Amperometric Microsensors Monitoring Glutamate-Evoked In Situ Responses of Nitric Oxide and Carbon Monoxide from Live Human Neuroblastoma Cells

Abstract
:
{kind=link}
{kind=link}
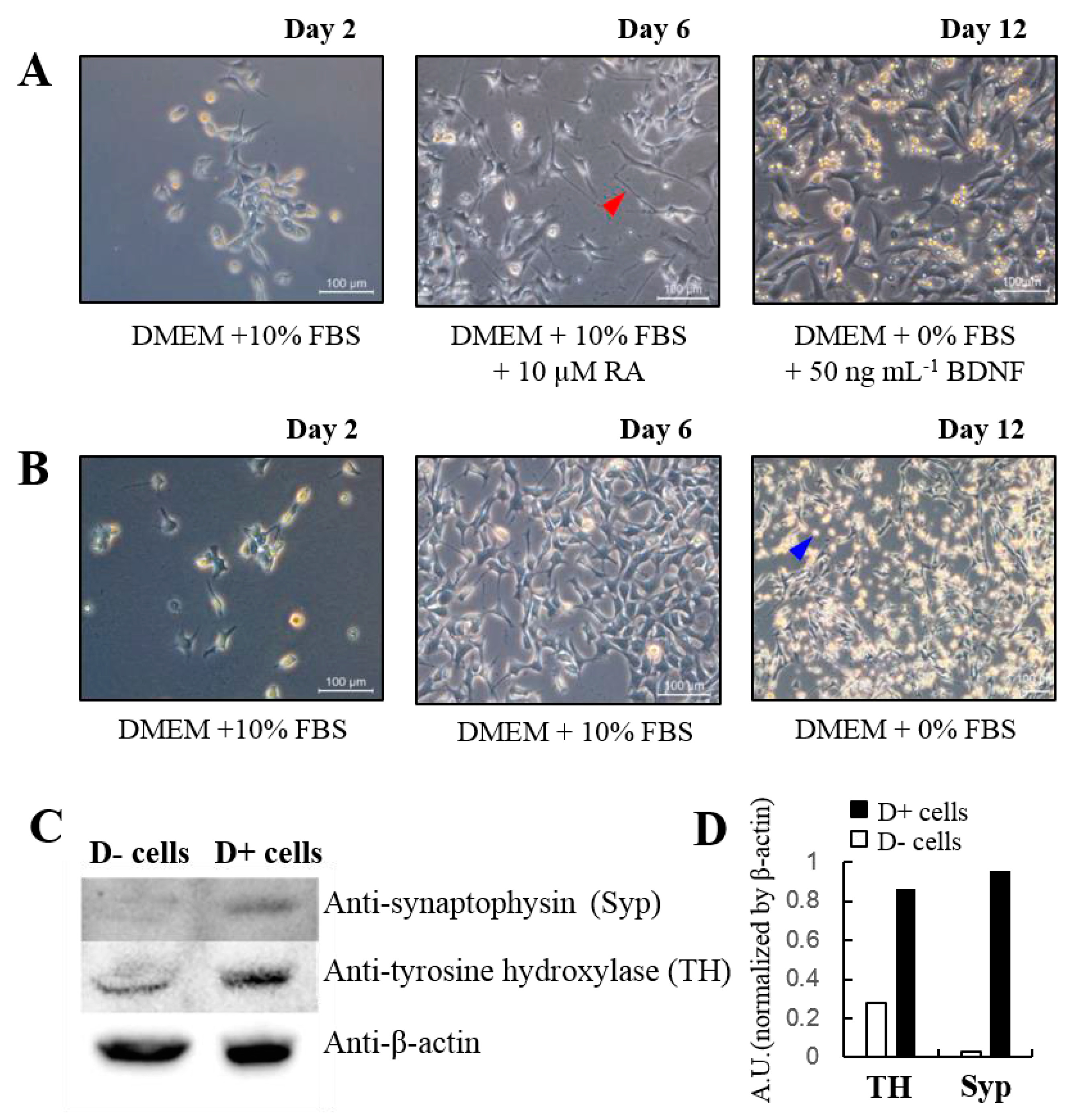{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Preparations of Dual and Septuple NO/CO Sensors
2.2. Cell Culture and Differentiation of SH-SY5Y Cell Line
2.3. NO and CO In Situ Measurements on Neuronal Cells
3. Results and Discussion
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Calabrese, V.; Mancuso, C.; Calvani, M.; Rizzarelli, E.; Butterfield, D.A.; Giuffrida Stella, A.M. Nitric oxide in the central nervous system: Neuroprotection versus neurotoxicity. Nat. Rev. Neurosci. 2007, 8, 766–775. [Google Scholar] [CrossRef] [PubMed]
- Tricoire, L.; Tania, V. Neuronal nitric oxide synthase expressing neurons: A journey from birth to neuronal circuits. Front. Neural Circuits 2012, 6. [Google Scholar] [CrossRef] [PubMed]
- Maines, M.D. The heme oxygenase system: A regulator of second messenger gases. Annu. Rev. Pharmacol. Toxicol. 1997, 37. [Google Scholar] [CrossRef] [PubMed]
- Bidmon, H.-J.; Emde, B.; Oermann, E.; Kubitz, R.; Witte, O.W.; Zilles, K. Heme oxygenase-1 (HSP-32) and heme oxygenase-2 induction in neurons and glial cells of cerebral regions and its relation to iron accumulation after focal cortical photothrombosis. Exp. Neurol. 2001, 16, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Heneka, M.T.; Feinstein, D.L. Expression and function of inducible nitric oxide synthase in neurons. J. Neuroimmunol. 2001, 114, 8–18. [Google Scholar] [CrossRef]
- Cabell, L.; Ferguson, C.; Luginbill, D.; Kern, M.; Weingart, A.; Audesirk, G. Differential induction of heme oxygenase and other stress proteins in cultured hippocampal astrocytes and neurons by inorganic lead. Toxicol. Appl. Pharmacol. 2004, 198, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Rogers, B.; Yakopson, V.; Teng, Z.-P.; Guo, Y.; Regan, R.F. Heme oxygenase-2 knockout neurons are less vulnerable to hemoglobin toxicity. Free Radic. Biol. Med. 2003, 35, 872–881. [Google Scholar] [CrossRef]
- Rössler, O.G.; Bauer, I.; Chung, H.-Y.; Thiel, G. Glutamate-induced cell death of immortalized murine hippocampal neurons: Neuroprotective activity of heme oxygenase-1, heat shock protein 70, and sodium selenite. Neurosci. Lett. 2004, 362, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Khandare, A.L.; Ankulu, M.; Aparna, N. Role of glutamate and nitric oxide in onset of motor neuron degeneration in neurolathyrism. Neurotoxicology 2013, 34, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Viadel, M.; Montoliu, C.; Monfort, P.; Canales, J.J.; Erceg, S.; Rowan, M.; Ceccatelli, S.; Felipo, V. Chronic exposure to 2,5-hexanedione impairs the glutamate-nitric oxide-cyclic GMP pathway in cerebellar neurons in culture and in rat brain in vivo. Neurochem. Int. 2003, 42, 525–533. [Google Scholar] [CrossRef]
