74) | |
| DHFR rs70991108, 86+59_86+60insACCTGGGCGGGACGCGCCA | ||||
| 19+/+ | 13 (54.2) | 7 (26.9) | 1.00 | 0.144 |
| 19+/− | 9 (37.5) | 14 (53.9) | 0.36 (0.08–1.41) | |
| 19-/− | 2 (8.3) | 5 (19.2) | 0.23 (0.02–1.85) | |
| 19+/−_19−/− | 11 (45.8) | 19 (73.1) | 0.32 (0.08–1.17) | |
| TS 28BP rs45445694. *34+169_*34+196CGGGACGGAGGCAGGCCAAGTGGCGCGG [2,3] | ||||
| 2R2R | 6 (25.0) | 4 (15.4) | 1.00 | 0.013 |
| 2R3R | 16 (66.7) | 10 (38.5) | 1.06 (0.17–5.90) | |
| 3R3R | 2 (8.3) | 12 (46.1) | 0.12 (0.01–1.05) | |
| 2R3R/3R3R | 18 (75.0) | 22 (84.6) | 0.55 (0.10–2.75) | |
| SHMT rs1979277, p.Leu474Phe | ||||
| CC | 16 (66.7) | 14 (53.9) | 1.00 | 0.686 |
| CT | 7 (29.2) | 11 (42.3) | 0.56 (0.14–2.12) | |
| TT | 1 (4.1) | 1 (3.9) | 0.88 (0.01–73.53) | |
| CT/TT | 8 (33.3) | 12 (46.2) | 0.59 (0.16–2.11) | |
| MTHFR rs1801133, p.Ala222Val | ||||
| CC | 7 (29.2) | 6 (23.1) | 1.00 | 0.867 |
| CT | 13 (54.2) | 14 (53.8) | 0.80 (0.17–3.65) | |
| TT | 4 (16.6) | 6 (23.1) | 0.59 (0.08–4.00) | |
| CT/TT | 17 (79.8) | 20 (76.9) | 0.73 (0.17–3.11) | |
| MTHFR r1801131, p.Glu429Ala | ||||
| AA | 15 (62.5) | 11 (42.3) | 1.00 | 0.424 |
| AC | 8 (33.3) | 13 (50.0) | 0.46 (0.12–1.69) | |
| CC | 1 (4.2) | 2 (7.7) | 0.38 (0.01–8.16) | |
| AC/CC | 9 (37.5) | 15 (57.7) | 0.45 (0.12–1.57) | |
| MTRR rs1801394, p.Ile49Met | ||||
| AA | 13 (54.2) | 6 (23.1) | 1.00 | 0.032 |
| AG | 9 (37.5) | 12 (46.1) | 0.36 (0.08–1.50) | |
| GG | 2 (8.3) | 8 (30.8) | 0.13 (0.01–0.89) | |
| AG/GG | 11 (45.8) | 20 (76.9) | 0.26 (0.06–0.99) | |

3. Methods
3.1. Study Population
| Characteristics | Controls | Colorectal Polyps | CRC | |
|---|---|---|---|---|
| Subjects | 26 | 23 | 27 | |
| Age | Mean ± SD | 59.62 ± 5.82 | 60.43 ± 6.14 | 67.52 ± 9.61 |
| Range | 51−70 | 48−70 | 53−88 | |
| Gender n (%) | Female | 7 (26.9%) | 10 (43.5%) | 10 (38.5%) |
| Male | 19 (73.1%) | 13 (56.5%) | 16 (61.5%) | |
| BMI | Mean ± SD | 25.65 ± 3.31 | 26.88 ± 3.09 | 24.51 ± 3.48 |
| Range | 18.37−32.42 | 22.03−35.16 | 19.61−33.73 | |
| Smoke n (%) | No | 19 (73.1%) | 18 (78.3%) | 18 (69.2%) |
| Yes | 7 (26.9%) | 5 (21.7%) | 8 (30.8%) |
3.2. Methylation Analysis
3.3. Micronuclei Analysis
3.4. Cell Culture
3.5. Genotyping Analysis
3.6. Statistical Analysis
Supplementary Materials
Acknowledgments
Authors Contributions
Conflicts of Interest
References
- Colussi, D.; Brandi, G.; Bazzoli, F.; Ricciardiello, L. Molecular pathways involved in colorectal cancer: Implication for disease behavior and prevention. Int. J. Mol. Sci. 2013, 14, 16365–16385. [Google Scholar] [CrossRef] [PubMed]
- Ait Ouakrim, D.; Lockett, T.; Boussiooutas, A.; Keogh, L.; Flander, L.B.; Hopper, J.L.; Jenkins, M.A. Screening participation predictors for people at familial risk of colorectal cancer: A systematic review. Am. J. Prev. Med. 2013, 44, 496–506. [Google Scholar] [CrossRef] [PubMed]
