
Exploring Prostate Cancer Genome Reveals Simultaneous Losses of PTEN, FAS and PAPSS2 in Patients with PSA Recurrence after Radical Prostatectomy

Abstract
:1. Introduction
2. Results and Discussion
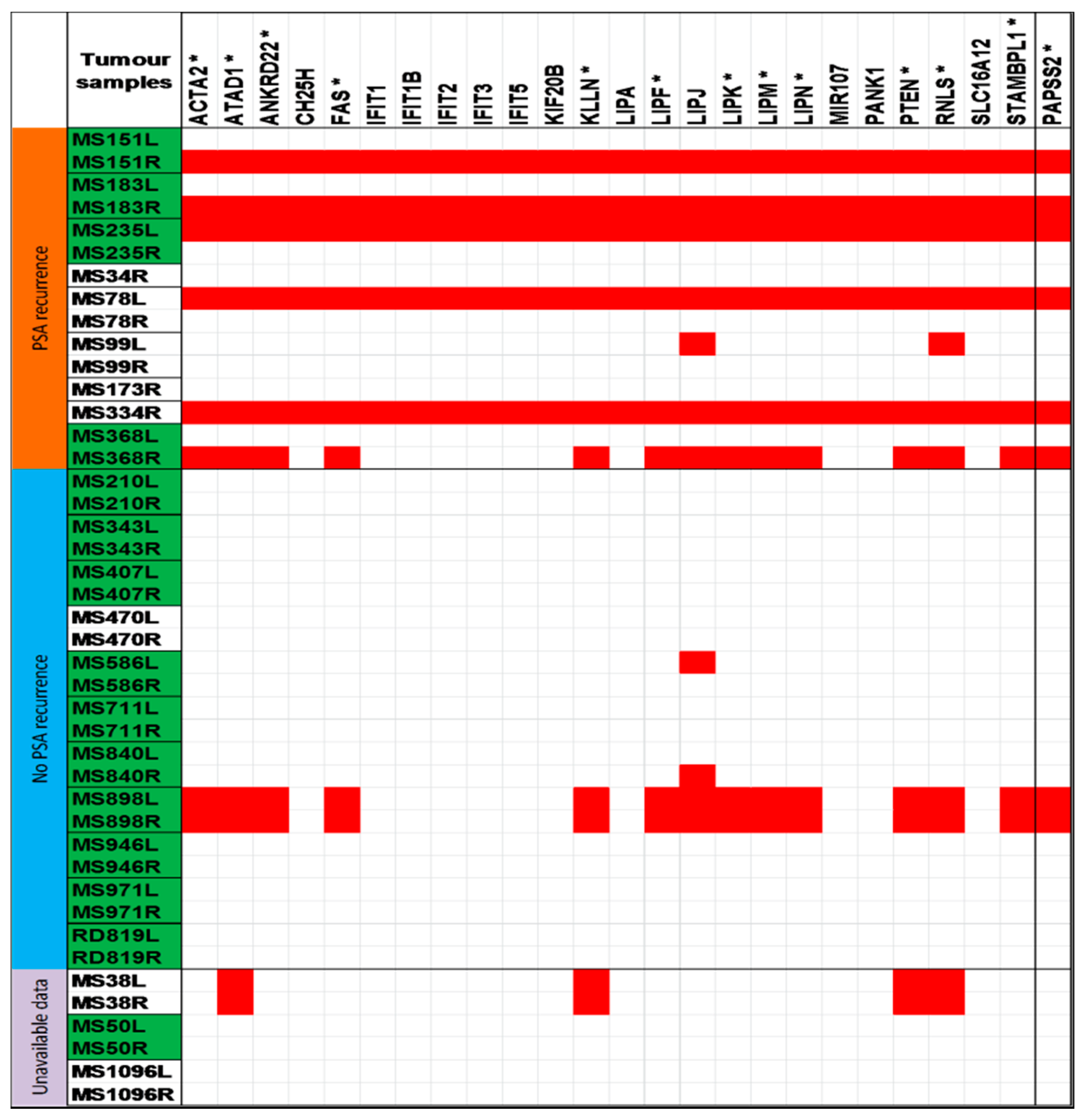
2.1. Chromosomal Copy Number Variation (CNV) Events

{kind=link}
{kind=link}
| Copy Number Variation Regions | Number of Individual Tumor Foci (n = 43) | Genes Annotated to the Region |
|---|---|---|
| Gain | ||
| 22q11.1 | 16 (37.2%) | OR11H1, POTEH |
| 16p12.1 | 13 (30.2%) | GSG1L |
| 15q22.31 | 11 (25.6%) | MEGF11 |
| 9q21.11 | 10 (23.3%) | ANKRD20A4, C9orf71, CBWD3, CBWD5, CBWD6, FAM122A, FOXD4L3, FOXD4L5, FOXD4L6, PGM5 |
| 8q21.11 | 7 (16.3%) | CRISPLD1, GDAP1, HNF4G, JPH1, LY96, MIR2052, PI15, RDH10, RPL7, STAU2, UBE2W |
| 8q22.3 | 7 (16.3%) | ATP6V1C1, AZIN1, C8orf56, CTHRC1, DPYS, FZD6, GRHL2, KLF10, LRP12, MIR3151, NCALD, ODF1, SLC25A32, TM7SF4 |
| 8q24.13 | 6 (14%) | ANXA13, ATAD2, C8orf76, DERL1, FAM83A, FAM91A1, FBXO32, FER1L6, KIAA0196, KLHL38, MTSS1, NDUFB9, NSMCE2, RNF139, SQLE, TATDN1, TMEM65, TRIB1, TRMT12, WDR67, WDYHV1, ZHX1, ZHX1-C8ORF76, ZHX2, ZNF572 |
| Loss | ||
| 8p21.2 | 21 (48.8%) | ADAM28, ADAMDEC1, BNIP3L, CDCA2, DPYSL2, KCTD9, NEFL, NEFM, NKX2-6, NKX3-1, PNMA2, PPP2R2A, STC1 |
| 8p21.3 | 21 (48.8%) | ATP6V1B2, BIN3, BMP1, C8orf58, CHMP7, CSGALNACT1, DOK2, EGR3, EPB49, FAM160B2, FGF17, GFRA2, HR, INTS10, KIAA1967, LGI3, LPL, LZTS1, MIR320A, NPM2, NUDT18, PDLIM2, PEBP4, PHYHIP, PIWIL2, POLR3D, PPP3CC, R3HCC1, REEP4, RHOBTB2, SFTPC, SH2D4A, SLC18A1, SLC39A14, SORBS3, TNFRSF10A, TNFRSF10B, TNFRSF10C, TNFRSF10D, XPO7 |
| 8p21.2–8p21.1 | 19 (44.2%) | CHRNA2, EPHX2, PTK2B, STMN4, TRIM35 |
| 8p21.3–8p21.2 | 18 (41.9%) | ENTPD4 |
| 8p23.1–8p22 | 15 (34.9%) | C8orf48, CTSB, DEFB130, DEFB134, DEFB135, DEFB136, DLC1, FAM66A, FAM66D, FAM86B1, FAM86B2, FDFT1, KIAA1456, LONRF1, MIR3926-1, MIR3926-2, USP17L2, ZNF705D |
| 16q24.1 | 14 (32.6%) | ATP2C2, C16orf74, COTL1, COX4I1, COX4NB, CRISPLD2, FAM92B, FOXC2, FOXF1, FOXL1, GINS2, IRF8, KCNG4, KIAA0182, KIAA0513, KIAA1609, MIR1910, MTHFSD, USP10, WFDC1, ZDHHC7 |
| 18q11.2 | 14 (32.6%) | ANKRD29, AQP4, C18orf45, C18orf8, CABYR, CHST9, HRH4, IMPACT, KCTD1, LAMA3, MIR320C2, NPC1, OSBPL1A, PSMA8, RIOK3, SS18, TAF4B, TTC39C, ZNF521 |
| 8p11.22–8p11.21 | 13 (30.2%) | ADAM2, ADAM18 |
