
Development of a Smart Wireless Multisensor Platform for an Optogenetic Brain Implant
 , , , , , , , , , , , , , and
, , , , , , , , , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Overview
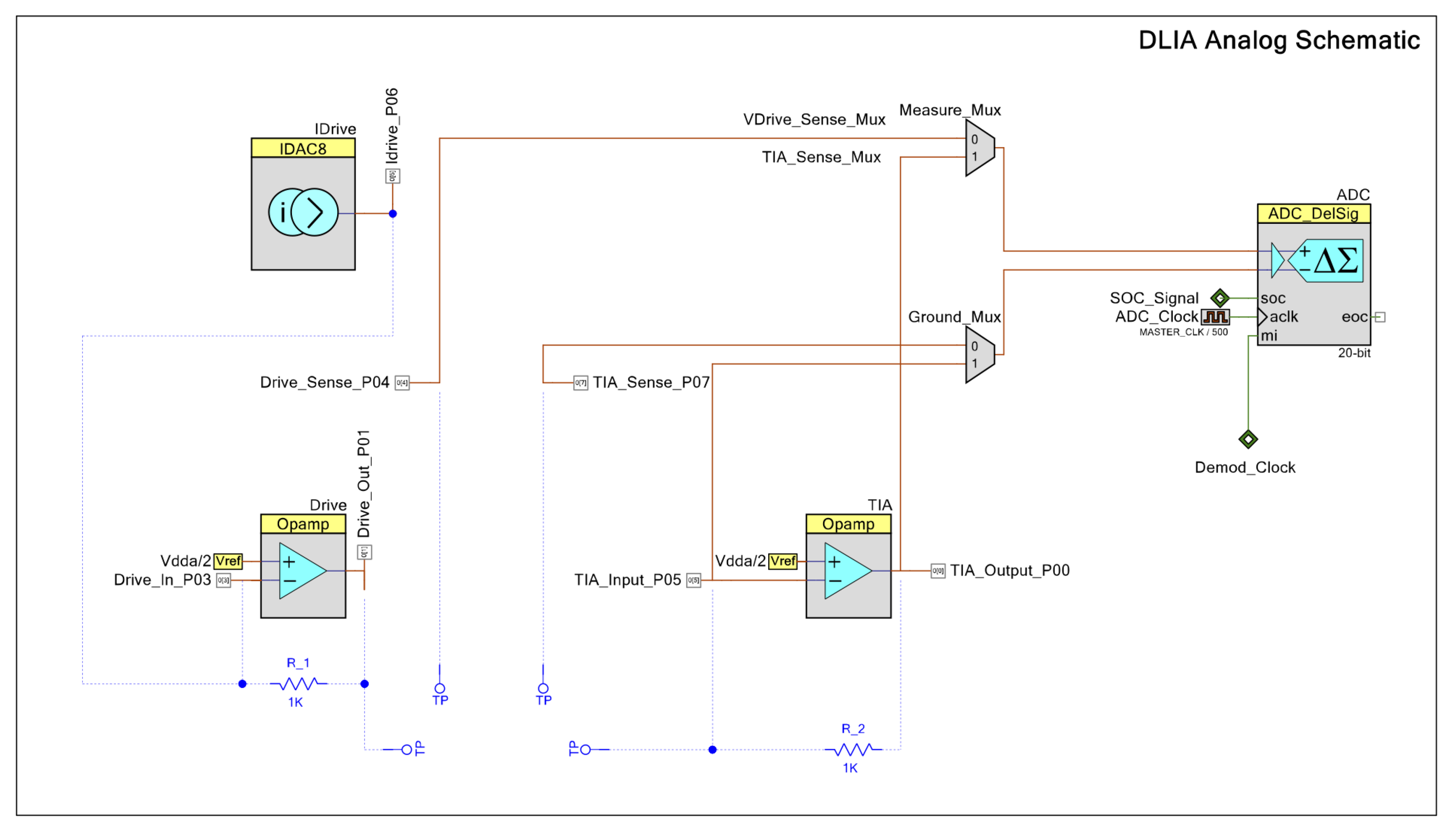
2.2. Electronic Instrumentation
2.2.1. Optical Stimulator
2.2.2. Impedance Analyzer
2.2.3. Potentiostat
2.2.4. Communications and Control
2.3. System under Test (SUT)
2.3.1. Cell Culture
2.3.2. Neuronal Differentiation
2.3.3. Immunocytochemistry
2.3.4. Electrodes and In Vitro System
2.3.5. Amperometric Detection of Neurotransmitter Release
3. Results
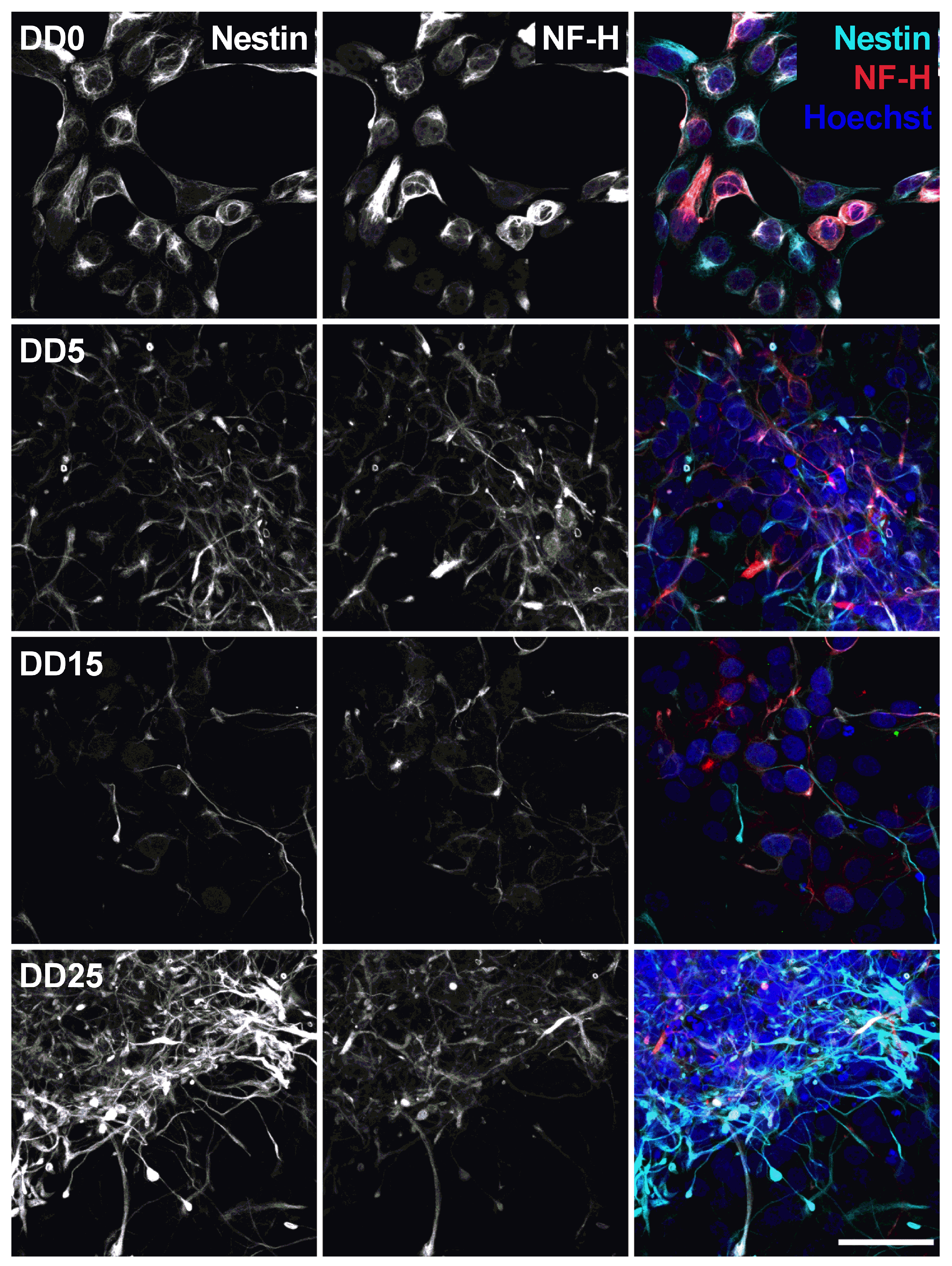
3.1. Neural Progenitor Cell Growth and Differentiation
3.2. Performance of the Miniaturized Impedance Analyzer Is Comparable to a Desktop Instrument
3.3. Potentiostat
3.3.1. The Miniaturized Headstage Potentiostat Is Able to Detect Dopamine Release from Neurons upon Depolarization by Increasing the Extracellular Potassium Concentration
3.3.2. The Miniaturized Potentiostat Can Also Detect Dopamine Release from Optogenetically Modified Neurons upon Optical Stimulation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AC | alternating current |
| ADC | analog-to-digital converter |
| BLE | Bluetooth low energy |
| BM | basic medium |
| CE | counter electrode |
| ChR2 | channelrhodopsin-2 |
| CRM | cell-based regenerative medicine |
| CV | cyclic voltammetry |
| DAC | digital-to-analog converter |
| DBS | deep brain stimulation |
| DC | direct current |
| DD | differentiation day |
| DM | differentiation medium |
| FBS | fetal bovine serum |
| FPGA | field-programmable gate array |
| GFAP | glial fibrillary acidic protein |
| GM | growth medium |
| LD | laser diode |
| LIA | lock-in amplifier |
| MAP-2 | microtubule-associated protein 2 |
| ML | machine learning |
| ND | neutral density |
| NeuN | neuronal nuclear antigen |
| NF-H | neurofilament heavy |
| NPC | neural progenitor cell |
| TH | tyrosine hydroxylase |
| PBS | phosphate-buffered saline |
| PCB | printed circuit board |
| PD | Parkinson’s disease |
| PSoC™ | programmable system on chip |
| RE | reference electrode |
| SNR | signal-to-noise ratio |
| SUT | sample/system under test |
| TIA | transimpedance amplifier |
| WE | working electrode |
Appendix A. Additional Figures
Appendix A.1. Electric Circuit Boards



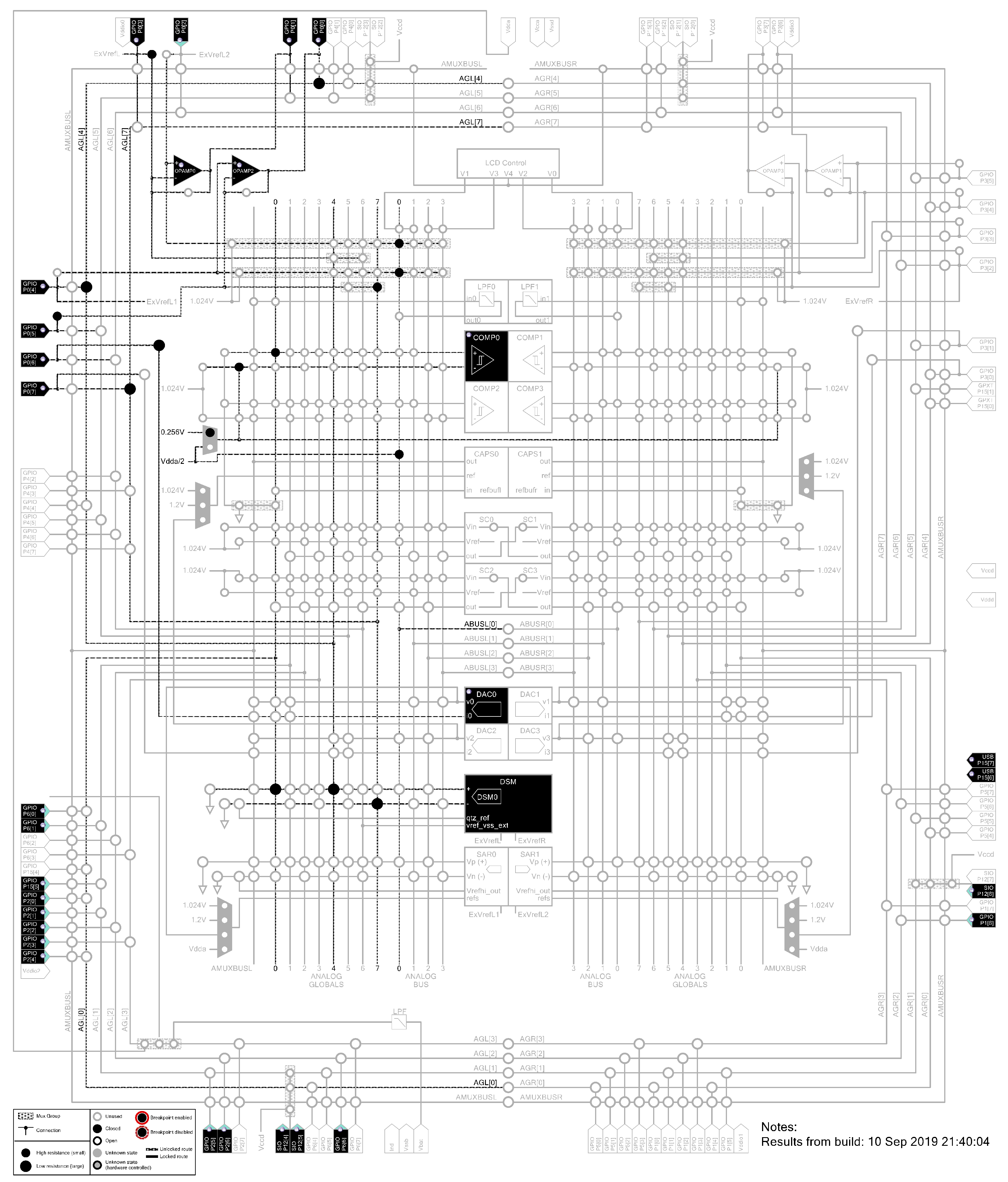
Appendix A.2. Schematics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Input Hex | in1 | in0 | out2 | out1 | out0 | Output Hex |
|---|---|---|---|---|---|---|
| 00 | 0 | 0 | 0 | 0 | 1 | 0x01 |
| 01 | 0 | 1 | 0 | 1 | 0 | 0x02 |
| 02 | 1 | 0 | 1 | 1 | 1 | 0x07 |
| 03 | 1 | 1 | 1 | 0 | 0 | 0x04 |
| Input Hex | in4 | in3 | in2 | in1 | in0 | out2 | out1 | out0 | Output Hex |
|---|---|---|---|---|---|---|---|---|---|
| 00 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0x00 |
| 01 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0x02 |
| 02 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0x00 |
| 03 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 0x01 |
| 04 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0x02 |
| 05 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 0x02 |
| 06 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 0x01 |