- Mobley, L.; Kuo, T.M.; Urato, M.; Boos, J.; Lozano-Garcia, N.; Anselin, L. Predictors of endoscopic colorectal cancer screening over time in 11 states. Cancer Causes Control. 2010, 21, 445–461. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; Brenner, D.E.; Turgeon, D.K. Early detection of colon cancer: New tests on the horizon. Mol. Diagn. Ther. 2008, 12, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Maffei, F.; Angeloni, C.; Malaguti, M.; Moraga, J.M.; Pasqui, F.; Poli, C.; Colecchia, A.; Festi, D.; Hrelia, P.; Hrelia, S. Plasma antioxidant enzymes and clastogenic factors as possible biomarkers of colorectal cancer risk. Mutat. Res. 2011, 714, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.W.; Liu, H.; Shin, D.H.; Yu, G.I.; Hwang, J.S.; Km, E.S.; Yun, J.W. Proteomic and cytokine plasma biomarkers for predicting progression from colorectal adenoma to carcinoma in human patients. Proteomics 2013, 13, 2361–2374. [Google Scholar] [CrossRef] [PubMed]
- Bonassi, S.; El-Zein, R.; Bolognesi, C.; Fenech, M. Micronuclei frequency in peripheral blood lymphocytes and cancer risk: Evidence from human studies. Mutagenesis 2011, 26, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Barrera, V.; Peinado, M.A. Evaluation of single CpG site as proxies of CpG island methylation status at the genome scale. Nucleic Acid Res. 2012, 40, 1490–1498. [Google Scholar] [CrossRef] [PubMed]
- Ferraro, A.; Kontos, C.K.; Boni, T.; Bantounas, I.; Siakouli, D.; Kosmidou, V.; Vlassi, M.; Spyridakis, Y.; Tsipras, I.; Zografos, G.; et al. Epigenetic regulation of miR-21 in colorectal cancer: ITGB4 as a novel miR-21 target and a three-gene network (miR21-ITGB4-PCDC4) as a predictor of tumor potential. Epigenetics 2013, 9, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Carmona, F.J.; Esteller, M. Moving closer to a prognostic DNA methylation signature in colon cancer. Clin. Cancer Res. 2011, 17, 1215–1217. [Google Scholar] [CrossRef] [PubMed]
- Lange, C.P.; Campan, M.; Hinoue, T.; Schmitz, R.F.; van der Meulen-de Jong, A.E.; Slingerland, H.; Kok, P.J.; van Dijk, C.M.; Weisenberger, D.J.; Shen, H.; et al. Genome-scale discovery of DNA-methylation biomarkers for blood-based detection of colorectal cancer. PLoS ONE 2012, 7, e50266. [Google Scholar] [CrossRef] [PubMed]
- Summers, T.; Langan, R.C.; Nissan, A.; Brücher, B.L.; Bilchik, A.J.; Protic, M.; Daumer, M.; Avital, I.; Stojadinovic, A. Serum-based DNA methylation biomarkers in colorectal cancer: Potential for screening and early detection. J. Cancer 2013, 4, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Tänzer, M.; Balluff, B.; Distler, J.; Hale, K.; Leodolter, A.; Röcken, C.; Molnar, B.; Schmid, R.; Lofton-Day, C.; Schuster, T.; et al. Performance of epigenetic markers SEPT9 and ALX4 in plasma for detection of colorectal precancerous lesions. PLoS ONE 2010, 5, e9061. [Google Scholar] [CrossRef] [PubMed]
- Warren, J.D.; Xiong, W.; Bunker, A.M.; Vaughn, C.P.; Furtado, L.V.; Roberts, W.L.; Fang, J.C.; Samowitz, W.S.; Heichman, K.A. Septin 9 methylated DNA is a sensitive and specific blood test for colorectal cancer. BMC Med. 2011, 9, 133. [Google Scholar] [CrossRef] [PubMed]
