The Influence of Environmental Temperatures on Neurotoxicity Induced by Methamphetamine in Male Rats

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Introduction
Materials and Methods
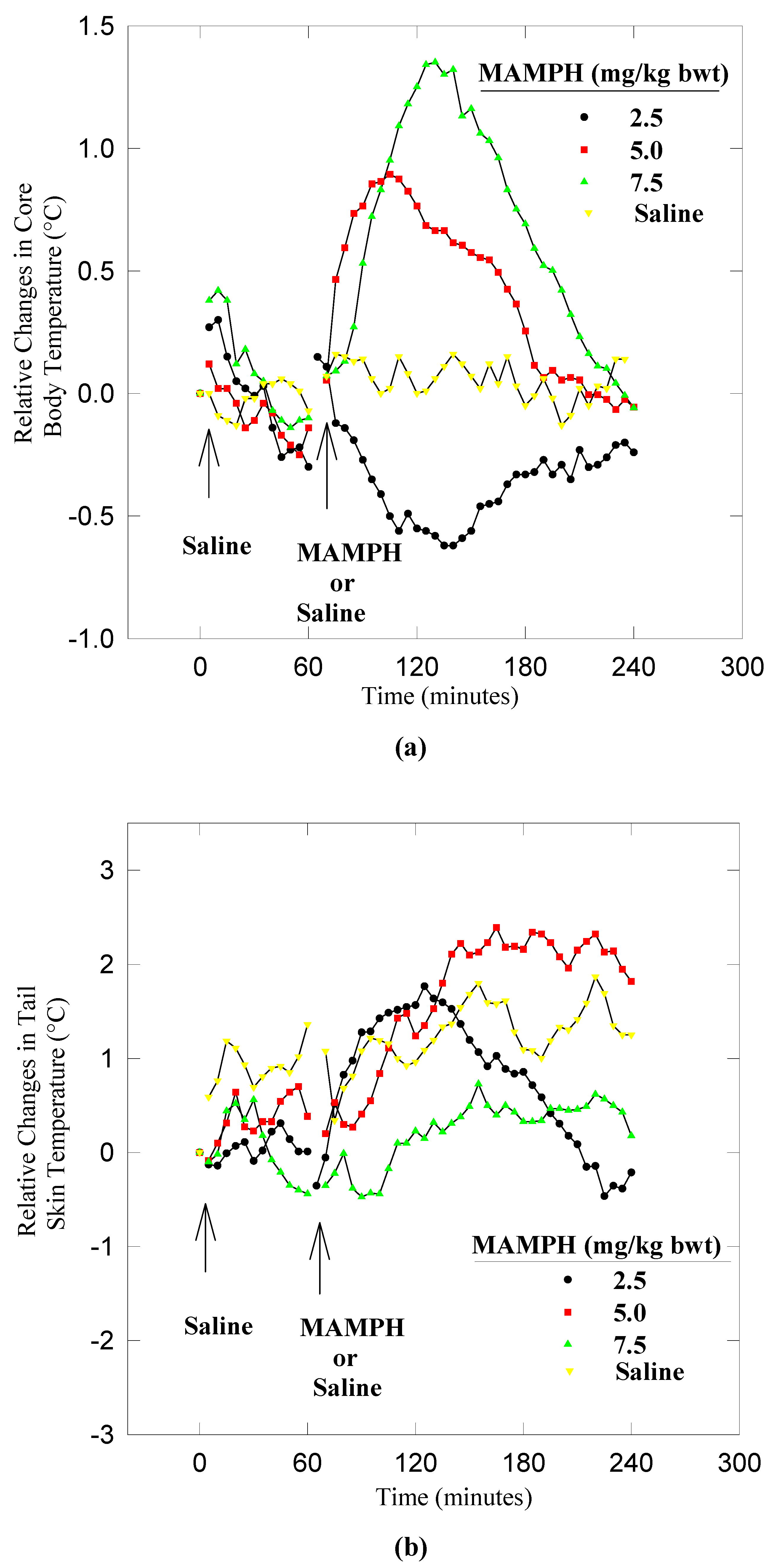
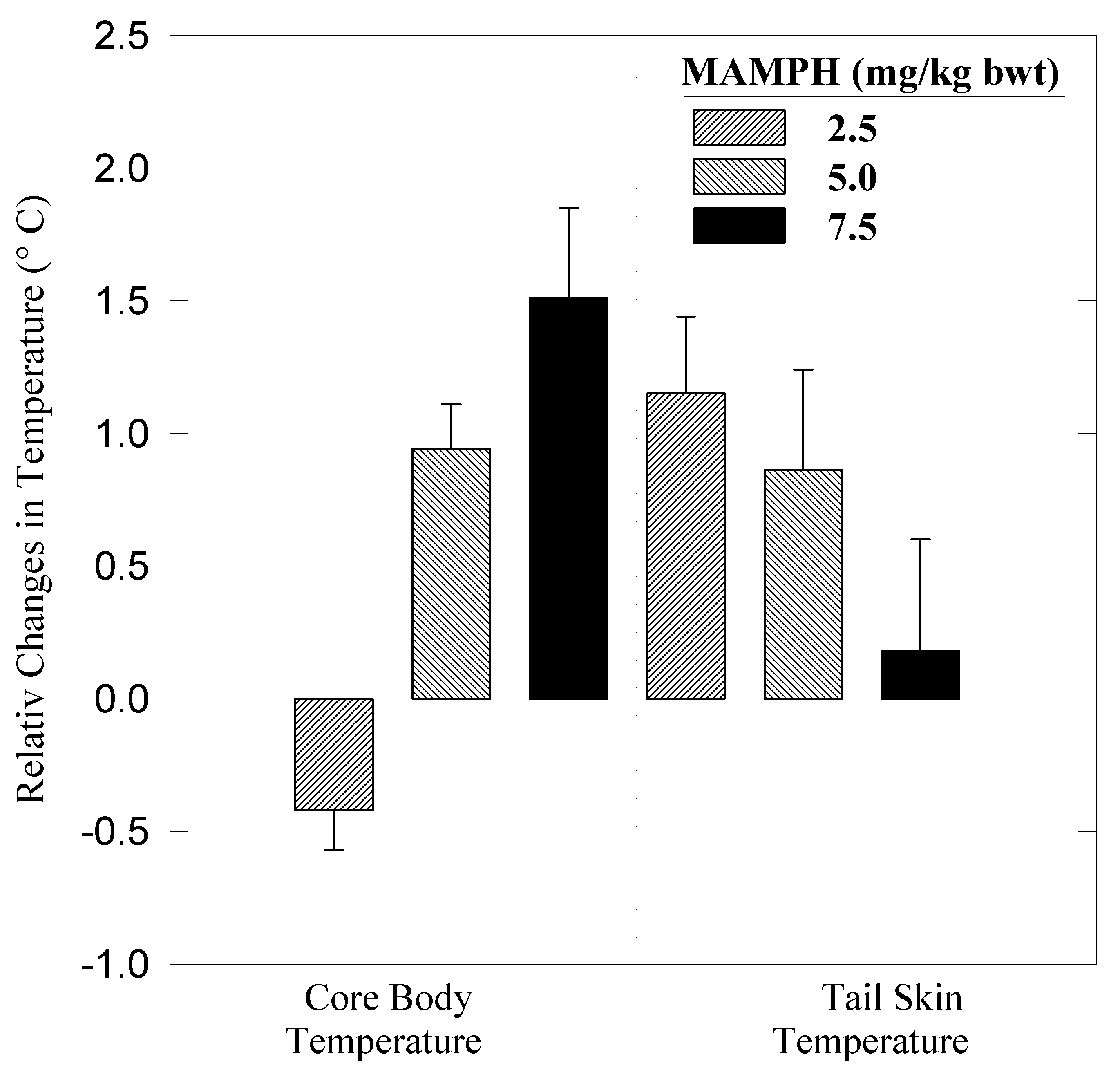
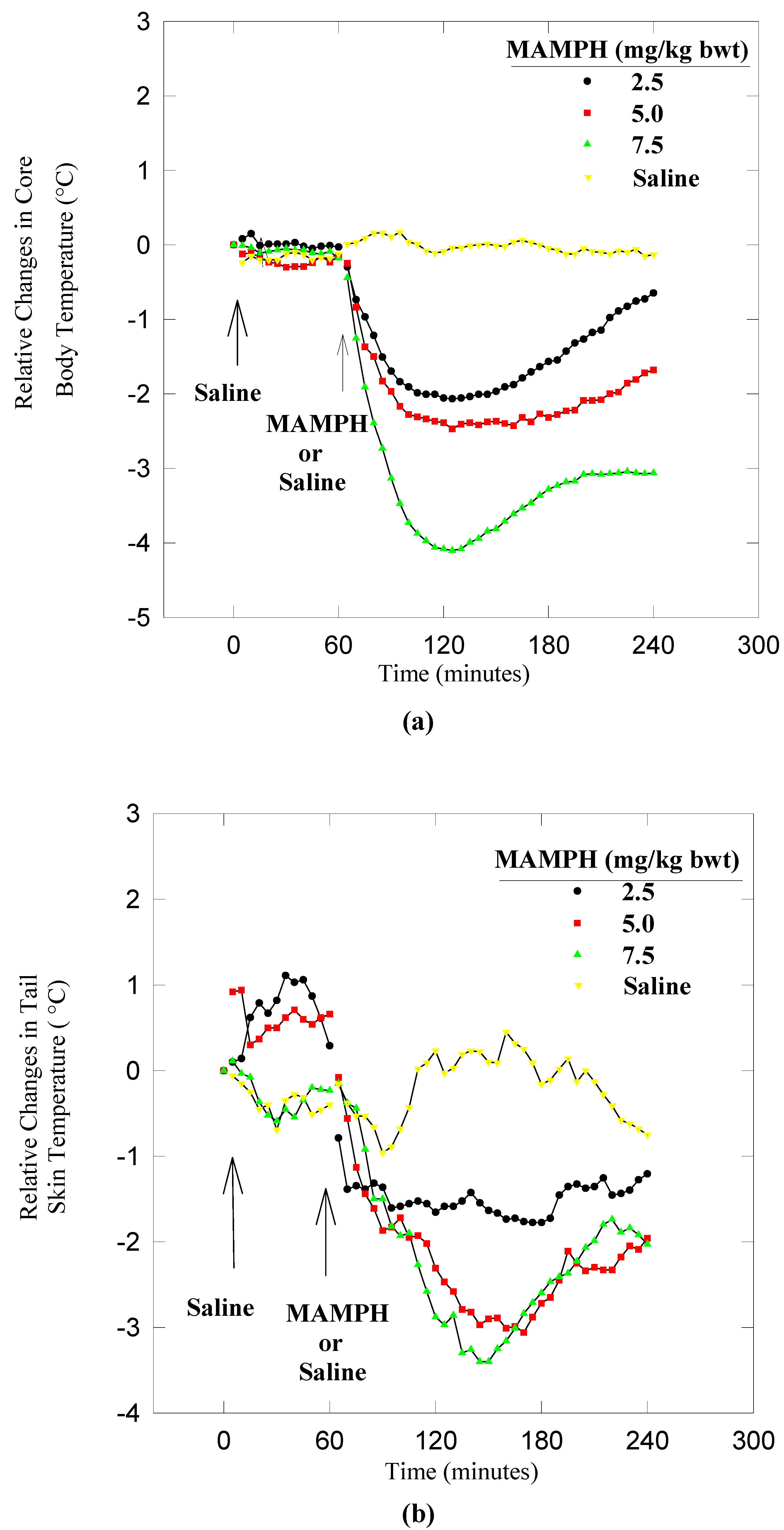
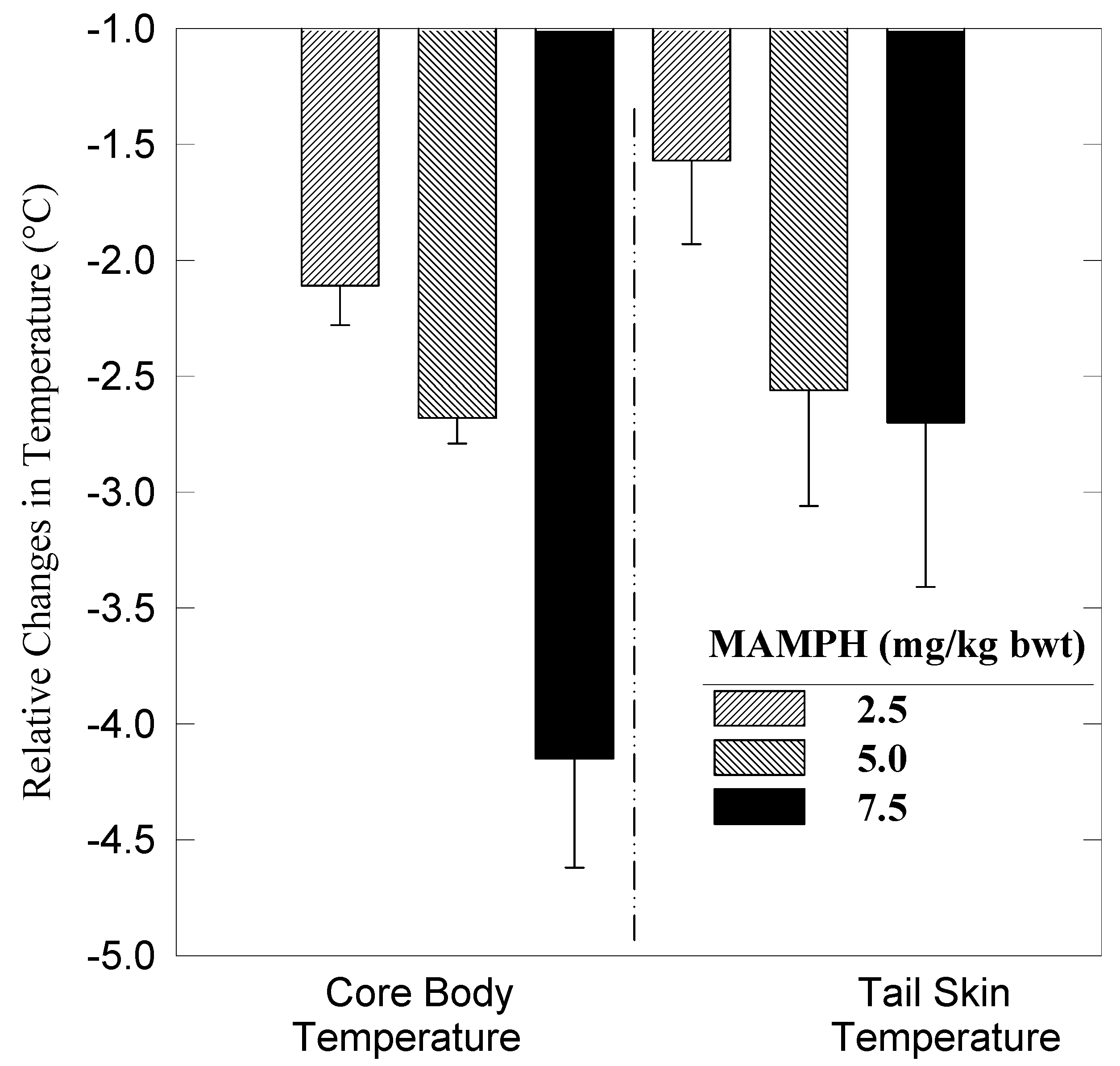
Results
Dynamics of heat dissipation


Dynamics of heat conservation


Discussion
Acknowledgement
References and Notes
- Ricuarte, G.A.; Schuster, C.R.; Seiden, L.S. Long-term effects of repeated methyl amphetamine administration on dopamine and serotonin neurons in the rat brain: A regional study. Brain Res. 1980, 19, 153–163. [Google Scholar] [CrossRef]
- Wagner, G.C.; Ricuarte, G.A.; Seiden, L.S.; Schuster, C.R.; Miller, R.J.; Westley, J. Long-lasting depletions of striatal dopamine and loss of dopamine uptake sites following repeated administration of methamphetamine. Brain Res. 1980, 181, 151–160. [Google Scholar] [CrossRef]
- Seiden, L.S. Neurotoxicity of methamphetamine and related drugs. In Psychopharmacology: A Third Generation of Progress; Meltzer, H.Y., Ed.; Raven Press: New York, 1987; pp. 359–366. [Google Scholar]
- Kogan, F.J.; Nichols, W.K.; Gibb, J.W. Influence of methamphetamine on nigral and striatal tyrosine hydroxylase activity and on striatal dopamine levels. Eur. J. Pharmacol. 1976, 36, 363–371. [Google Scholar]
