Preparation and Characterization of a Chloroperoxidase-like Catalytic Antibody

Abstract
:1. Introduction
2. Results and Discussion
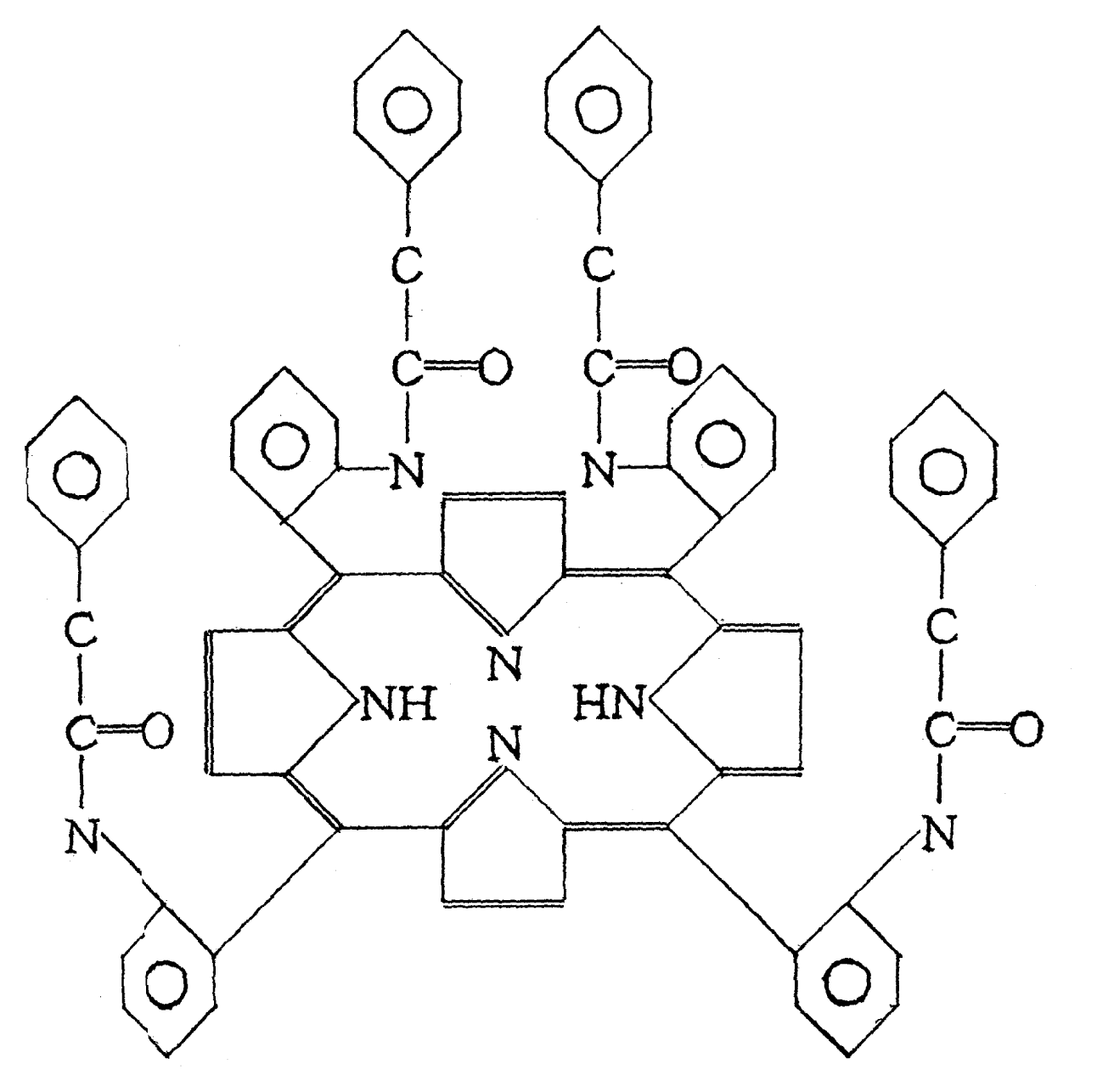
2.1 Preparation of antigen
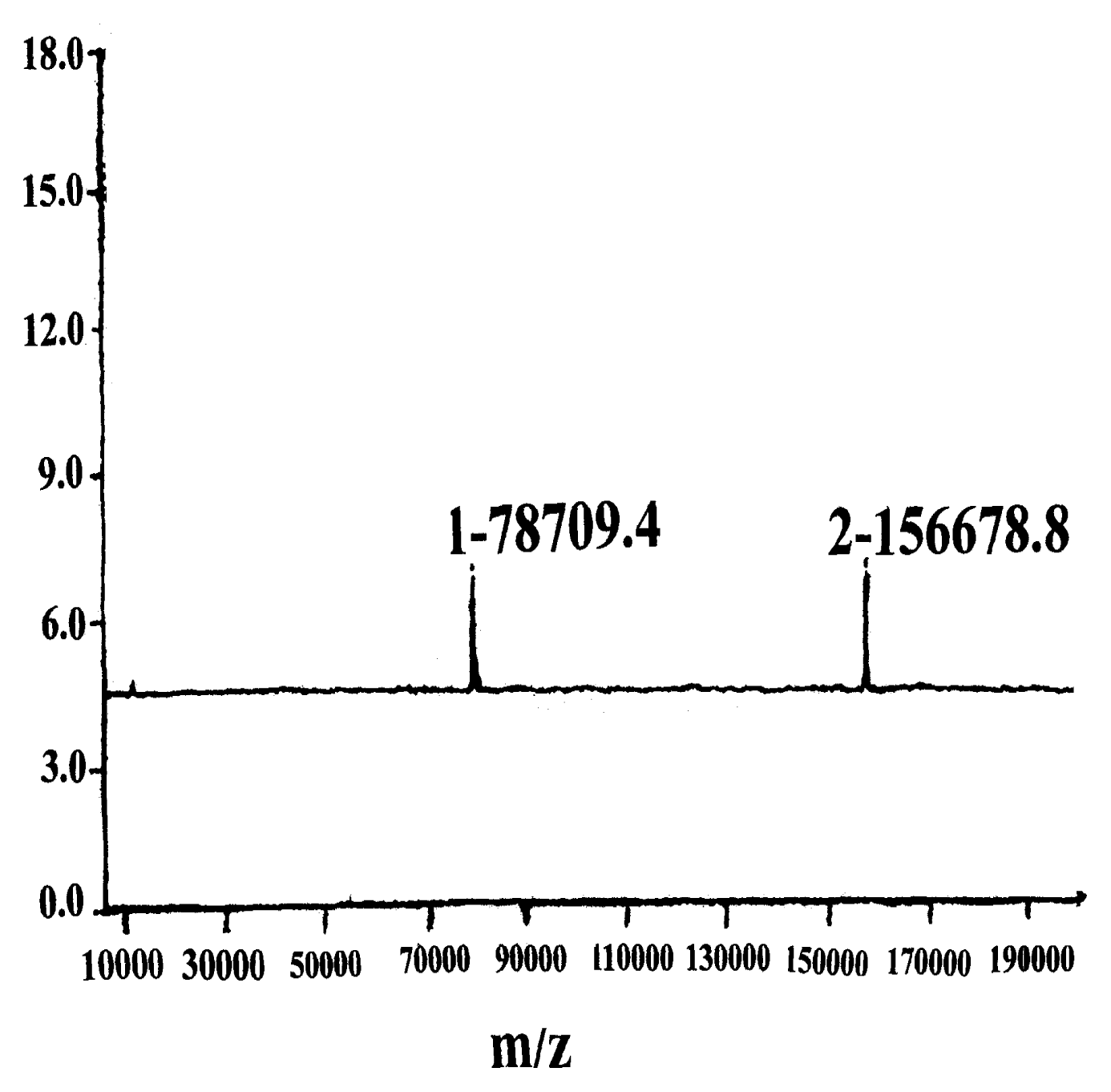
2.2 Building of hybridoma cells
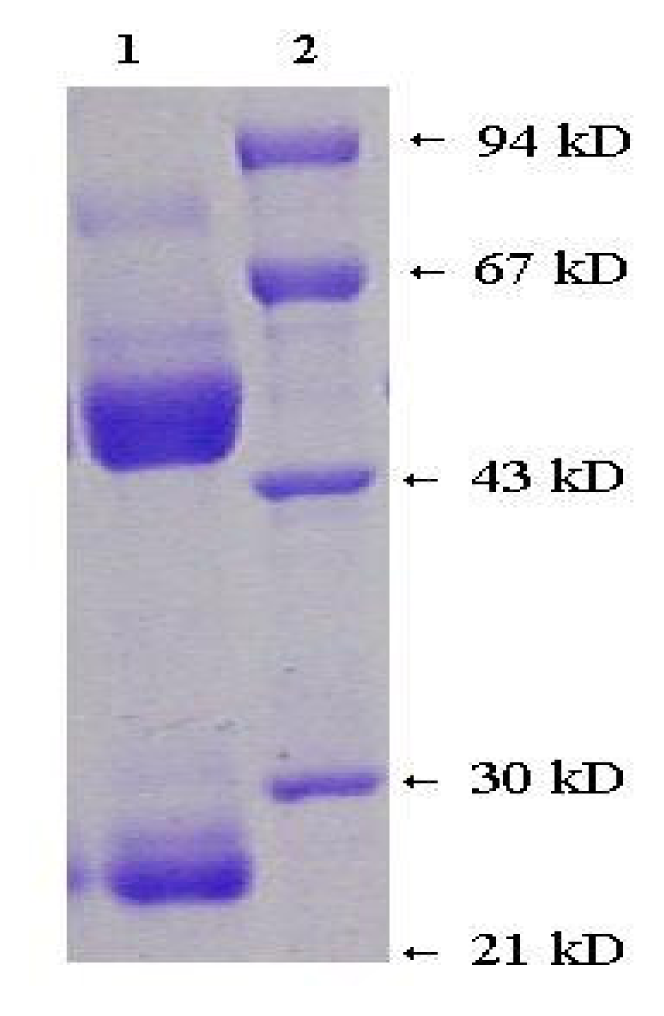
2.3 Antibody properties
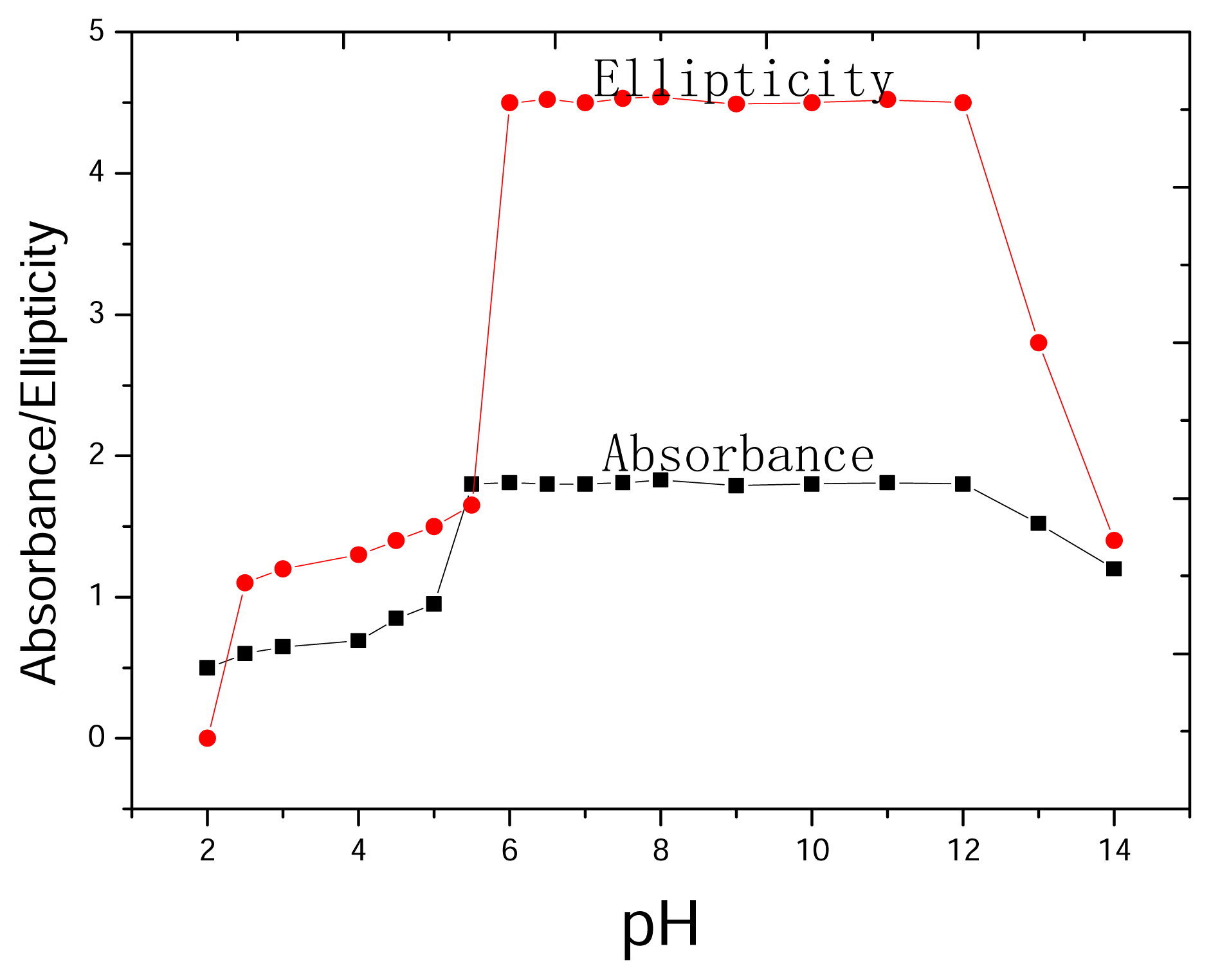
2.4 Dependence on pH
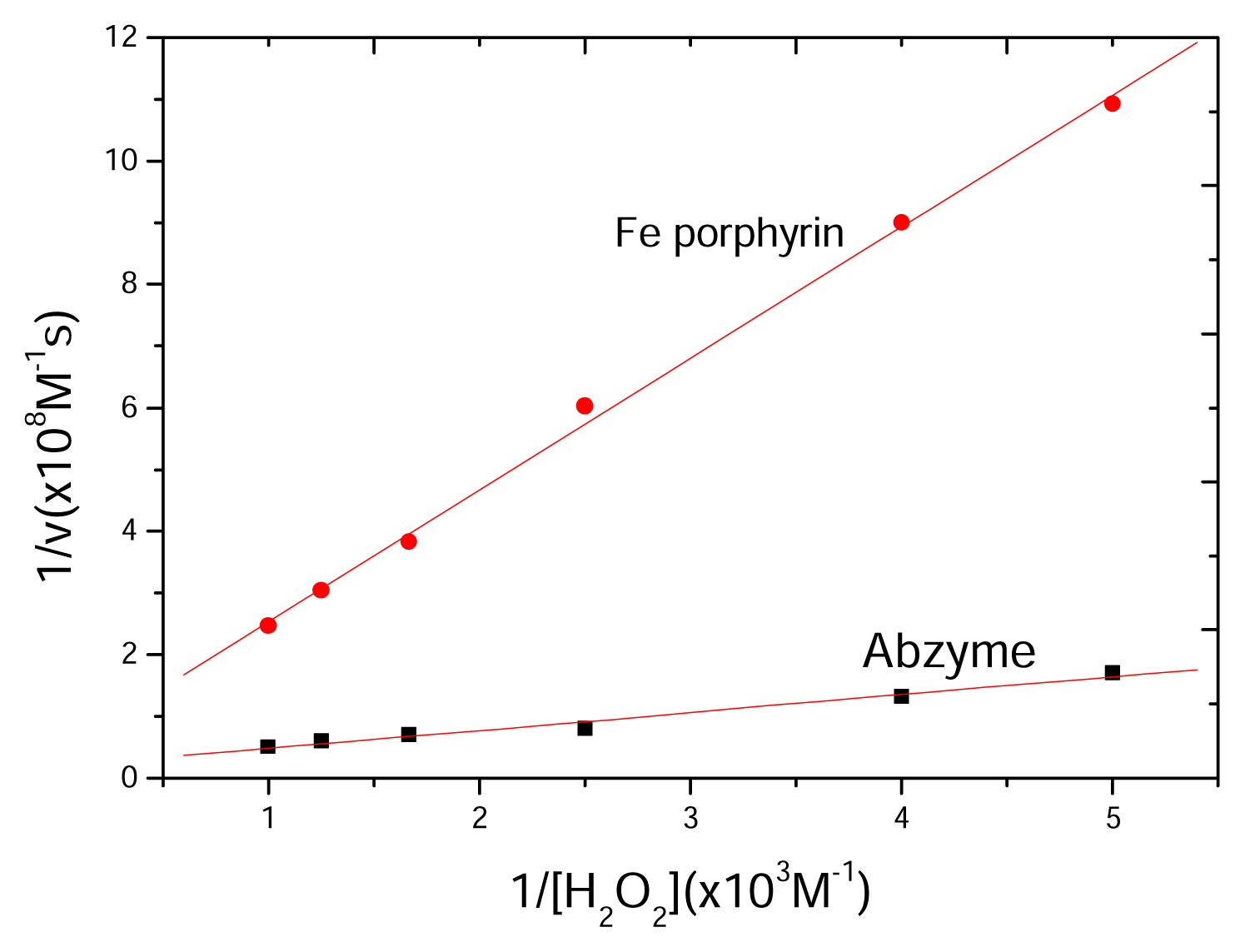
2.5 Kinetics of catalytic reaction
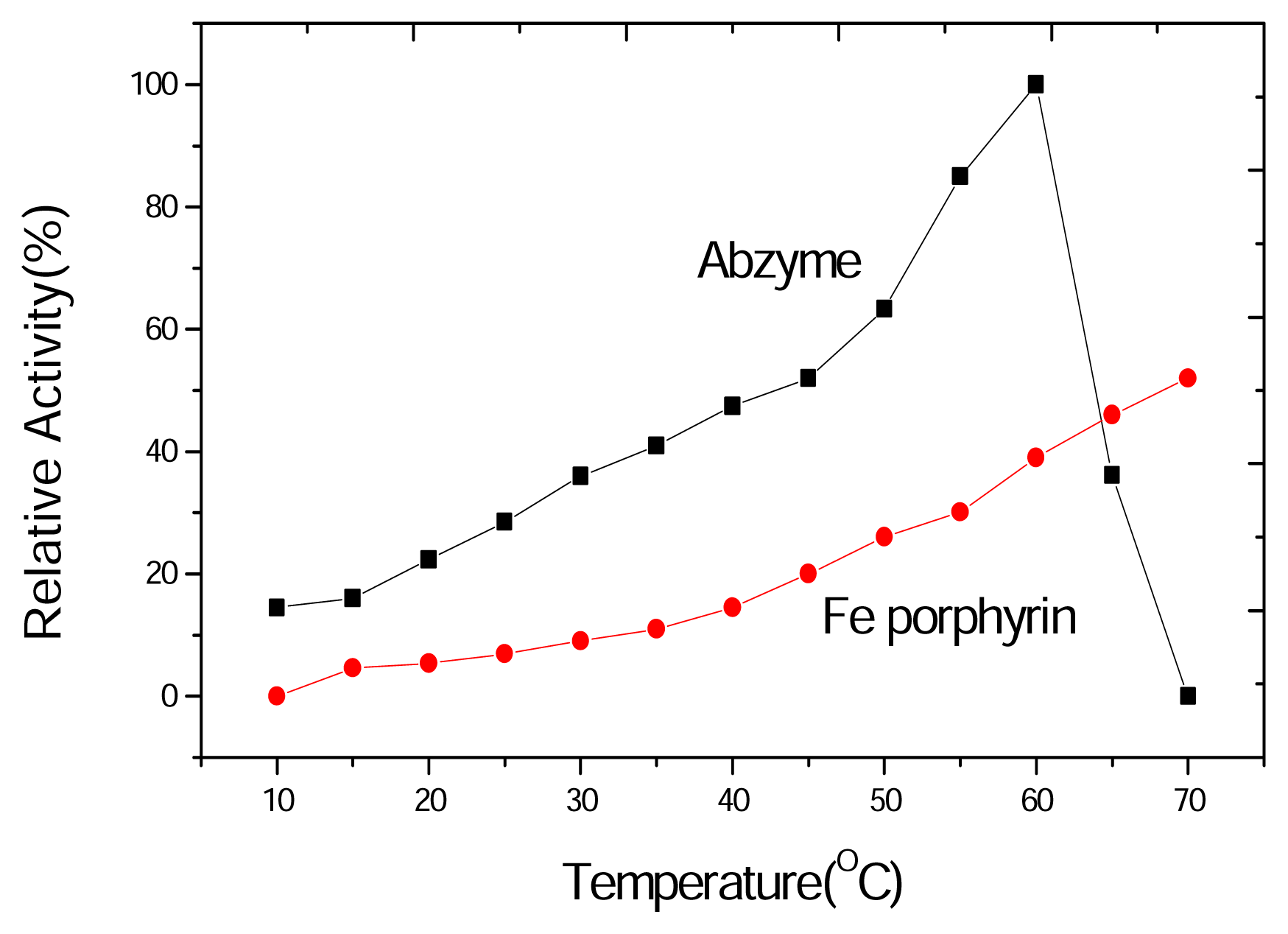
2.6 Thermostability of the chloroperoxidase activity
3. Experimental section
3.1 Materials and Reagents
3.2 Antigen Preparation
