Effect of Limited Hydrolysis on Traditional Soy Protein Concentrate

Institute for Food Technology, Faculty of Agriculture, University of Belgrade, Nemanjina 6, Serbia
*
Author to whom correspondence should be addressed.
Sensors 2006, 6(9), 1087-1101; https://doi.org/10.3390/s6091087
Submission received: 15 June 2006
/
Accepted: 15 September 2006
/
Published: 17 September 2006
Abstract
:The influence of limited proteolysis of soy protein concentrate on protein extractability, the composition of the extractable proteins, their emulsifying properties and some nutritional properties were investigated. Traditional concentrate (alcohol leached concentrate) was hydrolyzed using trypsin and pepsin as hydrolytic agents. Significant differences in extractable protein composition between traditional concentrate and their hydrolysates were observed by polyacrylamide gel electrophoresis (PAGE) and by SDS-PAGE. All hydrolysates showed better extractability than the original protein concentrate, whereas significantly better emulsifying properties were noticed at modified concentrates obtained by trypsin induced hydrolysis. These improved properties are the result of two simultaneous processes, dissociation and degradation of insoluble alcohol-induced protein aggregates. Enzyme induced hydrolysis had no influence on trypsin-inibitor activity, and significantly reduced phytic acid content.
1. Introduction
Soybeans represent the most important vegetable source of protein ingredients for food formulation [1]. Three major soy protein products are: flour, protein concentrate and soy protein isolate. Soy protein concentrate is a purified, bland protein product containing a minimum of 65% protein on a moisture-free basis (calculated from: N x 6.25). It is obtained from defatted soybean flakes or flour by removal of non-protein components. There are three major methods for preparing soy protein concentrates: the aqueous alcohol wash process, the acidic wash process and the heat denaturation/water wash process. The aqueous alcohol-washed protein concentrate is termed “traditional concentrate” and the acid-washed product is termed “functional concentrate”. Nutritionally, the soy protein concentrate maintains the same excellent nutritional profile of soy flour [2]. The most widely used method based on aqueous alcohol leaching denatures and insolubilize proteins, rendering them poorly functional in food [3].
Glycinin (11S globulin) and β-congycinin (7S globulin) are two major storage soybean proteins and constitute about 87 % of the total proteins [4]. They have complex quaternary structures and high tendency towards association-dissociation reactions. Thus, the composition and structure of these proteins reflects the properties of soy protein products [5].
The physicochemical and functional properties of soy proteins could be changed by physical, chemical and enzymatic treatments. These treatments include heating, pH adjustment, hydrolysis and covalent attachment of other constituents [6]. Many authors [1, 4, 7, 8] have studied the effect of heath treatment under different conditions on different functional properties of soy isolates. It has been reported that the functionality depends basically on the degree of dissociation, denaturation, and aggregation of the glycinin and β-congycinin [9, 10]. Also, as an indicator of modification during preparation processes of commercial and laboratory-prepared soy protein isolates, has been recognized the relationship between solubility and surface hydrophobicity [11].
In the last few years, a significant attention has been given to the limited enzymatic modification of soy proteins. Many approaches have been conducted on the purified soy proteins [12, 13, 14], soy flour and isolate [15, 16, 17, 18]. They have shown that hydrolysates obtained from soy protein isolates and flour can be characterized by physicochemical properties superior to the original isolate or flour. Also, better solubility and foaming properties of modified soy flour and isolate could be obtained through the hydrolysis and subsequent succinylation [19, 20]. However, soy proteins can be extracted enzymatically to the high extent, but not completely. Fisher et al. [21] analyzed the composition of unheated and heat-treated soybean meals after extensive enzymatic extraction at the controlled conditions. They suggested that the protein aggregates from soybean meal proceeded in the enzyme-unextractable residues during hydrolysis. The main cause of that was the low accessibility of aggregates to enzymatic hydrolysis and not theirs resistance to degradation.