| 10q23.31 | 13 (30.2%) | ACTA2, ANKRD22, ATAD1, CH25H, FAS, IFIT1, IFIT1B, IFIT2, IFIT3, IFIT5, KIF20B, KLLN, LIPA, LIPF, LIPJ, LIPK, LIPM, LIPN, MIR107, PANK1, PTEN, RNLS, SLC16A12, STAMBPL1 |
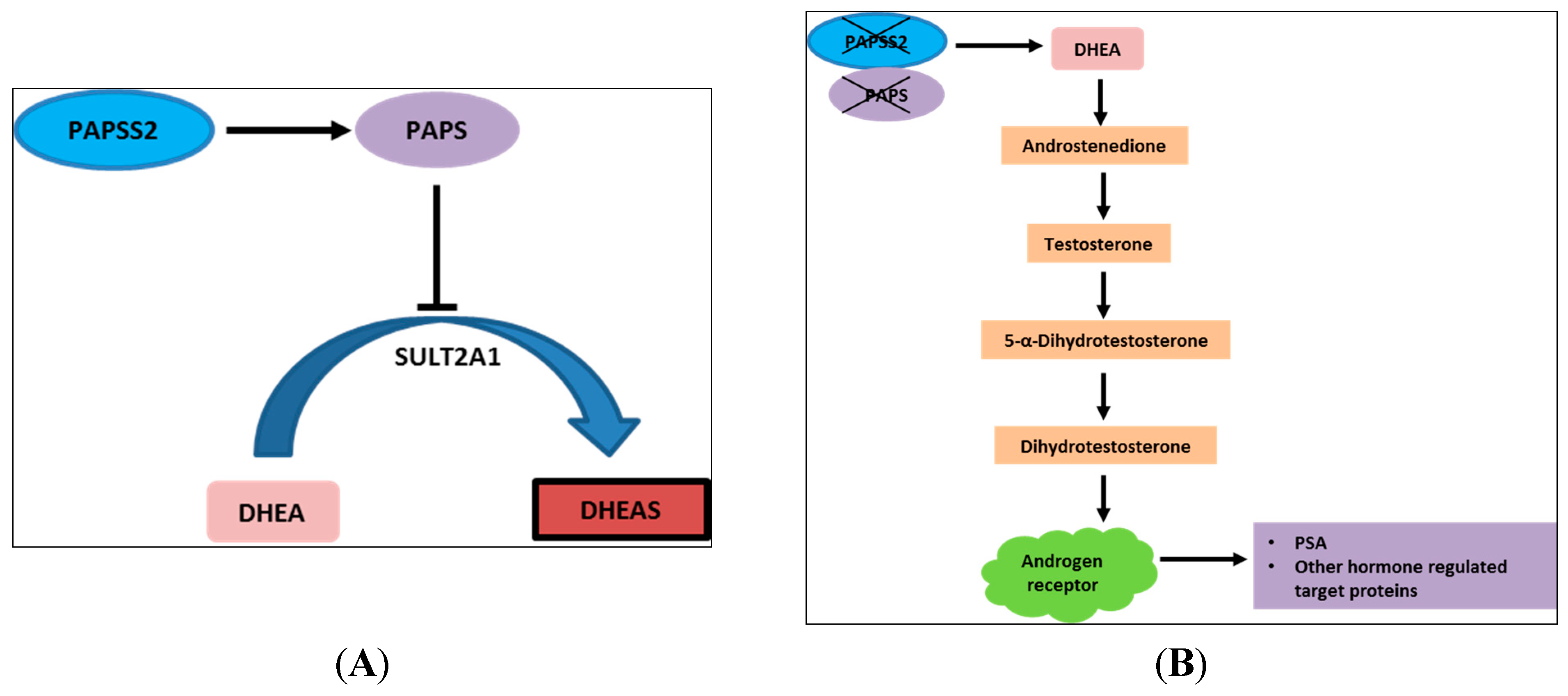
| 10q23.2–10q23.31 | 10 (23.3%) | PAPSS2 |
2.2. Establishing Relationship between Copy Number Variation Regions and PSA Recurrence
| Regions with Copy Number Loss Genomic Position (Start–End) a | Genes Implicated b | PSA Recurrence | |
|---|---|---|---|
| Yes (n = 9) | No (n = 11) | ||
| 10q23.2–10q23.31 chr. 10: 89351602–89572982 | PAPSS2 | 6/9 (66.7%) * | 1/11 (9.1%) |
| 10q23.31 chr. 10: 89572982–91817088 | ACTA2, ANKRD22, ATAD1, CH25H, FAS, IFIT1, IFIT1B, IFIT2, IFIT3, IFIT5, KIF20B, KLLN, LIPA, LIPF, LIPJ, LIPK, LIPM, LIPN, MIR107, PANK1, PTEN, RNLS, SLC16A12, STAMBPL1 | 7/9 (77.8%) | 3/11(27.3%) |

2.3. Discussion

3. Experimental Section
3.1. Patients
| Clinical Parameter | Status | n (%) |
|---|---|---|
| Age (years) | <60 | 10 (43.5%) |
| >60 | 13 (56.5%) | |
| Focality | Multifocal | 15 (65%) |
| Unifocal | 8 (35%) | |
| Pathological tumor stage (pT) | pT2 | 5 (21.7%) |
| pT3 | 17 (73.9%) | |
| pT4 | 1 (4.35%) | |
| Gleason score | ≤7 | 22 (51.2%) |
| >7 | 21 (48.8%) | |
| PSA recurrence * | yes | 9 (45%) |
| no | 11 (55%) |
3.2. Clinical Specimens
3.3. Quality Control and Data Analysis
3.4. Statistical Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, H.; DeAngelis, G.; Eltze, E.; Gockel, I.; Semjonow, A.; Brandt, B. Asynchronous growth of prostate cancer is reflected by circulating tumor cells delivered from distinct, even small foci, harboring loss of heterozygosity of the PTEN Gene. Cancer Res. 2006, 66, 8959–8965. [Google Scholar] [CrossRef] [PubMed]
- Andreoiu, M.; Cheng, L. Multifocal prostate cancer: Biologic, prognostic, and therapeutic implications. Hum. Pathol. 2010, 41, 781–793. [Google Scholar] [CrossRef] [PubMed]
- Kluth, M.; Hesse, J.; Heinl, A.; Krohn, A.; Steurer, S.; Sirma, H.; Simon, R.; Mayer, P.S.; Schumacher, U.; Grupp, K.; et al. Genomic deletion of MAP3K7 at 6q12–22 is associated with early PSA recurrence in prostate cancer and absence of TMPRSS2: ERG fusions. Mod. Pathol. 2013, 26, 975–983. [Google Scholar]