- Connoly, D.; Abdesselam, I.; Verdier-Pinard, P.; Montagna, C. Septin roles in tumorigenesis. Biol. Chem. 2011, 392, 725–738. [Google Scholar] [CrossRef] [PubMed]
- Lofton-Day, C.; Model, F.; DeVos, T.; Tetzner, R.; Distler, J.; Schuster, M.; Song, X.; Lesche, R.; Liebenberg, V.; Ebert, M.; et al. DNA methylation biomarkers for blood-based colorectal cancer screening. Clin. Chem. 2008, 54, 414–423. [Google Scholar] [CrossRef] [PubMed]
- DeVos, T.; Tetzner, R.; Model, F.; Weiss, G.; Schuster, M.; Ditler, J.; Steiger, K.V.; Grützmann, R.; Pilarsky, C.; Habermann, J.K.; et al. Circulating methylated SEPT9 DNA in plasma is a biomarker for colorectal cancer. Clin. Chem. 2009, 55, 1337–1346. [Google Scholar] [CrossRef] [PubMed]
- Ahlquist, D.A.; Taylor, W.R.; Mahoney, D.W.; Zou, H.; Domanico, M.; Thibodeau, S.N.; Boardman, L.A.; Berger, B.M.; Lidgard, G.P. The stool DNA Test is more accurate than the plasma septin9 test in detecting colorectal Neoplasia. Clin. Gastroenterol. Hepatol. 2012, 10, 272–277. [Google Scholar] [CrossRef] [PubMed]
- Gyparaki, M.-T.; Basdra, E.K.; Papavassiliou, A.G. DNA methylation biomarkers as diagnostic and prognostic tools in colorectal cancer. J. Mol. Med. 2013, 91, 1249–1256. [Google Scholar] [CrossRef] [PubMed]
- Friso, S.; Udali, S.; Guarini, P.; Pellegrini, C.; Pattini, P.; Moruzzi, S.; Girelli, D.; Pizzolo, F.; Martinelli, N.; Corrocher, R.; et al. Global DNA hypomethylation in peripheral blood mononuclear cells as bimarkers of cancer risk. Cancer Epidemiol. Biomark. Prev. 2013, 22, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Maffei, F.; Zolezzi Moraga, J.M.; Angelini, S.; Zenesini, C.; Musti, M.; Festi, D.; Cantelli-Forti, G.; Hrelia, P. Micronucleus frequency in human peripheral blood lymphocytes as a biomarker for the early detection of colorectal cancer risk. Mutagenesis 2014, 29, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Karaman, A.; Binici, D.N.; Kabalar, M.E.; Calikuşu, Z. Micronucleus analysis in patients with colorectal adenocarcinoma and colorectal polyps. World J. Gastroenterol. 2008, 14, 6835–6839. [Google Scholar] [CrossRef] [PubMed]
- Botto, N.; Andreassi, M.G.; Manfredi, S.; Masetti, S.; Cocci, F.; Colombo, M.G.; Storti, S.; Rizza, A.; Biagini, A. Genetic polymorphisms in folate and homocysteine metabolism as risk factor for DNA damage. Eur. J. Hum. Genet. 2003, 11, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, V.; Thomas, P.; Fenech, M. Effect of common polymorphisms in folate uptake and metabolism genes on frequency of micronucleated lymphocytes in a South Australian cohort. Mutat Res. 2009, 665, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Asomaning, K.; Su, L.; Wain, J.C.; Mark, E.J.; Christiani, D.C. MTHFR polymorphisms, folate intake and carcinogen DNA adducts in the lung. Int. J. Cancer 2012, 131, 1203–1209. [Google Scholar] [CrossRef] [PubMed]