The physicochemical changes affecting the functional properties of the proteins were occurred during the preparation of soy protein concentrates from defatted soy flour. The traditional concentrates have poor functionality but high nutritive value and low costs. Better functional properties will make them useful as a replacement ingredient for more expensive isolates and dairy proteins in many food systems. The method based on partial enzymatic hydrolysis is a possible way for this purpose. For this reason the main aim of this study was to investigate the effect of enzymatic modifications of traditional concentrate on extractable protein content and composition, some nutritive and functional properties. Due to the different structure and characteristics of glycinin and β-conglycinin, the knowledge of their alterations during processing is important for the food industry to evaluate the desirable functional properties.
2. Experimental Section
Concentrate preparation. Soybean seeds (Glycine max var. Hodgson) were dehulled and ground in a mill. The meal was than defatted by extraction with n-hexane and air-dried. To prepare the traditional concentrate (TC), non-protein components were extracted by stirring the flour with 65 % aqueous alcohol solution (flour: aqueous alcohol ratio wt/V, 1:10) for 90 minutes at 40°C. Concentrate was separated on the glass filter at the reduced pressure, washed with the same aqueous alcohol solution (1:5 wt/vol), dried over night at 40 °C and ground. Functional concentrate was prepared from defatted flour by extracting with dilute acid (HCl) at pH 4.5 for 90 minutes at 30 °C (flour: dilute acid ratio, 1:10 wt/vol). Concentrate was than separated on glass filter at reduced pressure and washed with the same acid solution. The resulting slurry was neutralized with 1M NaOH and lyophilized.
Modified concentrate preparation. The traditional concentrate was modified with bovine trypsin type III and pepsin (Sigma Chemical Co, St. Louis, USA).Three 10-g portion of the traditional concentrate were each dispersed in 140 ml deionized water and stirred for 15 min to obtain uniform dispersions. The pH of the obtained suspensions was adjusted with 1M NaOH to pH 8.0. After the incubation for 15 min at 37°C, the dispersions were hydrolyzed with 10 ml of 0.02 % trypsin solution. The enzyme to substrate (E/S) ratio was 0.2 mg/g. The first dispersion was hydrolyzed for 30 min (TMC30), the second dispersion was hydrolyzed for 60 min (TMC60) and the third dispersion was hydrolyzed for 90 min (TMC 90) with constant stirring. The resulting trypsin hydrolysates were each adjusted to pH 7.0 and heated at 90˚C for 3 min to inactivate the enzyme. The hydrolysates were than freeze-dried and ground. The heat treated and alkali-treated (AMC) controls were also prepared similarly without added enzyme. The traditional concentrate was also modified with pepsin for 60 min at 37°C (PMC). The resulting pepsin hydrolysate was adjusted to pH 7, heated at 90˚C for 3 min, freeze-dried and ground.
Protein determination. The total protein content was determined by the micro-Kjeldahl method [22] using a nitrogen to protein conversion factor of 6.25. To determine the extractable soluble protein content, the protein was extracted according to the method of [23]. The soy flour and concentrates (2g) were extracted for 120 min at room temperature with Tris-HCl buffer pH 8.0 (the sample to buffer ratio was 1: 20) and were centrifuged at 17000 x g for 15 min. The protein content in the supernatant was determined according to the method of Lowry et al. [24] using bovine serum albumin (Sigma, USA) as a standard. The protein extractability was expressed as the milligram of the extractable soluble protein per gram of the dry sample.
PAGE. The protein hydrolysates were analyzed by PAGE following the general procedure of Davis [25] using 7 % (wt/vol) acrylamide gels. Briefly, the proteins, extracted with 0.03 M Tris-HCl buffer containing 0.01M 2-mercaptoethanol (pH 8.0), were diluted to 2 mg/mL with the sample buffer pH 8.0. A 25 μl sample was loaded per well. The gels were run at 30 mA per gel for 7 hours to completion. Gels were fixed, stained with 0.1 % (wt/vol) Coomassie Blue R-250 [dissolved in 12% (vol/vol) acetic acid, and 50% (vol/vol) methanol] for 45 min and destained with 7% (vol/vol) acetic acid and 5% (vol/vol) methanol for 48h.