- Taylor, B.S.; Schultz, N.; Hieronymus, H.; Gopalan, A.; Xiao, Y.; Carver, B.S.; Arora, V.K.; Kaushik, P.; Cerami, E.; Reva, B.; et al. Integrative genomic profiling of human prostate cancer. Cancer Cell 2010, 18, 11–22. [Google Scholar]
- Jenster, G. The role of the androgen receptor in the development and progression of prostate cancer. Semin. Oncol. 1999, 26, 407–421. [Google Scholar] [PubMed]
- Baca, S.C.; Garraway, L.A. The genomic landscape of prostate cancer. Front. Endocrinol. 2012, 3. [Google Scholar] [CrossRef]
- Francis, J.C.; Thomsen, M.K.; Taketo, M.M.; Swain, A. β-Catenin is required for prostate development and cooperates with Pten loss to drive invasive carcinoma. PLoS Genet. 2013, 9, e1003180. [Google Scholar] [CrossRef] [PubMed]
- Kluth, M.; Harasimowicz, S.; Burkhardt, L.; Grupp, K.; Krohn, A.; Prien, K.; Gjoni, J.; Hass, T.; Galal, R.; Graefen, M.; et al. Clinical significance of different types of P53 Gene alteration in surgically treated prostate cancer. Int. J. Cancer 2014, 135, 1369–1380. [Google Scholar]
- Krohn, A.; Diedler, T.; Burkhardt, L.; Mayer, P.S.; de Silva, C.; Meyer-Kornblum, M.; Kotschau, D.; Tennstedt, P.; Huang, J.; Gerhauser, C.; et al. Genomic deletion of PTEN is associated with tumor progression and early PSA recurrence in erg fusion-positive and fusion-negative prostate cancer. Am. J. Pathol. 2012, 181, 401–412. [Google Scholar]
- Liu, W.; Xie, C.C.; Thomas, C.Y.; Kim, S.T.; Lindberg, J.; Egevad, L.; Wang, Z.; Zhang, Z.; Sun, J.; Sun, J.; et al. Genetic markers associated with early cancer-specific mortality following prostatectomy. Cancer 2013, 119, 2405–2412. [Google Scholar]
- Chaux, A.; Peskoe, S.B.; Gonzalez-Roibon, N.; Schultz, L.; Albadine, R.; Hicks, J.; de Marzo, A.M.; Platz, E.A.; Netto, G.J. Loss of PTEN expression is associated with increased risk of recurrence after prostatectomy for clinically localized prostate cancer. Mod. Pathol. 2012, 25, 1543–1549. [Google Scholar] [CrossRef] [PubMed]
- Ibeawuchi, C.; Schmidt, H.; Voss, R.; Titze, U.; Abbas, M.; Neumann, J.; Eltze, E.; Hoogland, A.M.; Jenster, G.; Brandt, B.; et al. Genome-wide investigation of multifocal and unifocal prostate cancer-are they genetically different? Int. J. Mol. Sci. 2013, 14, 11816–11829. [Google Scholar]