- Attwell, D.; Buchan, A.M.; Charpak, S.; Lauritzen, M.; MacVicar, B.A.; Newman, E.A. Glial and neuronal control of brain blood flow. Nature 2010, 468, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Llansola, M.; Montoliu, C.; Boix, J.; Felipo, V. Polychlorinated biphenyls PCB 52, PCB 180, and PCB 138 impair the glutamate−nitric oxide−cGMP pathway in cerebellar neurons in culture by different mechanisms. Chem. Res. Toxicol. 2010, 23, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.-H.; Kim, H.J.; Jeong, I.; Hong, Y.J.; Kim, M.-J.; Rhie, D.-J.; Jo, Y.-H.; Hahn, S.J.; Yoon, S.H. Grape seed proanthocyanidin extract inhibits glutamate-induced cell death through inhibition of calcium signals and nitric oxide formation in cultured rat hippocampal neurons. BMC Neurosci. 2011, 12, 78. [Google Scholar] [CrossRef] [PubMed]
- Mannella, P.; Sanchez, A.M.; Giretti, M.S.; Genazzani, A.R.; Simoncini, T. Oestrogen and progestins differently prevent glutamate toxicity in cortical neurons depending on prior hormonal exposure via the induction of neural nitric oxide synthase. Steroids 2009, 74, 650–656. [Google Scholar] [CrossRef] [PubMed]
- Quesada, A.; Ogi, J.; Schultz, J.; Handforth, A. C-terminal mechano-growth factor induces heme oxygenase-1-mediated neuroprotection of SH-SY5Y cells via the protein kinase Cϵ/Nrf2 pathway. J. Neurosci. Res. 2011, 89, 394–405. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Wang, R. Carbon monoxide: Endogenous production, physiological functions, and pharmacological applications. Pharmacol. Rev. 2005, 57, 585–630. [Google Scholar] [CrossRef] [PubMed]
- Knowles, R.G.; Moncada, S. Nitric oxide synthases in mammals. Biochem. J. 1994, 298, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Liu, Q.; Han, H.-C. Buckling reduces eNOS production and stimulates extracellular matrix remodeling in arteries in organ culture. Ann. Biomed. Eng. 2016, 44, 2840–2850. [Google Scholar] [CrossRef] [PubMed]
- Palipoch, S.; Koomhin, P.; Punsawad, C.; Na-Ek, P.; Sattayakhom, A.; Suwannalert, P. Heme oxygenase-1 alleviates alcoholic liver steatosis: Histopathological study. J. Toxicol. Pathol. 2016, 29, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Retterstol, L.; Lyberg, T.; Aspelin, T.; Berg, K. A twin study of nitric oxide levels measured by serum nitrite/nitrate. Twin Res. Hum. Genet. 2006, 9, 210–214. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Yang, G.; Li, H.; Du, D.; Lin, Y. Electrochemical sensors and biosensors based on nanomaterials and nanostructures. Anal. Chem. 2015, 87, 230–249. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Scafa, N.; Xu, L.P.; Zhou, S.; Abdullah Al-Ghanem, K.; Mahboob, S.; Fugetsu, B.; Zhang, X. Electrochemical hydrogen sulfide biosensors. Analyst 2016, 141. [Google Scholar] [CrossRef] [PubMed]
- Govindhan, M.; Liu, Z.; Chen, A. Design and electrochemical study of platinum-based nanomaterials for sensitive detection of nitric oxide in biomedical applications. Nanomaterials 2016, 6, 211. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Scafa, N.; Xu, L.-P.; Su, L.; Li, C.; Zhou, S.; Liu, Y.; Zhang, X. Electrochemical sensors for nitric oxide detection in biological applications. Electroanalysis 2014, 26, 449–468. [Google Scholar] [CrossRef]
- Dang, X.; Hu, H.; Wang, S.; Hu, S. Nanomaterials-based electrochemical sensors for nitric oxide. Microchim. Acta 2015, 182, 455–467. [Google Scholar] [CrossRef]