SDS-PAGE. SDS-PAGE was performed according to the procedures of Fling and Gregerson [26] using 5% (wt/vol) stacking and 12.5% (wt/vol) resolving gel. Prior to electrophoresis, the protein extract was diluted to 2mg/mL with sample buffer (0.055 M Tris-HCl, pH 6.8, 2% (wt/vol) SDS, 7% (vol/vol) glycerol, 4.3% (vol/vol) β-mercaptoethanol, 0.0025% (wt/vol) bromophenol blue), heated at 90 ˚C for 5 min and cooled at the room temperature. A 25 μl sample was loaded per well. The gels were run at 30 mA per gel for 6 hours to completion. Gels were fixed, stained with 0.23 % (wt/vol) Coomassie Blue R-250 [dissolved in 3.9 % (wt/vol) trichloroacetic acid (TCA), 6% (vol/vol) acetic acid, and 17% (vol/vol) methanol] for 45 min and destained with 8% acetic acid and 18% (vol/vol) ethanol. Molecular weights of the polypeptides were estimated by using low molecular weight calibration kit (Pharmacia, Sweden). Molecular weight markers included: phosphorylase B (94.0), bovine albumin (67.0), ovalbumin (43.0), carbonic anhydrase (30.0), soybean trypsin inhibitor (20.1), and α-laktalbumin (14.4). Also, the identification was done using 7S and 11S protein fraction obtained according to the procedure of Than and Shibasaki [23].
SDS-PAGE as well as PAGE were performed with electrophoresis unit LKB-2001-100 in conjunction with power supply LKB-Macrodrive 5 and LKB-Multitemp as a cooling unit (LKB, Sweden).
Densitometric analysis. The destained gels were scanned by Scanexpress 12000SP (Mustek, Germany) and analyzed by SigmaGel software version 1.1 (Jandel Scientific, San Rafalel, CA). The quantitative estimation of each identified subunits was calculated as the percentage of the corresponding area of the subunits with respect to the total area of the densitogram.
Emulsifying properties. Emulsifying properties were measured according to a modified method of Wu et al [27]. Pure sunflower oil (15 mL) and 45 mL 0.1% protein solution, prepared using protein extracted from defatted meal as described for protein determination, were homogenized in a mechanical homogenizer at the highest settings for 1 min. Fifty-micro liter portions of the emulsions were pipetted from the bottom of the container at 0 and 10 min after homogenization. Each portion was diluted with 10mL of 0.1% (wt/vol) SDS solution. Absorbances of these diluted emulsions were measured at 500nm. The absorbances measured immediately (A0) and 10 min (A10) after emulsion formation were used to calculate the emulsifying activity index (EAI) and the emulsifying stability index (ESI):
where T = 2.303; A0 = absorbance measured immediately after emulsion formation; dilution factor = 200; C = weight of protein/unit volume (g/ml) of aqueous phase before emulsion formation; Φ = oil volume fraction of the emulsion; and
where Δt = 10 min and ΔA = Ao – A10.
Trypsin-inhibtor (TI) activity. The Ti-activity was assayed and quantitated as described by the method of Liu and Markakis [28] using α-N-benzoyl-DL-arginine-p-nitroanilide hydrochloride as a substrate. The samples were extracted with distilled water [,] for 30 min on a mechanical shaker. The extract was filtered through No 4 Whatman paper. A 10 ml of the extract was diluted with 0.05 M Tris/HCl buffer pH 8.2 (1:1, extract/buffer) and filtered. The filtrate was then further diluted with distilled water (1:5, filtrate/water). One ml of the diluted filtrate was incubated with 1 ml of 0.92 mM BAPA and 16μg/mL enzyme solution (in the 0.001M HCl) at 37 ◦C for 10 min. The reaction was stopped by the addition of 30% (vol/vol) acetic acid. The reaction was also run in the absence of inhibitors by replacing the sample with 1ml of water. The absorbance was measured at 410 nm (Unicam, England). Distilled water was used as a blank. Defining a trypsin unit as an A 410 increase of 0.01 under the conditions of the assay, the inhibitor activity was expressed in trypsin units inhibited (TUI) per milligram of the dry sample.