- Lindberg, J.; Klevebring, D.; Liu, W.; Neiman, M.; Xu, J.; Wiklund, P.; Wiklund, F.; Mills, I.G.; Egevad, L.; Gronberg, H. Exome sequencing of prostate cancer supports the hypothesis of independent tumour origins. Eur. Urol. 2013, 63, 347–353. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, G.; Rittenhouse, H.G.; Mikolajczyk, S.D.; Blair Shamel, L.; Semjonow, A. Twenty years of PSA: From prostate antigen to tumor marker. Rev. Urol. 2007, 9, 113–123. [Google Scholar] [PubMed]
- Visakorpi, T.; Kallioniemi, A.H.; Syvanen, A.C.; Hyytinen, E.R.; Karhu, R.; Tammela, T.; Isola, J.J.; Kallioniemi, O.P. Genetic changes in primary and recurrent prostate cancer by comparative genomic hybridization. Cancer Res. 1995, 55, 342–347. [Google Scholar] [PubMed]
- Barbieri, C.E.; Baca, S.C.; Lawrence, M.S.; Demichelis, F.; Blattner, M.; Theurillat, J.P.; White, T.A.; Stojanov, P.; van Allen, E.; Stransky, N.; et al. Exome sequencing identifies recurrent SPOP, FOXA1 and MED12 mutations in prostate cancer. Nat. Genet. 2012, 44, 685–689. [Google Scholar]
- Cheng, I.; Levin, A.M.; Tai, Y.C.; Plummer, S.; Chen, G.K.; Neslund-Dudas, C.; Casey, G.; Rybicki, B.A.; Witte, J.S. Copy number alterations in prostate tumors and disease aggressiveness. Genes Chromosomes Cancer 2012, 51, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Bettendorf, O.; Schmidt, H.; Staebler, A.; Grobholz, R.; Heinecke, A.; Boecker, W.; Hertle, L.; Semjonow, A. Chromosomal imbalances, loss of heterozygosity, and immunohistochemical expression of TP53, RB1, and PTEN in intraductal cancer, intraepithelial neoplasia, and invasive adenocarcinoma of the prostate. Genes Chromosomes Cancer 2008, 47, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Chang, B.L.; Liu, W.; Sun, J.; Dimitrov, L.; Li, T.; Turner, A.R.; Zheng, S.L.; Isaacs, W.B.; Xu, J. Integration of somatic deletion analysis of prostate cancers and germline linkage analysis of prostate cancer families reveals two small consensus regions for prostate cancer genes at 8p. Cancer Res. 2007, 67, 4098–4103. [Google Scholar] [CrossRef] [PubMed]
- Bhatia-Gaur, R.; Donjacour, A.A.; Sciavolino, P.J.; Kim, M.; Desai, N.; Young, P.; Norton, C.R.; Gridley, T.; Cardiff, R.D.; Cunha, G.R.; et al. Roles for Nkx3.1 in prostate development and cancer. Genes Dev. 1999, 13, 966–977. [Google Scholar]