| 07 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 0x01 |
| 08 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0x02 |
| 09 | 0 | 1 | 0 | 0 | 1 | 0 | 1 | 0 | 0x02 |
| 0A | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 0x04 |
| 0B | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 0x04 |
| 0C | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0x00 |
| 0D | 0 | 1 | 1 | 0 | 1 | 0 | 1 | 1 | 0x03 |
| 0E | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0x00 |
| 0F | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 0x03 |
| 10 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0x05 |
| 11 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0x05 |
| 12 | 1 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 0x05 |
| 13 | 1 | 0 | 0 | 1 | 1 | 1 | 0 | 1 | 0x05 |
| 14 | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 0x06 |
| 15 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 0x06 |
| 16 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 0x06 |
| 17 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 0x06 |
| 18 | 1 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 0x07 |
| 19 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 0x07 |
| 1A | 1 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 0x07 |
| 1B | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 0x07 |
| 1C | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 1 | 0x03 |
| 1D | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 1 | 0x03 |
| 1E | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 0x03 |
| 1F | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 0x03 |

Appendix A.3. Electrode Chips, Cell Culture Chip Holder and Impedance Performance Measurements

Appendix A.4. Electrochemistry Performance Measurements







References
- de Lau, L.M.L.; Breteler, M.M.B. Epidemiology of Parkinson’s disease. Lancet Neurol. 2006, 5, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Dorsey, E.R.; Elbaz, A.; Nichols, E.; Abbasi, N.; Abd-Allah, F.; Abdelalim, A.; Adsuar, J.C.; Ansha, M.G.; Brayne, C.; Choi, J.Y.J.; et al. Global, regional, and national burden of Parkinson’s disease, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2018, 17, 939–953. [Google Scholar] [CrossRef]
- Kalia, L.V.; Lang, A.E. Parkinson’s disease. Lancet 2015, 386, 896–912. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, K.R.; Schapira, A.H.V. Non-motor symptoms of Parkinson’s disease: Dopaminergic pathophysiology and treatment. Lancet Neurol. 2006, 5, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Barker, R.A.; Parmar, M.; Studer, L.; Takahashi, J. Human trials of stem cell-derived dopamine neurons for Parkinson’s disease: Dawn of a new era. Cell Stem Cell 2017, 21, 569–573. [Google Scholar] [CrossRef]
- Ganz, J.; Lev, N.; Melamed, E.; Offen, D. Cell replacement therapy for Parkinson’s disease: How close are we to the clinic? Expert Rev. Neurother. 2011, 11, 1325–1339. [Google Scholar] [CrossRef] [PubMed]
- Mendez, I.; Viñuela, A.; Astradsson, A.; Mukhida, K.; Hallett, P.; Robertson, H.; Tierney, T.; Holness, R.; Dagher, A.; Klassen, H. Dopamine neurons implanted into people with Parkinson’s disease survive without pathology for 14 years. Nat. Med. 2008, 14, 507–509. [Google Scholar] [CrossRef]