Phytic acid (PA) content determination. Phytic acid content was measured according to the supernatant difference method of Thompson and Erdman [29] modified by Velickovic et al. [30]. The content of phytic acid was calculated by multiplying the amount of phytic acid phosphorus with the corresponding conversion factor (3.55).
Statistical analysis. All experiments were repeated three times except for electrophoretic analysis, which were duplicated. The data were analyzed using Statistica software ver 5.0 (StatSoft Co., USA). The significance of differences between means was determined by t-test procedure for independent samples at P < 0.05. The results are given as the mean values.
3. Results and Discussion
3.1. Protein extractability
The protein extractabality was measured in Tris-HCl (pH 8.0) extracts of concentrates and hydrolysates. The buffer was selected to identify the components that may participate as the extractable fraction. The method of SPC preparation, i.e., acid or alcohol wash, significantly influenced the extractability of soy protein concentrate, as shown in Table 1. Due to the alcohol induced denaturation, traditional alcohol-washed concentrate had the lowest extractability (247.01 ± 1.26 mg/g). The acid wash resulted in significantly higher extractability than the alcohol wash. Also, protein content of FC was higher than that from TC. These data suggest that the acid wash removed more soluble sugars and recovered relatively more proteins than the alcohol wash.

{kind=link}
{kind=link}
{kind=link}
| Sample** | Extractability
mg/g | Total protein (%)
(N x 6.25) |
| defatted soy flour | 388.30±0.75a | 51.66a |
| TC | 247.01±1.26b | 66.02b |
| FC | 524.76±3.80c | 68.08c |
| AMC | 304.30±0.46d | 67.04d |
| TMC30 | 507.40±0.75e | 70.02e |
| TMC60 | 615.90±2.09f | 70.36e |
| TMC90 | 660.50±1.85g | 70.76 e, f |
| PMC | 466.40±0.59h | 71.30f |
Means with different letters are significantly different (p<0.05), **TC-traditional protein concentrate; FC- functional concentrate; AMC-samples treated at 37 °C and pH 8.0 without added enzyme; TMC30, TMC60 and TMC90- samples treated with trypsin for 30, 60 and 90 minutes, respectively; PMC- samples treated with pepsin.
The limited proteolysis is an effective way to improve the traditional protein concentrate extractability. The protein extractability of modified samples varied from ca. 46% to 66%, depending upon the enzyme used. Trypsin increased the extractability most efficiently during the initial 30 min of incubation, reaching about two fold rise (507.4 ± 0.75 mg/g). After that, a slightly lower increase was observed. The average protein extractability of 90 min-treated samples was 660.50 mg/g. It is interesting to note that the control samples treated at pH 8.0 without added enzyme (AMC) had higher protein extractability (304.30±0.46) than the initial concentrates. These data suggest that the improved protein extractability of trypsin-modified samples is mostly the result of the enzyme activity, but partly could be due to the hydrolytic effect of mild alkali hydrolysis. The synergistic effect of an alkali pretreatment (pH 10) on the subsequent papain-induced hydrolysis of the soy protein isolate has been reported by Wu et al. [27]. Protein extractability of traditional concentrate was also improved with pepsin although the effect was lower (466.40 mg/g) than for trypsin-modified concentrates. According to our results, trypsin was more effective for proteolysis than pepsin. This may be accounted partly for compositional features of traditional soy protein concentrate. The major soy proteins, 7S and 11S globulins contain a larger number of basic amino acids than aromatic ones and thus may contain more cleavage sites for trypsin to act on. Also, it is possible that the products formed during pepsin-induced hydrolysis aggregate and become insoluble at pH 8.0.