- Shen, M.M.; Abate-Shen, C. Roles of the Nkx3.1 homeobox gene in prostate organogenesis and carcinogenesis. Dev. Dyn. 2003, 228, 767–778. [Google Scholar]
- Reid, A.H.; Attard, G.; Brewer, D.; Miranda, S.; Riisnaes, R.; Clark, J.; Hylands, L.; Merson, S.; Vergis, R.; Jameson, C.; et al. Novel, Gross chromosomal alterations involving PTEN cooperate with allelic loss in prostate cancer. Mod. Pathol. 2012, 25, 902–910. [Google Scholar]
- Haffner, M.C.; Mosbruger, T.; Esopi, D.M.; Fedor, H.; Heaphy, C.M.; Walker, D.A.; Adejola, N.; Gurel, M.; Hicks, J.; Meeker, A.K.; et al. Tracking the clonal origin of lethal prostate cancer. J. Clin. Investig. 2013, 123, 4918–4922. [Google Scholar]
- Di Cristofano, A.; Pandolfi, P.P. The Multiple roles of PTEN in tumor suppression. Cell 2000, 100, 387–390. [Google Scholar] [CrossRef] [PubMed]
- Pourmand, G.; Ziaee, A.A.; Abedi, A.R.; Mehrsai, A.; Alavi, H.A.; Ahmadi, A.; Saadati, H.R. Role of PTEN gene in progression of prostate cancer. Urol. J. 2007, 4, 95–100. [Google Scholar] [PubMed]
- Lima, L.; Morais, A.; Lobo, F.; Calais-da-Silva, F.M.; Calais-da-Silva, F.E.; Medeiros, R. Association between FAS polymorphism and prostate cancer development. Prostate Cancer Prostatic Dis. 2008, 11, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Takayama, H.; Takakuwa, T.; Dong, Z.; Nonomura, N.; Okuyama, A.; Nagata, S.; Aozasa, K. Fas gene mutations in prostatic intraepithelial neoplasia and concurrent carcinoma: Analysis of laser capture microdissected specimens. Lab. Investig. 2001, 81, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Park, S.M.; Tumanov, A.V.; Hau, A.; Sawada, K.; Feig, C.; Turner, J.R.; Fu, Y.X.; Romero, I.L.; Lengyel, E.; et al. CD95 promotes tumour growth. Nature 2010, 465, 492–496. [Google Scholar]
- Yuan, X.J.; Whang, Y.E. PTEN sensitizes prostate cancer cells to death receptor-mediated and drug-induced apoptosis through a FADD-dependent pathway. Oncogene 2002, 21, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Faiyaz ul Haque, M.; King, L.M.; Krakow, D.; Cantor, R.M.; Rusiniak, M.E.; Swank, R.T.; Superti-Furga, A.; Haque, S.; Abbas, H.; Ahmad, W.; et al. Mutations in orthologous genes in human spondyloepimetaphyseal dysplasia and the brachymorphic mouse. Nat. Genet. 1998, 20, 157–162. [Google Scholar]