- Xie, T.; McCann, U.D.; Kim, S. Yuan; Ricaurte, G.A. Effect of temperature on dopamine transporter function and intracellular accumulation of methamphetamine: Implications for methamphetamine-induced dopaminergic neurotoxicity. J. Neurosci. 2000, 20, 7838–7845. [Google Scholar]
- Brown, J.M.; Riddle, E.L.; Sandoval, V.; Weston, R.K.; Hanson, J.E.; Crosby, M.J.; Ugarte, Y.V.; Gibb, J.W.; Hanson, G.R.; Fleckenstein, A.E. A single methamphetamine administration rapidly decreases vesicular dopamine uptake. J. Pharmacol. Exp. Ther. 2002, 302, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.T.; Cord, B.J.; Yuan, J.; McCann, U.D.; Ricaurte, G.A. Inhibitors of Na (+)/H(+) and Na(+)/Ca(2+) exchange potentiate methamphetamine-induced dopamine neurotoxicity: possible role of ionic dysregulation in methamphetamine neurotoxicity. J. Neurochem. 2001, 77, 1348–62. [Google Scholar]
- Bakhit, C.; Gibb, J.W. Methamphetamine-induced depression of tryptophan hydroxylase: Recovery following acute treatment. Eur. J. Pharmacol. 1981, 76, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Trulson, M.E.; Trulson, V.M. Effects of chronic methamphetamine administration on tryptophan hydroxylase activity, [3H] serotonin synaptosomal uptake, and serotonin metabolism in rat brain following systemic tryptophan loading. Neuropharmacology 1982, 21, 521–527. [Google Scholar]