3.3 Preparation and purification of monoclonal antibody against meso-tetra(α,α,α,α-o-phenylacetamidophenyl) porphyrin
3.4 Preparation of Fe-Porphyrin
3.5 Dependence on pH
3.6 Assay of chloroperoxidase activity
3.7 Thermostability of chloroperoxidase







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vmax (M·s−1) | Km (M) | κcat (s−1) | κcat/Km (M−1·s−1) | |
|---|---|---|---|---|
| Abzyme | 5.18±0.12×10−8 | 1.50±0.03×10−8 | 0.518±0.1 | 3.45±0.15×107 |
| Fe porphyrin | 2.44±0.05×10−8 | 5.2±0.14×10−8 | 0.244±0.08 | 4.69±0.18×107 |
Acknowledgements
References
- Taurog, A.; Potter, G.D.; Chaikoff, I.L. Conversion of inorganic I131 to organic I131 by cell-free preparation of thyroid tissue. J. Biol. Chem 1955, 213, 119–134. [Google Scholar]
- DeGroot, L.J.; Carvalho, E. Iodide binding in thyroid cellar fractions. J. Biol. Chem 1960, 235, 1390–1397. [Google Scholar]
- Alexander, N.M. Iodide peroxides in rat thyroid and salivary glands and its inhibition by antithyroid compounds. J. Biol. Chem 1959, 234, 1530–1533. [Google Scholar]
- Morris, D.R.; Hager, L.P. Chloroperoxidase I. Isolation and properties of the crystalline glycoprotein. J. Biol. Chem 1966, 241, 1763–1768. [Google Scholar]
- Taurog, A.; Howells, E.M. Enzymatic iodination of tyrosine and thymoglobulin with chloroperoxidase. J. Biol. Chem 1966, 243, 1329–1339. [Google Scholar]