- Xu, Z.H.; Freimuth, R.R.; Eckloff, B.; Wieben, E.; Weinshilboum, R.M. Human 3'-phosphoadenosine 5'-phosphosulfate synthetase 2 (PAPSS2) pharmacogenetics: Gene resequencing, genetic polymorphisms and functional characterization of variant allozymes. Pharmacogenetics 2002, 12, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Venkatachalam, K.V. Human 3'-Phosphoadenosine 5'-Phosphosulfate (PAPS) synthase: Biochemistry, molecular biology and genetic deficiency. IUBMB Life 2003, 55, 1–11. [Google Scholar] [PubMed]
- Noordam, C.; Dhir, V.; McNelis, J.C.; Schlereth, F.; Hanley, N.A.; Krone, N.; Smeitink, J.A.; Smeets, R.; Sweep, F.C.; Claahsen-van der Grinten, H.L.; et al. Inactivating PAPSS2 mutations in a patient with premature pubarche. N. Engl. J. Med. 2009, 360, 2310–2318. [Google Scholar]
- Franzon, V.L.; Gibson, M.A.; Hatzinikolas, G.; Woollatt, E.; Sutherland, G.R.; Cleary, E.G. Molecular cloning of a novel human PAPS synthetase which is differentially expressed in metastatic and non-metastatic colon carcinoma cells. Int. J. Biochem. Cell Biol. 1999, 31, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Hermans, K.G.; van Alewijk, D.C.; Veltman, J.A.; van Weerden, W.; van Kessel, A.G.; Trapman, J. Loss of a small region around the PTEN locus is a major chromosome 10 alteration in prostate cancer xenografts and cell lines. Genes Chromosomes Cancer 2004, 39, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Heinlein, C.A.; Chang, C. Androgen receptor in prostate cancer. Endocr. Rev. 2004, 25, 276–308. [Google Scholar] [CrossRef] [PubMed]
- Leube, B.; Drechsler, M.; Muhlmann, K.; Schafer, R.; Schulz, W.A.; Santourlidis, S.; Anastasiadis, A.; Ackermann, R.; Visakorpi, T.; Muller, W.; et al. Refined mapping of allele loss at chromosome 10q23–26 in prostate cancer. Prostate 2002, 50, 135–144. [Google Scholar]
- Attard, G.; Reid, A.H.; Olmos, D.; de Bono, J.S. Antitumor activity with CYP17 blockade indicates that castration-resistant prostate cancer frequently remains hormone driven. Cancer Res. 2009, 69, 4937–4940. [Google Scholar] [CrossRef] [PubMed]