- Kuczenski, R.; Segal, D.S. In vivo measures of monoamines during amphetamine-induced behaviors in rats. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 1991, 14, S37–S50. [Google Scholar]
- Randrup, A.; Munkvad, I. Pharmacology and physiology of stereotyped behavior. J. Psychiat. Res. 1974, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bo, P.; Marchioni, E.; Dollocchio, C.; Albergati, A.; Savoldi, F. Antagonism of EEGgraphic and behavioral effects of methamphetamine by selective receptor blockers (SCH 23390 and Raclopride) in the rabbit. Prog. Neuro-Psychopharmacol. & Biol. Psychit. 1991, 15, 803–815. [Google Scholar] [CrossRef]
- Wagstaff, J.D.; Bush, L.G.; Gibb, J.W.; Hanson, G.R. Endogenous neurotensin antagonizes methamphetamine-enhanced dopaminergic activity. Brain Res. 1994, 665, 237–244. [Google Scholar]
- Segal, D.S. Kuczenski, RRepeated binge exposures to amphetamine and methamphetamine: Behavioral and neurochemical characterization. J. Pharmacol. Exp. Ther. 1997, 282, 561–573. [Google Scholar]
- Itzhak, Y. Modulation of cocaine- and Methamphetamine-induced behavioral sensitization by inhibition of brain nitric oxide synthase. J. Pharmacol. Exp. Ther. 1997, 282, 521–527. [Google Scholar]
- Yehuda, S.; Wutman, R.J. Paradoxical effects of D-amphetamine on behavioral thermoregulation: Possible mediation by brain dopamine. J. Pharmacol. Expe. Therapy. 1974, 1990, 118–122. [Google Scholar]