- Politis, M.; Lindvall, O. Clinical application of stem cell therapy in Parkinson’s disease. BMC Med. 2012, 10, 1. [Google Scholar] [CrossRef]
- Kirkeby, A.; Nelander, J.; Hoban, D.B.; Rogelius, N.; Bjartmarz, H.; Storm, P.; Fiorenzano, A.; Adler, A.F.; Vale, S.; Mudannayake, J.; et al. Preclinical quality, safety, and efficacy of a human embryonic stem cell-derived product for the treatment of Parkinson’s disease, STEM-PD. Cell Stem Cell 2023, 30, 1299–1314. [Google Scholar] [CrossRef]
- Kordower, J.H.; Sortwell, C.E. Neuropathology of fetal nigra transplants for Parkinson’s disease. Prog. Brain Res. 2000, 127, 333–344. [Google Scholar]
- Wichmann, T.; DeLong, M.R. Deep brain stimulation for neurologic and neuropsychiatric disorders. Neuron 2006, 52, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Okun, M.S. Deep-brain stimulation for Parkinson’s disease. N. Engl. J. Med. 2012, 367, 1529–1538. [Google Scholar] [CrossRef] [PubMed]
- Deisseroth, K. Optogenetics. Nat. Methods 2011, 8, 26–29. [Google Scholar] [CrossRef] [PubMed]
- Park, H.E.; Kim, D.; Koh, H.S.; Cho, S.; Sung, J.S.; Kim, J.Y. Real-time monitoring of neural differentiation of human mesenchymal stem cells by electric cell-substrate impedance sensing. J. Biomed. Biotechnol. 2011, 2011, 485173. [Google Scholar] [CrossRef]
- Gamal, W.; Wu, H.; Underwood, I.; Jia, J.; Smith, S.; Bagnaninchi, P.O. Impedance-based cellular assays for regenerative medicine. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2018, 373, 20170226. [Google Scholar] [CrossRef]
- Asif, A.; Heiskanen, A.; Emnéus, J.; Keller, S.S. Pyrolytic carbon nanograss electrodes for electrochemical detection of dopamine. Electrochim. Acta 2021, 379, 138122. [Google Scholar] [CrossRef]
- Robinson, D.L.; Hermans, A.; Seipel, A.T.; Wightman, R.M. Monitoring rapid chemical communication in the brain. Chem. Rev. 2008, 108, 2554–2584. [Google Scholar] [CrossRef] [PubMed]
- Quinones, C.I.D. Microsystems for Real-Time Neurochemical Monitoring Using Fast-Scan Cyclic Voltammetry; Cornell University: Ithaca, NY, USA, 2014. [Google Scholar]
- Elgrishi, N.; Rountree, K.J.; McCarthy, B.D.; Rountree, E.S.; Eisenhart, T.T.; Dempsey, J.L. A practical beginner’s guide to cyclic voltammetry. J. Chem. Educ. 2018, 95, 197–206. [Google Scholar] [CrossRef]
- Venton, B.J.; Cao, Q. Fundamentals of fast-scan cyclic voltammetry for dopamine detection. Analyst 2020, 145, 1158–1168. [Google Scholar] [CrossRef]
- Wu, F.; Yu, P.; Mao, L. New Opportunities of Electrochemistry for Monitoring, Modulating, and Mimicking the Brain Signals. JACS Au 2023, 3, 2062–2072. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Z.; Zhou, Y.; Tian, Y. Implantable Electrochemical Sensors for Brain Research. JACS Au 2023, 3, 1572–1582. [Google Scholar] [CrossRef] [PubMed]
- Park, J.O.; Choi, Y.; Ahn, H.M.; Lee, C.K.; Chun, H.; Park, Y.M.; Kim, K.B. Aggregation of Ag nanoparticle based on surface acoustic wave for surface-enhanced Raman spectroscopy detection of dopamine. Anal. Chim. Acta 2024, 1285, 342036. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekar, N.; Balaji, R.; Perala, R.S.; Nik Humaidi, N.Z.; Shanmugam, K.; Liao, Y.C.; Hwang, M.T.; Govindaraju, S. A brief review of graphene-based biosensors developed for rapid detection of COVID-19 biomarkers. Biosensors 2023, 13, 307. [Google Scholar] [CrossRef] [PubMed]