- Eminaga, O.; Abbas, M.; Hinkelammert, R.; Titze, U.; Bettendorf, O.; Eltze, E.; Ozgur, E.; Semjonow, A. CMDX(c)-based single source information system for simplified quality management and clinical research in prostate cancer. BMC Med. Inform. Decis. Mak. 2012, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinkelammert, R.; Eminaga, O.; Bettendorf, O.; Eltze, E.; Abbas, M.; Hertle, L.; Semjonow, A. Tumor percentage but not number of tumor foci predicts disease-free survival after radical prostatectomy especially in high-risk patients. Urol. Oncol. 2014, 32, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Affymetrix Genotyping Console, version 4.1.2; Software for creating custom SQLite format annotations; Affymetrix: Santa Clara, CA, USA, 2011.
- Partek Genomic Suite Software, version 6.6; Software for analyzing statistics; Partek Inc.: St. Louis, MO, USA, 2012.
- Optimizing Copy Number Segmentation in Partek; Partek Inc.: St. Louis, MO, USA, 2009.
- Pruitt, K.D.; Tatusova, T.; Maglott, D.R. NCBI reference sequences (RefSeq): A curated non-redundant sequence database of genomes, transcripts and proteins. Nucleic Acids Res. 2007, 35, 61–65. [Google Scholar] [CrossRef]
- User Guide: Associating a Spreadsheet with an Annotation File in Partek Genomics Suite; Partek Inc.: St. Louis, MO, USA, 2010.
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibeawuchi, C.; Schmidt, H.; Voss, R.; Titze, U.; Abbas, M.; Neumann, J.; Eltze, E.; Hoogland, A.M.; Jenster, G.; Brandt, B.; et al. Exploring Prostate Cancer Genome Reveals Simultaneous Losses of PTEN, FAS and PAPSS2 in Patients with PSA Recurrence after Radical Prostatectomy. Int. J. Mol. Sci. 2015, 16, 3856-3869. https://doi.org/10.3390/ijms16023856
Ibeawuchi C, Schmidt H, Voss R, Titze U, Abbas M, Neumann J, Eltze E, Hoogland AM, Jenster G, Brandt B, et al. Exploring Prostate Cancer Genome Reveals Simultaneous Losses of PTEN, FAS and PAPSS2 in Patients with PSA Recurrence after Radical Prostatectomy. International Journal of Molecular Sciences. 2015; 16(2):3856-3869. https://doi.org/10.3390/ijms16023856
Chicago/Turabian StyleIbeawuchi, Chinyere, Hartmut Schmidt, Reinhard Voss, Ulf Titze, Mahmoud Abbas, Joerg Neumann, Elke Eltze, Agnes Marije Hoogland, Guido Jenster, Burkhard Brandt, and et al. 2015. "Exploring Prostate Cancer Genome Reveals Simultaneous Losses of PTEN, FAS and PAPSS2 in Patients with PSA Recurrence after Radical Prostatectomy" International Journal of Molecular Sciences 16, no. 2: 3856-3869. https://doi.org/10.3390/ijms16023856