- Krediet, T.G.; Cirkel, G.A.; Vreman, H.J.; Wong, R.J.; Stevenson, D.K.; Groenendaal, F.; Egberts, J.; Van Bel, F. End-tidal carbon monoxide measurements in infant respiratory distress syndrome. Acta Paediatr. 2006, 95, 1075–1082. [Google Scholar] [CrossRef] [PubMed]
- Obermeier, J.; Trefz, P.; Wex, K.; Sabel, B.; Schubert, J.K.; Miekisch, W. Electrochemical sensor system for breath analysis of aldehydes, CO and NO. J. Breath Res. 2015, 9, 016008. [Google Scholar] [CrossRef] [PubMed]
- Park, S.S.; Kim, J.; Lee, Y. Improved electrochemical microsensor for the real-time simultaneous analysis of endogenous nitric oxide and carbon monoxide generation. Anal. Chem. 2012, 84. [Google Scholar] [CrossRef] [PubMed]
- Park, S.S.; Hong, M.; Ha, Y.; Sim, J.; Jhon, G.J.; Lee, Y.; Suh, M. The real-time in vivo electrochemical measurement of nitric oxide and carbon monoxide release upon direct epidural electrical stimulation of the rat neocortex. Analyst 2015, 140. [Google Scholar] [CrossRef] [PubMed]
- Ha, Y.; Sim, J.; Lee, Y.; Suh, M. Insertable fast-response amperometric NO/CO dual microsensor: Study of neurovascular coupling during acutely induced seizures of rat brain cortex. Anal. Chem. 2016, 88, 2563–2569. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.-R.; Hu, L.-S.; Li, G.-Y. SH-SY5Y human neuroblastoma cell line: In vitro cell model of dopaminergic neurons in Parkinson’s disease. Chin. Med. J. 2010, 123, 1086–1092. [Google Scholar] [PubMed]
- Encinas, M.; Iglesias, M.; Liu, Y.; Wang, H.; Muhaisen, A.; Ceña, V.; Gallego, C.; Comella, J.X. Sequential treatment of SH-SY5Y cells with retinoic acid and brain-derived neurotrophic factor gives rise to fully differentiated, neurotrophic factor-dependent, human neuron-like cells. J. Neurochem. 2000, 75, 991–1003. [Google Scholar] [CrossRef] [PubMed]
- Kaewsuk, S.; Sae-ung, K.; Phansuwan-Pujito, P.; Govitrapong, P. Melatonin attenuates methamphetamine-induced reduction of tyrosine hydroxylase, synaptophysin and growth-associated protein-43 levels in the neonatal rat brain. Neurochem. Int. 2009, 55, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Wise, D.L.; Houghton, G. Diffusion coefficients of neon, krypton, xenon, carbon monoxide and nitric oxide in water at 10–60 °C. Chem. Eng. Sci. 1968, 23, 1211–1216. [Google Scholar] [CrossRef]




© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ha, Y.; Heo, C.; Woo, J.; Ryu, H.; Lee, Y.; Suh, A.M. Amperometric Microsensors Monitoring Glutamate-Evoked In Situ Responses of Nitric Oxide and Carbon Monoxide from Live Human Neuroblastoma Cells. Sensors 2017, 17, 1661. https://doi.org/10.3390/s17071661
Ha Y, Heo C, Woo J, Ryu H, Lee Y, Suh AM. Amperometric Microsensors Monitoring Glutamate-Evoked In Situ Responses of Nitric Oxide and Carbon Monoxide from Live Human Neuroblastoma Cells. Sensors. 2017; 17(7):1661. https://doi.org/10.3390/s17071661
Chicago/Turabian StyleHa, Yejin, Chaejeong Heo, Juhyun Woo, Hyunwoo Ryu, Youngmi Lee, and And Minah Suh. 2017. "Amperometric Microsensors Monitoring Glutamate-Evoked In Situ Responses of Nitric Oxide and Carbon Monoxide from Live Human Neuroblastoma Cells" Sensors 17, no. 7: 1661. https://doi.org/10.3390/s17071661