- Gokulkumar, K.; Huang, S.J.; Wang, S.F.; Balaji, R.; Chandrasekar, N.; Hwang, M.T. Zinc molybdate/functionalized carbon nanofiber composites modified electrodes for high-performance amperometric detection of hazardous drug Sulfadiazine. OpenNano 2023, 10, 100131. [Google Scholar] [CrossRef]
- Ellington, A.D.; Szostak, J.W. In vitro selection of RNA molecules that bind specific ligands. Nature 1990, 346, 818–822. [Google Scholar] [CrossRef]
- Chen, G.; Xu, M.; He, C. Preparation of an aptamer electrochemical sensor for the highly sensitive detection of glioma cells. Int. J. Electrochem. Sci. 2023, 18, 100129. [Google Scholar] [CrossRef]
- Nakatsuka, N.; Yang, K.A.; Abendroth, J.M.; Cheung, K.M.; Xu, X.; Yang, H.; Zhao, C.; Zhu, B.; Rim, Y.S.; Yang, Y.; et al. Aptamer–field-effect transistors overcome Debye length limitations for small-molecule sensing. Science 2018, 362, 319–324. [Google Scholar] [CrossRef]
- Liu, X.; Hou, Y.; Chen, S.; Liu, J. Controlling dopamine binding by the new aptamer for a FRET-based biosensor. Biosens. Bioelectron. 2021, 173, 112798. [Google Scholar] [CrossRef]
- Li, X.; Jin, Y.; Zhu, F.; Liu, R.; Jiang, Y.; Jiang, Y.; Mao, L. Electrochemical Conjugation of Aptamers on a Carbon Fiber Microelectrode Enables Highly Stable and Selective In Vivo Neurosensing. Angew. Chem. 2022, 134, e202208121. [Google Scholar] [CrossRef]
- Hao, R.; Liu, L.; Yuan, J.; Wu, L.; Lei, S. Recent Advances in Field Effect Transistor Biosensors: Designing Strategies and Applications for Sensitive Assay. Biosensors 2023, 13, 426. [Google Scholar] [CrossRef]
- Abrantes, M.; Rodrigues, D.; Domingues, T.; Nemala, S.S.; Monteiro, P.; Borme, J.; Alpuim, P.; Jacinto, L. Ultrasensitive dopamine detection with graphene aptasensor multitransistor arrays. J. Nanobiotechnol. 2022, 20, 495. [Google Scholar] [CrossRef]
- Liu, C.; Zhao, Y.; Cai, X.; Xie, Y.; Wang, T.; Cheng, D.; Li, L.; Li, R.; Deng, Y.; Ding, H.; et al. A wireless, implantable optoelectrochemical probe for optogenetic stimulation and dopamine detection. Microsyst. Nanoeng. 2020, 6, 64. [Google Scholar] [CrossRef] [PubMed]
- Vasudevan, S.; Kajtez, J.; Bunea, A.I.; Gonzalez-Ramos, A.; Ramos-Moreno, T.; Heiskanen, A.; Kokaia, M.; Larsen, N.B.; Martínez-Serrano, A.; Keller, S.S.; et al. Leaky Optoelectrical Fiber for Optogenetic Stimulation and Electrochemical Detection of Dopamine Exocytosis from Human Dopaminergic Neurons. Adv. Sci. 2020, 7, 1902011. [Google Scholar] [CrossRef] [PubMed]
- Vasudevan, S.; Dotti, A.; Kajtez, J.; Martínez-Serrano, A.; Gundlach, C.; Maçãs, S.C.; Lauschke, K.; Vinngaard, A.M.; López, S.G.; Pereira, M.; et al. Omnidirectional leaky opto-electrical fiber for optogenetic control of neurons in cell replacement therapy. Bioelectrochemistry 2023, 149, 108306. [Google Scholar] [CrossRef] [PubMed]
- Da, Y.; Luo, S.; Tian, Y. Real-Time Monitoring of Neurotransmitters in the Brain of Living Animals. ACS Appl. Mater. Interfaces 2023, 15, 138–157. [Google Scholar] [CrossRef] [PubMed]