- Lerner, R.A.; Benkovic, S.J.; Schultz, P.G. At the crossroads of chemistry and immunology: catalytic antibodies. Science 1991, 252, 659–667. [Google Scholar]
- Patten, P.A.; Gray, N.S.; Yang, P.L.; Marks, C.B.; Wedemnayer, G.J.; Boniface, J.J.; Stevens, R.C.; Schultz, P.G. The immunological evolution of catalysis. Science 1996, 271, 1086–1091. [Google Scholar]
- Wentworth, P., Jr; Janda, K.D. Catalytic antibodies: structure and function. Cell. Biochem. Biophys 2001, 35, 63–87. [Google Scholar]
- Schultz, P.G. Bring biological solutions to chemical problems. Proc. Natl. Acad. Sci. USA 1998, 95, 14590–14591. [Google Scholar]
- Qi, C.; Ding, L.; Liu, L.Y.; Zhang, Y.J.; Zhao, D.Q.; Ni, J.Z. Antigen synthesis, characterization of artificial haloperoxidase. Chin. J. Appl. Chem 2000, 17, 533–535. [Google Scholar]
- Collman, J.P.; Gagne, R.R.; Reed, C.A.; Halbert, T.R.; Lang, G.; Robinson, W.T. “Picket fence porphyrins” synthetic models for oxygen binding hemoproteins. J. Am. Chem. Soc 1975, 97, 1427–1439. [Google Scholar]
- Thomas, N.R. Hapten design for the generation of catalytic antibodies. Appl. Biochem. Biotech 1994, 47, 345–372. [Google Scholar]
- Kohler, G.; Milstein, C. Continuous cultures of fused cells secreting antibody of predefined specificity. Nature 1975, 256, 495–497. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem 1951, 193, 265–275. [Google Scholar]