3.2 Extractable protein composition
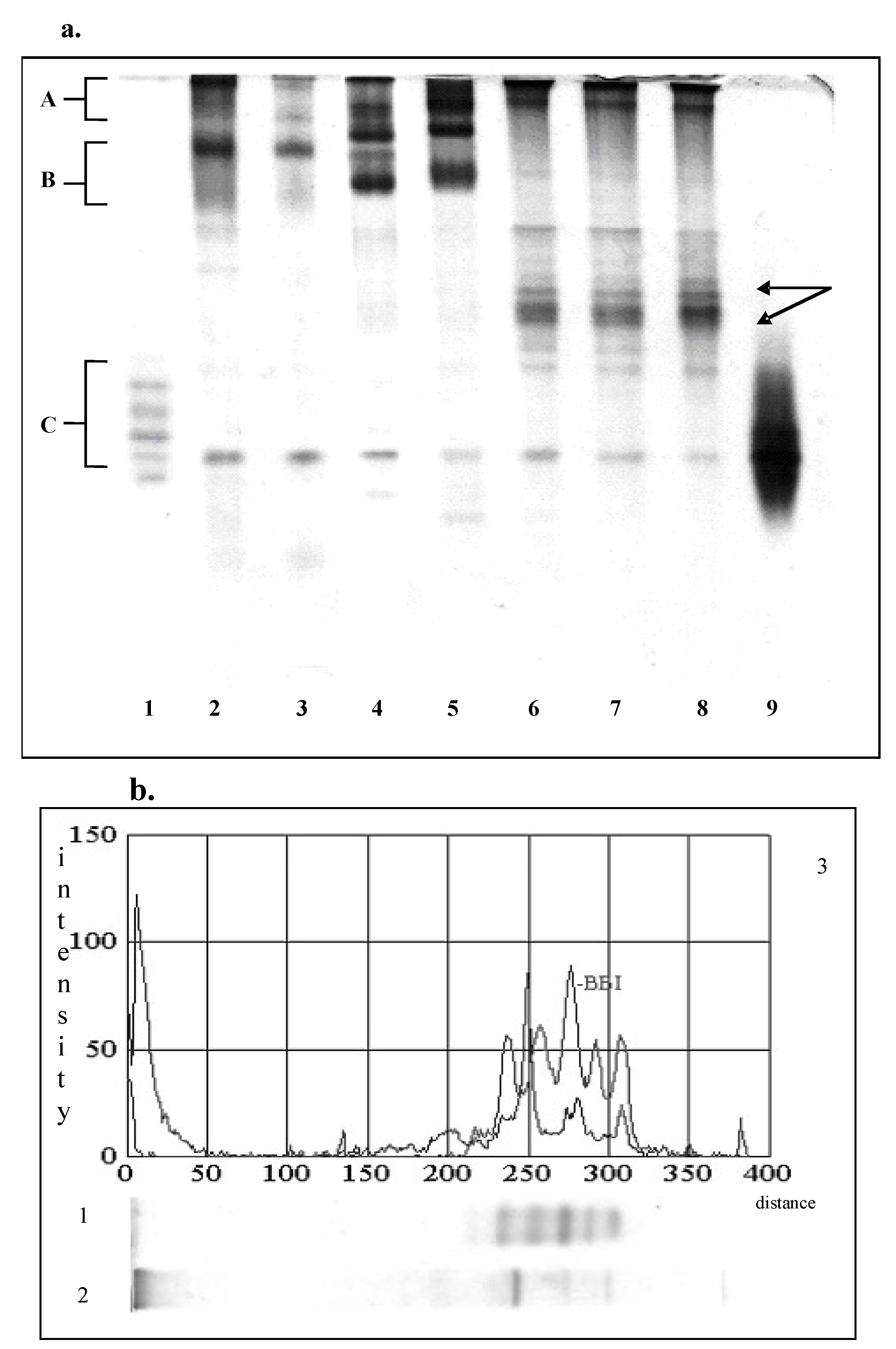
PAGE and SDS-PAGE separated extractable proteins of soy flour into multiple components. PAGE-pattern of the extractable soluble proteins of soy flour contained five major protein bands (Fig.1a, line 2). Two of them (Zone A) were identified as β-conglycinin, and two (zone B) as glycinin [23]. The band with lower electrophoretic mobility in zone B represent dimeric form of glycinin, whereas the other represents dissociating form of glycinin molecule i.e., monomeric form. The bands in zone C (Figure 1a) corresponds to the forms of Kunitz and Bowman-Birk types of inhibitor [31].