- Cunha, A.B.; Schuelke, C.; Heiskanen, A.; Asif, A.; Hassan, Y.M.; Keller, S.S.; Kalvøy, H.; Martínez-Serrano, A.; Emnéus, J.; Martinsen, Ø.G. Bioimpedance Measurements on Human Neural Stem Cells as a Benchmark for the Development of Smart Mobile Biomedical Applications. In Proceedings of the International Conference on Electrical Bioimpedance: ICEBI 2019, Joinville, SC, Brazil, 9–14 June 2019; pp. 38–47. [Google Scholar]
- Mesri, A.; Sampietro, M.; Cunha, A.B.; Ferrari, G.; Martinsen, Ø.G. A Laser Diode-Based Wireless Optogenetic Headstage. In Proceedings of the 2018 14th Conference on Ph.D. Research in Microelectronics and Electronics (PRIME), Prague, Czech Republic, 2–5 July 2018; pp. 257–260. [Google Scholar] [CrossRef]
- Chen, J.; Tarkhan, M.; Wu, H.; Noshahr, F.H.; Yang, J.; Sawan, M. Recent trends and future prospects of neural recording circuits and systems: A tutorial brief. IEEE Trans. Circuits Syst. II Express Briefs 2022, 69, 2654–2660. [Google Scholar] [CrossRef]
- Dryden, M.D.; Wheeler, A.R. DStat: A versatile, open-source potentiostat for electroanalysis and integration. PLoS ONE 2015, 10, e0140349. [Google Scholar] [CrossRef] [PubMed]
- Villa, A.; Liste, I.; Courtois, E.T.; Seiz, E.G.; Ramos, M.; Meyer, M.; Juliusson, B.; Kusk, P.; Martinez-Serrano, A. Generation and properties of a new human ventral mesencephalic neural stem cell line. Exp. Cell Res. 2009, 315, 1860–1874. [Google Scholar] [CrossRef]
- Courtois, E.T.; Castillo, C.G.; Seiz, E.G.; Ramos, M.; Bueno, C.; Liste, I.; Martínez-Serrano, A. In vitro and in vivo enhanced generation of human A9 dopamine neurons from neural stem cells by Bcl-XL. J. Biol. Chem. 2010, 285, 9881–9897. [Google Scholar] [CrossRef]
- Hassan, Y.M.; Caviglia, C.; Hemanth, S.; Mackenzie, D.; Alstrøm, T.S.; Petersen, D.H.; Keller, S.S. High temperature SU-8 pyrolysis for fabrication of carbon electrodes. J. Anal. Appl. Pyrolysis 2017, 125, 91–99. [Google Scholar] [CrossRef]
- Savitzky, A.; Golay, M. Smoothing and Differentiation of Data by Simplified Least Squares Procedures. Anal. Chem. 1964, 36, 1627–1639. [Google Scholar] [CrossRef]
- Lendahl, U.; Zimmerman, L.B.; McKay, R.D. CNS stem cells express a new class of intermediate filament protein. Cell 1990, 60, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Bernal, A.; Arranz, L. Nestin-expressing progenitor cells: Function, identity and therapeutic implications. Cell. Mol. Life Sci. 2018, 75, 2177–2195. [Google Scholar] [CrossRef] [PubMed]
- Mignone, J.L.; Kukekov, V.; Chiang, A.S.; Steindler, D.; Enikolopov, G. Neural stem and progenitor cells in nestin-GFP transgenic mice. J. Comp. Neurol. 2004, 469, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Yuan, A.; Rao, M.; Veeranna Nixon, R. Neurofilaments and neurofilament proteins in health and disease. Cold Spring Harb. Perspect. Biol. 2017, 9, a018309. [Google Scholar] [CrossRef]
- Bignami, A.; Eng, L.; Dahl, D.; Uyeda, C. Localization of the glial fibrillary acidic protein in astrocytes by immunofluorescence. Brain Res. 1972, 43, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Liu, X.; Liu, T.; Liu, H.; Tong, L.; Jia, S.; Wang, Y.F. Neurochemical regulation of the expression and function of glial fibrillary acidic protein in astrocytes. Glia 2020, 68, 878–897. [Google Scholar] [CrossRef]