- Pardini, B.; Kumar, R.; Naccarati, A.; Prasad, R.B.; Forsti, A.; Polakova, V.; Vodickova, L.; Novotny, J.; Hemminki, K.; Vodicka, P. MTHFR and MTRR genotype and haplotype analysis and colorectal cancer susceptibility in a case-control study from the Czech Republic. Mutat. Res. 2011, 721, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.C.; Chang, P.Y.; Butler, B.; Goldstein, B.Y.; Mu, L.; Cai, L.; You, N.C.; Baecker, A.; Yu, S.Z.; Heber, D.; et al. Single nucleotide polymorphisms of one-carbon metabolism and cancers of the esophagus, stomach, and liver in a Chinese population. PLoS ONE 2014, 9, e109235. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, J.C.; Levine, A.J.; Crott, J.W.; Baurley, J.; Haile, R.W. Folate-genetics and colorectal neoplasia: What we know and need to know next. Mol. Nutr. Food Res. 2013, 57, 607–627. [Google Scholar] [CrossRef] [PubMed]
- Angelini, S.; Ravegnini, G.; Nannini, M.; Bermejo, J.L.; Musti, M.; Pantaleo, M.A.; Fumagalli, E.; Venturoli, N.; Palassini, E.; Consolini, N.; et al. Folate-related polymorphisms in gastrointestinal stromal tumours: Susceptibility and correlation with tumour characteristics and clinical outcome. Eur. J. Hum. Genet. 2015, 23, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Yao, S.; Zirpoli, G.; David Cheng, T.Y.; Roberts, M.; Khoury, T.; Ciupak, G.; Davis, W.; Pawlish, K.; Jandorf, L.; et al. Genetic variants in one-carbon metabolism genes and breast cancer risk in European American and African American women. Int. J. Cancer 2015, 137, 666–677. [Google Scholar] [CrossRef] [PubMed]
- Collin, S.M.; Metcalfe, C.; Refsum, H.; Lewis, S.J.; Smith, G.D.; Cox, A.; Davis, M.; Marsden, G.; Johnston, C.; Lane, J.A.; et al. Associations of folate, vitamin B12, homocysteine, and folate-pathway polymorphisms with prostate-specific antigen velocity in men with localized prostate cancer. Cancer Epidemiol. Biomark. Prev. 2010, 19, 2833–2838. [Google Scholar] [CrossRef] [PubMed]
- Jokić, M.; Brčić-Kostić, K.; Stefulj, J.; Catela Ivković, T.; Božo, L.; Gamulin, M.; Kapitanović, S. Association of MTHFR, MTR, MTRR, RFC1, and DHFR gene polymorphisms with susceptibility to sporadic colon cancer. DNA Cell Biol. 2011, 30, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.J.; Jin, T.B.; Yin, J.K.; Du, X.L.; Wang, Q.; Dong, R.; Wang, S.Z.; Cui, Y.; Chen, C.; Lu, J.G.; et al. Polymorphisms in tumor related genes IL-10, PSCA, MTRR and NOC3L are associated with the risk of gastric cancer in the Chinese Han population. Cancer Epidemiol. 2012, 36, e366–e372. [Google Scholar] [CrossRef] [PubMed]
- Fang, D.H.; Ji, Q.; Fan, C.H.; An, Q.; Li, J. Methionine synthase reductase A66G polymorphisms and leukaemia risk: Evidence from published studies. Leuk. Lymphoma 2014, 55, 1910–1914. [Google Scholar] [CrossRef] [PubMed]