As shown in Figure 1a (line 3) the major soy flour protein expressed different stability to the alcohol-induced denaturation; the most susceptible were the forms of β-conglycinin. As a result of their insolubilization, dimeric form of glycinin dominated on the PAGE-pattern of the traditional concentrate.
As one can see, the enzymic treatments caused significantly different composition of the extractable proteins. Trypsin-induced hydrolysis promoted the degradation of the insoluble protein aggregates, hydrolysis of the major soluble fractions and the formation of new soluble products. During the initial 30 min of hydrolysis, the bands of glycinin were disappeared, almost completely and were observed as a diffused zone (Figure 1a, line 6). As a result of trypsin acting during the initial incubation, several new protein bands with higher relative mobility were noticed. Three of them (indicated in Figure 1a with arrows) were the most intensive and were retained even after 90 minutes of trypsin acting. This suggests that these fractions are the stable intermediate products. According to the investigation of Shutov et al [14], they could be intermediate products of glycinin. Also, they could be partly degraded protein aggregates.
Figure 1.
PAGE-analysis of the extractable soluble proteins of soy flour, traditional and modified concentrates.(a) 1. Standard of Bowman-Birk inhibitor, 2. Raw soy flour, 3. Traditional concentrate,4. heat treated sample, 5. Functional concentrate, 6, 7, 8. Samples modified with trypsin for 30, 60, 90 minutes, respectively; (b) 1.Bowman-Birk inhibitor, 2. Traditional concentrate modified with pepsin. 3. Densitometric analysis of BBI and pepsin-modified sample.
Figure 1.
PAGE-analysis of the extractable soluble proteins of soy flour, traditional and modified concentrates.(a) 1. Standard of Bowman-Birk inhibitor, 2. Raw soy flour, 3. Traditional concentrate,4. heat treated sample, 5. Functional concentrate, 6, 7, 8. Samples modified with trypsin for 30, 60, 90 minutes, respectively; (b) 1.Bowman-Birk inhibitor, 2. Traditional concentrate modified with pepsin. 3. Densitometric analysis of BBI and pepsin-modified sample.