- Qian, X.; Shen, Q.; Goderie, S.K.; He, W.; Capela, A.; Davis, A.A.; Temple, S. Timing of CNS cell generation: A programmed sequence of neuron and glial cell production from isolated murine cortical stem cells. Neuron 2000, 28, 69–80. [Google Scholar] [CrossRef]
- Mullen, R.J.; Buck, C.R.; Smith, A.M. NeuN, a neuronal specific nuclear protein in vertebratesxs. Development 1992, 116, 201–211. [Google Scholar] [CrossRef]
- Nagatsu, T.; Levitt, M.; Udenfriend, S. Tyrosine hydroxylase: The initial step in norepinephrine biosynthesis. J. Biol. Chem. 1964, 239, 2910–2917. [Google Scholar] [CrossRef]
- Daubner, S.C.; Le, T.; Wang, S. Tyrosine hydroxylase and regulation of dopamine synthesis. Arch. Biochem. Biophys. 2011, 508, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Tønnesen, J.; Seiz, E.G.; Ramos, M.; Lindvall, O.; Martinez-Serrano, A.; Kokaia, M. Functional properties of the human ventral mesencephalic neural stem cell line hVM1. Exp. Neurol. 2010, 223, 653–656. [Google Scholar] [CrossRef] [PubMed]
- Riederer, B.M.; Draberova, E.; Viklicky, V.; Draber, P. Changes of MAP2 phosphorylation during brain development. J. Histochem. Cytochem. 1995, 43, 1269–1284. [Google Scholar] [CrossRef] [PubMed]
- Dehmelt, L.; Halpain, S. The MAP2/Tau family of microtubule-associated proteins. Genome Biol. 2005, 6, 204. [Google Scholar]
- Mohamed Hassan, Y.; Massa, L.; Caviglia, C.; Sylvest Keller, S. Electrochemical Monitoring of Saos-2 Cell Differentiation on Pyrolytic Carbon Electrodes. Electroanalysis 2019, 31, 256–266. [Google Scholar] [CrossRef]
- Spégel, C.; Heiskanen, A.; Pedersen, S.; Emnéus, J.; Ruzgas, T.; Taboryski, R. Fully automated microchip system for the detection of quantal exocytosis from single and small ensembles of cells. Lab Chip 2008, 8, 323–329. [Google Scholar] [CrossRef]
- Nagel, G.; Szellas, T.; Huhn, W.; Kateriya, S.; Adeishvili, N.; Berthold, P.; Ollig, D.; Hegemann, P.; Bamberg, E. Channelrhodopsin-2, a directly light-gated cation-selective membrane channel. Proc. Natl. Acad. Sci. USA 2003, 100, 13940–13945. [Google Scholar] [CrossRef]
- Warden, M.R.; Cardin, J.A.; Deisseroth, K. Optical neural interfaces. Annu. Rev. Biomed. Eng. 2014, 16, 103–129. [Google Scholar] [CrossRef]
- Luo, J.w.; Nikolic, K.; Degenaar, P. Modelling optogenetic subthreshold effects. In Proceedings of the 2019 41st Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Berlin, Germany, 23–27 July 2019; pp. 6136–6140. [Google Scholar]
- Richner, T.J.; Baumgartner, R.; Brodnick, S.K.; Azimipour, M.; Krugner-Higby, L.A.; Eliceiri, K.W.; Williams, J.C.; Pashaie, R. Patterned optogenetic modulation of neurovascular and metabolic signals. J. Cereb. Blood Flow Metab. 2015, 35, 140–147. [Google Scholar] [CrossRef]
- Stockley, J.H.; Evans, K.; Matthey, M.; Volbracht, K.; Agathou, S.; Mukanowa, J.; Burrone, J.; Káradóttir, R.T. Surpassing light-induced cell damage in vitro with novel cell culture media. Sci. Rep. 2017, 7, 849. [Google Scholar] [CrossRef]
- Senova, S.; Scisniak, I.; Chiang, C.C.; Doignon, I.; Palfi, S.; Chaillet, A.; Martin, C.; Pain, F. Experimental assessment of the safety and potential efficacy of high irradiance photostimulation of brain tissues. Sci. Rep. 2017, 7, 43997. [Google Scholar] [CrossRef] [PubMed]