- Mohaghegh, R. A. Physiological phenomena underlying the acute thermic effects of methamphetamine on rats in different environmental temperatures. Ph.D. Dissertation, University of Arkansas for Medical Sciences, Little Rock, AR, USA, 1994. [Google Scholar]
- Mohaghegh, R.A.; Soulsby, M.E.; Skinner, R.D.; Kennedy, R.H. The interaction between the central and peripheral nervous systems in mediating the thermic effect of methamphetamine. Annals New York Acad. Sci. 1997, 813, 197–203. [Google Scholar] [CrossRef]
- Bowyer, J.F.; Tank, A.W.; Newport, G.D.; Slikker, W., Jr.; Ali, S.F.; Holson, R.R. The influence of environmental temperature on the transient effects of methamphetamine on dopamine levels and dopamine release in rat striatum. J. Pharmacol. Exp. Ther. 1992, 260, 817–824. [Google Scholar] [PubMed]
- Bowyer, J.F.; Gough, B.; Slikker, W., Jr.; Lipe, G.W.; Newport, G.D.; Holson, R.R. Effects of a cold environment or age on methamphetamine-induced dopamine release in the caudate putamen of female rats. Pharmacol. Biochem. Behav. 1993, 44, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Bowyer, J.F.; Davies, D.L.; Schmued, L.; Broening, H.W.; Newport, G.D.; Slikker, W., Jr.; Holson, R.R. Further studies of the role of hyperthermia in methamphetamine neurotoxicity. J. Pharmacol. Exp. Ther. 1994, 268, 1571–1580. [Google Scholar]