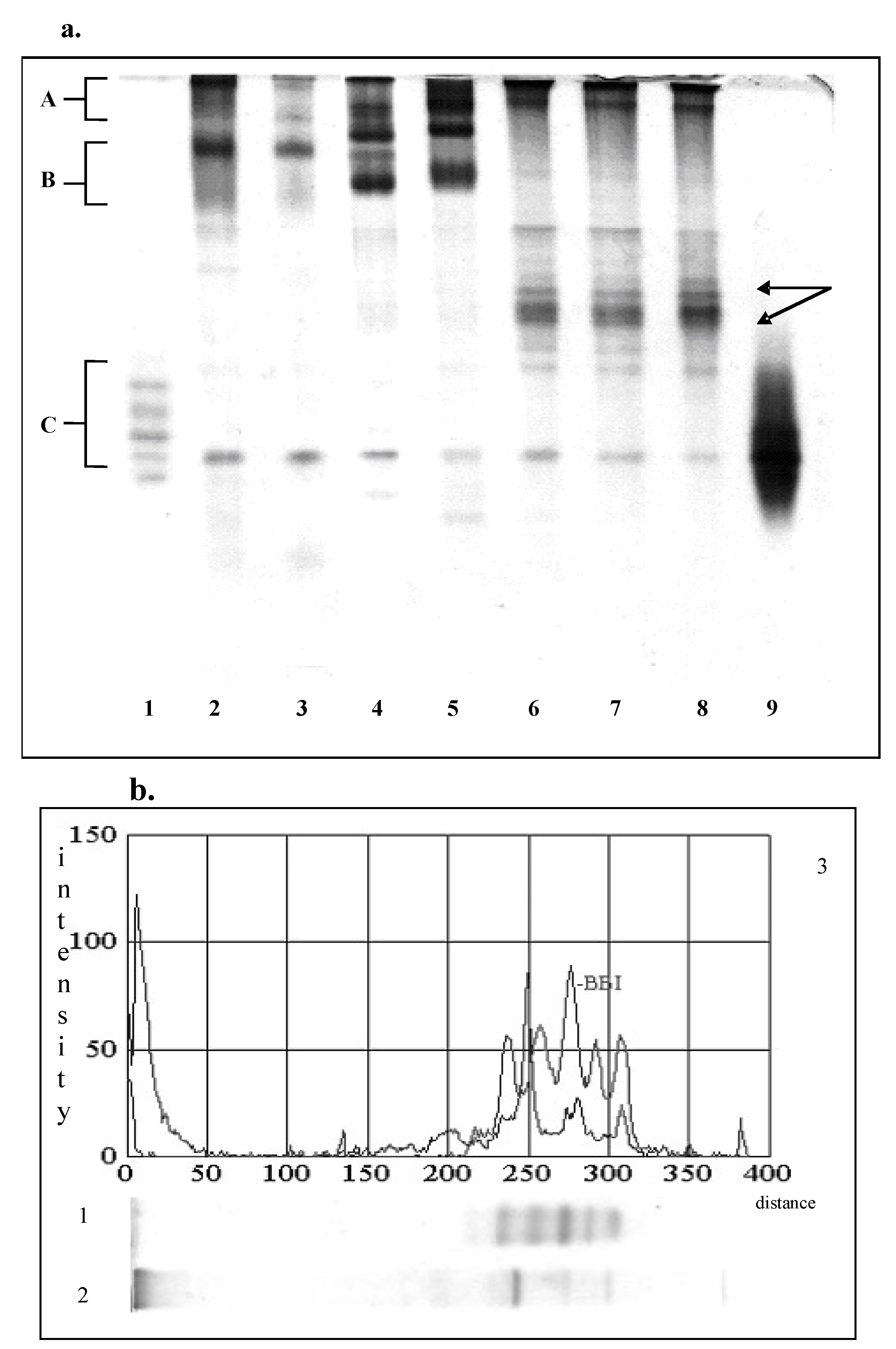
PAGE-pattern of pepsin-modified sample (Figure 1b, line 2) is significantly different from trypsin-modified concentrates. Since the fraction B (the fraction of glycinin) was disappeared completely; the band noticed in the zone of fraction A is dominant. This may imply that the fraction A was more resistant to the enzymatic breakdown than the other ones. On the other hand, it could be a new soluble aggregate formed from low molecular weight products of hydrolysis. Furthermore, several protein bands with higher relative mobility could be observed. According to the electrophoretic mobility, seven of them were in zone of Bowman-Birk and Kunitz inhibitors.