- Gagnon-Turcotte, G.; Bilodeau, G.; Tsiakaka, O.; Gosselin, B. Smart autonomous electro-optic platforms enabling innovative brain therapies. IEEE Circuits Syst. Mag. 2020, 20, 28–46. [Google Scholar] [CrossRef]
- Gagnon-Turcotte, G.; Keramidis, I.; Ethier, C.; De Koninck, Y.; Gosselin, B. A Wireless Electro-Optic Headstage with a 0.13 μm CMOS Custom Integrated DWT Neural Signal Decoder for Closed-Loop Optogenetics. IEEE Trans. Biomed. Circuits Syst. 2019, 13, 1036–1051. [Google Scholar] [CrossRef] [PubMed]
- Bilodeau, G.; Gagnon-Turcotte, G.; Gagnon, L.L.; Ethier, C.; Timofeev, I.; Gosselin, B. A wireless electro-optic headstage with digital signal processing and data compression for multimodal electrophysiology and optogenetic stimulation. In Proceedings of the 2020 IEEE International Symposium on Circuits and Systems (ISCAS), Seville, Spain, 12–14 October 2020; pp. 1–5. [Google Scholar]
- Chen, C.H.; McCullagh, E.A.; Pun, S.H.; Mak, P.U.; Vai, M.I.; Mak, P.I.; Klug, A.; Lei, T.C. An integrated circuit for simultaneous extracellular electrophysiology recording and optogenetic neural manipulation. IEEE Trans. Biomed. Eng. 2017, 64, 557–568. [Google Scholar] [CrossRef]
- Nguyen, T.K.T.; Navratilova, Z.; Cabral, H.; Wang, L.; Gielen, G.; Battaglia, F.P.; Bartic, C. Closed-loop optical neural stimulation based on a 32-channel low-noise recording system with online spike sorting. J. Neural Eng. 2014, 11, 046005. [Google Scholar] [CrossRef] [PubMed]
- Dufour, S.; De Koninck, Y. Optrodes for combined optogenetics and electrophysiology in live animals. Neurophotonics 2015, 2, 031205. [Google Scholar] [CrossRef]
- Jafari, H.M.; Genov, R. Chopper-stabilized bidirectional current acquisition circuits for electrochemical amperometric biosensors. IEEE Trans. Circuits Syst. I Regul. Pap. 2013, 60, 1149–1157. [Google Scholar] [CrossRef]
- Guo, J.; Ng, W.; Yuan, J.; Li, S.; Chan, M. A 200-channel area-power-efficient chemical and electrical dual-mode acquisition IC for the study of neurodegenerative diseases. IEEE Trans. Biomed. Circuits Syst. 2015, 10, 567–578. [Google Scholar] [CrossRef]
- Dorta-Quiñones, C.I.; Wang, X.Y.; Dokania, R.K.; Gailey, A.; Lindau, M.; Apsel, A.B. A wireless FSCV monitoring IC with analog background subtraction and UWB telemetry. IEEE Trans. Biomed. Circuits Syst. 2015, 10, 289–299. [Google Scholar] [CrossRef]
- Bozorgzadeh, B.; Schuweiler, D.R.; Bobak, M.J.; Garris, P.A.; Mohseni, P. Neurochemostat: A neural interface SoC with integrated chemometrics for closed-loop regulation of brain dopamine. IEEE Trans. Biomed. Circuits Syst. 2015, 10, 654–667. [Google Scholar] [CrossRef]
- Nasri, B.; Wu, T.; Alharbi, A.; You, K.D.; Gupta, M.; Sebastian, S.P.; Kiani, R.; Shahrjerdi, D. Hybrid CMOS-graphene sensor array for subsecond dopamine detection. IEEE Trans. Biomed. Circuits Syst. 2017, 11, 1192–1203. [Google Scholar] [CrossRef] [PubMed]
- Dorta-Quiñones, C.I.; Huang, M.; Ruelas, J.C.; Delacruz, J.; Apsel, A.B.; Minch, B.A.; Lindau, M. A bidirectional-current CMOS potentiostat for fast-scan cyclic voltammetry detector arrays. IEEE Trans. Biomed. Circuits Syst. 2018, 12, 894–903. [Google Scholar] [CrossRef]