- Ali, S.F.; Newport, G.D.; Holson, R.R.; Slikker, W., Jr.; Bowyer, J.F. Low environmental temperatures or pharmacologic agents that produce hypothermia decrease methamphetamine neurotoxicity in mice. Brain Res. 1994, 658, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.F.; Newport, G.D.; Holson, R.R.; Slikker, W., Jr.; Bowyer, J.F. Low environmental temperatures or pharmacologic agents that produce hyperthermia decrease methamphetamine neurotoxicity in mice. Ann. NY Acad. Sci. 1995, 765, 338. [Google Scholar] [CrossRef]
- Ali, S.F.; Newport, G.D.; Slikker, W., Jr. Methamphetamine-induced dopaminergic toxicity in mice. Role of environmental and pharmacological agents. Ann. NY Acad. Sci. 1996, 801, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Raman, E.R.; Roberts, M.F.; Vanhuyse, V.J. Body temperature control of rat tail blood flow. Am. J. Physiol. 1983, 245, R426–R432. [Google Scholar]
- Johansen, R. Heat exchanger through the muskrat tail evidence for vasodilator nerves to the skin. Acta Physiol. Scan. 1962, 55, 160–169. [Google Scholar] [CrossRef]
- Rand, R.P.; Burton, A.C.; Ing, T. The tail of the rat in temperature regulation and acclimatization. Can. J. Physiol. Pharmacol. 1965, 43, 257–267. [Google Scholar] [CrossRef]
- Gordon, J.G. Relationship between preferred ambient temperature and autonomic thermoregulatory function in rat. Am. J. Physiol. 1987, 252, R1130–R1137. [Google Scholar]
- Romanovsky, A.; Ivanov, I. A.; Shimansky, P.Y. Molecular biology of thermoregulation selected contribution: ambient temperature for experiments in rats: a new method for determining the zone of thermal neutrality. J. Appl. Physiol. 2002, 92, 2667–2679. [Google Scholar]