- Barault, L.; Charon-Barra, C.; Jooste, V.; de la Vega, M.F.; Martin, L.; Roignot, P.; Rat, P.; Bouvier, A.M.; Laurent-Puig, P.; Faivre, J.; et al. Hypermethylator phenotype in sporadic colon cancer: Study on a population-based series of 582 cases. Cancer Res. 2008, 68, 8541–8546. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Sun, L.; Yang, J.; Li, Y.Y. 3R variant of thymidylate synthase 5′-untranslated enhanced region contributes to colorectal cancer risk: A meta-analysis. Asian Pac. J. Cancer Prev. 2012, 13, 2605–2610. [Google Scholar] [CrossRef] [PubMed]
- Coppedè, F.; Migheli, F.; Lopomo, A.; Failli, A.; Legitimo, A.; Consolini, R.; Fontanini, G.; Sensi, E.; Servadio, A.; Seccia, M.; et al. Gene promoter methylation in colorectal cancer and healthy adjacent mucosa specimens: Correlation with physiological and pathological characteristics, and with biomarkers of one-carbon metabolism. Epigenetics 2014, 9, 621–633. [Google Scholar] [CrossRef] [PubMed]
- De Vogel, S.; Wouters, K.A.; Gottschalk, R.W.; van Schooten, F.J.; de Goeij, A.F.; de Bruïne, A.P.; Goldbohm, R.A.; van den Brandt, P.A.; Weijenberg, M.P.; van Engeland, M. Genetic variants of methyl metabolizing enzymes and epigenetic regulators: Associations with promoter CpG island hypermethylation in colorectal cancer. Cancer Epidemiol. Biomark. Prev. 2009, 18, 3086–3096. [Google Scholar] [CrossRef] [PubMed]
- Wettergren, Y.; Odin, E.; Carlsson, G.; Gustavsson, B. MTHFR, MTR, and MTRR polymorphisms in relation to p16INK4A hypermethylation in mucosa of patients with colorectal cancer. Mol. Med. 2010, 16, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Park, H.M.; Choi, Y.K.; Chong, S.Y.; Oh, D.; Kim, N.K. Polymorphisms in genes involved in folate metabolism and plasma DNA methylation in colorectal cancer patients. Oncol. Rep. 2011, 25, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Bleich, S.; Semmler, A.; Frieling, H.; Thumfart, L.; Muschler, M.; Hillemacher, T.; Kornhuber, J.; Kallweit, U.; Simon, M.; Linnebank, M. Genetic variants of methionine metabolism and DNA methylation. Epigenomics 2014, 6, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Fuso, A.; Ferraguti, G.; Scarpa, S.; Ferrer, I.; Lucarelli, M. Disclosing bias in bisulfite assay: MethPrimers underestimate high DNA methylation. PLoS ONE 2015, 10, e0118318. [Google Scholar] [CrossRef] [PubMed]
- Choy, M.K.; Movassag, M.; Goh, H.G.; Bennett, M.; Down, T.; Foo, R. Genome-wide conserved consensus transcritpion factor binding motifs are hypermethylated. BMC Genom. 2010, 11, 519. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Song, L.; Gong, Y.; He, B. Detection of colorectal cancer by DNA methylation biomarers SEPT9: Past, present and future. Biomark. Med. 2014, 8, 755–769. [Google Scholar] [CrossRef] [PubMed]
- Zorzi, M.; Fedato, C.; Grazzini, G.; Sassoli de Bianchi, P.; Naldoni, C.; Pendenza, M.; Sassatelli, R.; Senore, C.; Visioli, C.B.; Zappa, M. Screening for colorectal cancer in Italy, 2010 survey. Epidemiol. Prev. 2012, 36, 55–77. [Google Scholar] [PubMed]