- Giagkoulovits, C.; Cheah, B.C.; Al-Rawhani, M.A.; Accarino, C.; Busche, C.; Grant, J.P.; Cumming, D.R. A 16 × 16 CMOS amperometric microelectrode array for simultaneous electrochemical measurements. IEEE Trans. Circuits Syst. I Regul. Pap. 2018, 65, 2821–2831. [Google Scholar] [CrossRef]
- Tsai, J.H.; Kuo, C.Y.; Lin, S.H.; Lin, F.T.; Liao, Y.T. A wirelessly powered CMOS electrochemical sensing interface with power-aware RF-DC power management. IEEE Trans. Circuits Syst. I Regul. Pap. 2018, 65, 2810–2820. [Google Scholar] [CrossRef]
- Zamani, H.; Chan, S.A.; Smith, C.; Mohseni, P. A Neurochemical Recording Microsystem with Analog Background Current Subtraction and 400V/s FSCV Sensing Using a 1st-Order ΔΣM. In Proceedings of the 2020 IEEE 63rd International Midwest Symposium on Circuits and Systems (MWSCAS), Springfield, MA, USA, 9–12 August 2020; pp. 517–520. [Google Scholar]
- Pol, K.J.; Hegt, H.; van Roermund, A.; Ouzounov, S. A femto-ampere sensitive direct-interface current-input sigma delta ADC for amperometric bio-sensor signal acquisition. In Proceedings of the 2015 IEEE Biomedical Circuits and Systems Conference (BioCAS), Atlanta, GA, USA, 22–24 October 2015; pp. 1–4. [Google Scholar]
- Mollazadeh, M.; Murari, K.; Cauwenberghs, G.; Thakor, N. Wireless micropower instrumentation for multimodal acquisition of electrical and chemical neural activity. IEEE Trans. Biomed. Circuits Syst. 2009, 3, 388–397. [Google Scholar] [CrossRef]
- Chiu, W.T.; Lin, C.M.; Tsai, T.C.; Wu, C.W.; Tsai, C.L.; Lin, S.H.; Chen, J.J.J. Real-time electrochemical recording of dopamine release under optogenetic stimulation. PLoS ONE 2014, 9, e89293. [Google Scholar] [CrossRef]
- Budai, D.; Vizvári, A.D.; Bali, Z.K.; Márki, B.; Nagy, L.V.; Kónya, Z.; Madarász, D.; Henn-Mike, N.; Varga, C.; Hernádi, I. A novel carbon tipped single micro-optrode for combined optogenetics and electrophysiology. PLoS ONE 2018, 13, e0193836. [Google Scholar] [CrossRef]
- Cunha, A.B.; Hou, J.; Schuelke, C. Machine learning for stem cell differentiation and proliferation classification on electrical impedance spectroscopy. J. Electr. Bioimpedance 2019, 10, 124. [Google Scholar] [CrossRef]













Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cunha, A.B.; Schuelke, C.; Mesri, A.; Ruud, S.K.; Aizenshtadt, A.; Ferrari, G.; Heiskanen, A.; Asif, A.; Keller, S.S.; Ramos-Moreno, T.; et al. Development of a Smart Wireless Multisensor Platform for an Optogenetic Brain Implant. Sensors 2024, 24, 575. https://doi.org/10.3390/s24020575
Cunha AB, Schuelke C, Mesri A, Ruud SK, Aizenshtadt A, Ferrari G, Heiskanen A, Asif A, Keller SS, Ramos-Moreno T, et al. Development of a Smart Wireless Multisensor Platform for an Optogenetic Brain Implant. Sensors. 2024; 24(2):575. https://doi.org/10.3390/s24020575
Chicago/Turabian StyleCunha, André B., Christin Schuelke, Alireza Mesri, Simen K. Ruud, Aleksandra Aizenshtadt, Giorgio Ferrari, Arto Heiskanen, Afia Asif, Stephan S. Keller, Tania Ramos-Moreno, and et al. 2024. "Development of a Smart Wireless Multisensor Platform for an Optogenetic Brain Implant" Sensors 24, no. 2: 575. https://doi.org/10.3390/s24020575