- Sun, M.Z.; Ding, L.; Qi, C.; Zhao, D.Q.; Ni, J.Z. Application of MALDI/TOF MS on quality of fibrinolytic enzyme and phospholipids A2 from Chinese Agkistrodon blomhoffii Ussrensis Venom. Chin. J. Pharm. Anal 1999, 19, 367–370. [Google Scholar]
- Jolley, M.E.; Glaudemans, C.P. The determination of binding constants for binding between carbohydrate ligands and certain proteins. Carbohydr. Res 1974, 33, 377–382. [Google Scholar]
- Hallenberg, P.F.; Hager, L.P. Purification of chloroperoxidase from Caldariomyces fumago. Meth. Enzymol 1978, 52, 521–529. [Google Scholar]
© 2007 by MDPI Reproduction is permitted for noncommercial purposes.
Share and Cite
Wang, F.; Huang, X.; Du, L.; Li, W.; He, H.; Qi, C. Preparation and Characterization of a Chloroperoxidase-like Catalytic Antibody. Int. J. Mol. Sci. 2007, 8, 445-454. https://doi.org/10.3390/i8050445
Wang F, Huang X, Du L, Li W, He H, Qi C. Preparation and Characterization of a Chloroperoxidase-like Catalytic Antibody. International Journal of Molecular Sciences. 2007; 8(5):445-454. https://doi.org/10.3390/i8050445
Chicago/Turabian StyleWang, Fengyang, Xueying Huang, Li Du, Weiguo Li, Hongxuan He, and Chao Qi. 2007. "Preparation and Characterization of a Chloroperoxidase-like Catalytic Antibody" International Journal of Molecular Sciences 8, no. 5: 445-454. https://doi.org/10.3390/i8050445