- Lehmann, U.; Kreipe, H. Real-Time PCR-Based Assay for Quantitative Determination of Methylation Status in Epigenetic Protocols; Humana Press: New York, NY, USA, 2004; pp. 207–218. [Google Scholar]
- Angelini, S.; Kumar, R.; Carbone, F.; Bermejo, J.L.; Maffei, F.; Cantelli-Forti, G.; Hemminki, K.; Hrelia, P. Inherited susceptibility to bleomycin-induced micronuclei: Correlating polymorphisms in GSTT1, GSTM1 and DNA repair genes with mutagen sensitivity. Mutat. Res. 2008, 638, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, H.; Fujimaki, C.; Daimon, T.; Tsuboi, S.; Matsuyama, T.; Itoh, K. Genetic polymorphisms in folate pathway enzymes as a possible marker for predicting the outcome of methotrexate therapy in Japanese patients with rheumatoid arthritis. J. Clin. Pharm. Ther. 2009, 34, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Johnson, W.G.; Stenroos, E.S.; Spychala, J.R.; Chatkupt, S.; Ming, S.X.; Buyske, S. New 19 bp deletion polymorphism in intron-1 of dihydrofolate reductase (DHFR): A risk factor for spina bifida acting in mothers during pregnancy? Am. J. Med. Genet. A 2004, 124A, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Hishida, A.; Matsuo, K.; Hamajima, N.; Ito, H.; Ogura, M.; Kagami, Y.; Taji, H.; Morishima, Y.; Emi, N.; Tajima, K. Associations between polymorphisms in the thymidylate synthase and serine hydroxymethyltransferase genes and susceptibility to malignant lymphoma. Haematologica 2003, 88, 159–166. [Google Scholar] [PubMed]
- Stata Corporation. Stata Statistical Software, Release 12; Stata Corporation: College Station, TX, USA, 2011. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ravegnini, G.; Zolezzi Moraga, J.M.; Maffei, F.; Musti, M.; Zenesini, C.; Simeon, V.; Sammarini, G.; Festi, D.; Hrelia, P.; Angelini, S. Simultaneous Analysis of SEPT9 Promoter Methylation Status, Micronuclei Frequency, and Folate-Related Gene Polymorphisms: The Potential for a Novel Blood-Based Colorectal Cancer Biomarker. Int. J. Mol. Sci. 2015, 16, 28486-28497. https://doi.org/10.3390/ijms161226113
Ravegnini G, Zolezzi Moraga JM, Maffei F, Musti M, Zenesini C, Simeon V, Sammarini G, Festi D, Hrelia P, Angelini S. Simultaneous Analysis of SEPT9 Promoter Methylation Status, Micronuclei Frequency, and Folate-Related Gene Polymorphisms: The Potential for a Novel Blood-Based Colorectal Cancer Biomarker. International Journal of Molecular Sciences. 2015; 16(12):28486-28497. https://doi.org/10.3390/ijms161226113
Chicago/Turabian StyleRavegnini, Gloria, Juan Manuel Zolezzi Moraga, Francesca Maffei, Muriel Musti, Corrado Zenesini, Vittorio Simeon, Giulia Sammarini, Davide Festi, Patrizia Hrelia, and Sabrina Angelini. 2015. "Simultaneous Analysis of SEPT9 Promoter Methylation Status, Micronuclei Frequency, and Folate-Related Gene Polymorphisms: The Potential for a Novel Blood-Based Colorectal Cancer Biomarker" International Journal of Molecular Sciences 16, no. 12: 28486-28497. https://doi.org/10.3390/ijms161226113