- Lipton, J.M.; Welch, J.P.; Clark, W. G. Change in body temperature produced by injecting prostaglandin E1, EGTA and bacterial endotoxins into the PO/AH region and the medulla oblongata of the rat. Experimentia 1973, 29, S06–S08. [Google Scholar]
- Stitt, J. T. Fever versus hyperthermia. Fedn. Proc. 1979, 38, 39–43. [Google Scholar]
- Shimada, S.G.; Stitt, J.T.; Angelogianni, P. Effects of cold and capsaicin desensitization on prostaglandin E hypothermia in rats. J. Applied Physiol. 1990, 68, 2618–2622. [Google Scholar]
© 2002 by MDPI (http://www.mdpi.org).
Share and Cite
Mohaghegh, R.A.; Singleton, J.; Shockley, D. The Influence of Environmental Temperatures on Neurotoxicity Induced by Methamphetamine in Male Rats. Int. J. Mol. Sci. 2002, 3, 1095-1104. https://doi.org/10.3390/i3101095
Mohaghegh RA, Singleton J, Shockley D. The Influence of Environmental Temperatures on Neurotoxicity Induced by Methamphetamine in Male Rats. International Journal of Molecular Sciences. 2002; 3(10):1095-1104. https://doi.org/10.3390/i3101095
Chicago/Turabian StyleMohaghegh, Rymond A., Jeurel Singleton, and Dolores Shockley. 2002. "The Influence of Environmental Temperatures on Neurotoxicity Induced by Methamphetamine in Male Rats" International Journal of Molecular Sciences 3, no. 10: 1095-1104. https://doi.org/10.3390/i3101095