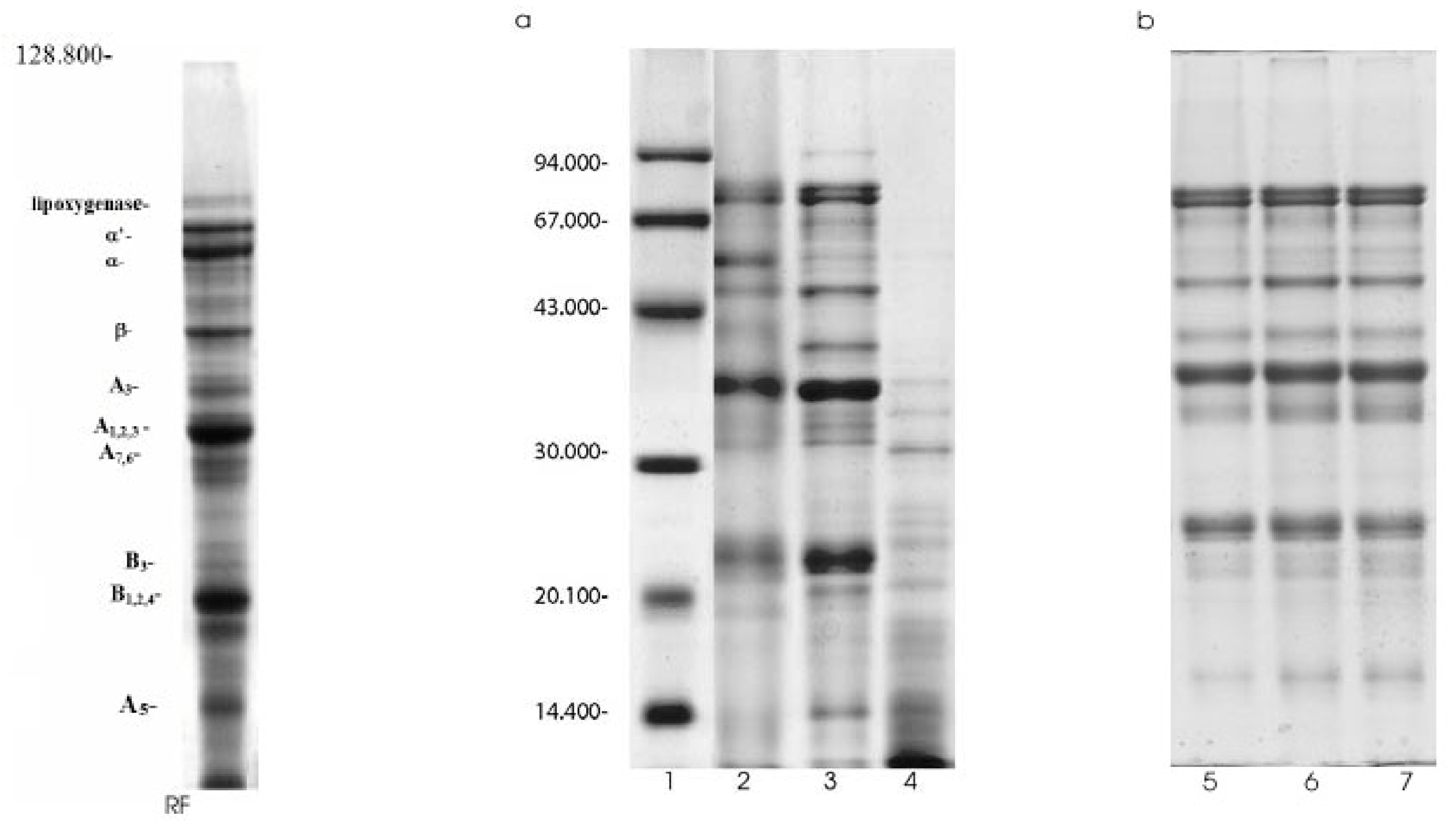
SDS-PAGE separated extractable proteins into subunits and polypeptides (Figure 2a,b). The concentration of protein subunits is shown in Table 2. The SDS-electrophoretic profile of raw soy flour (Fig. 2a, line RF) contained three major bands (mw 81 kDa, 74 kDa, 49 kDa) identified as α`-, α- and β- subunits of β-conglycinin, and two major (MW 36.3 kDa, 22.7 kDa) and four minor bands (MW 39.7, 32.1, 25.5, 14.9 kDa) of acidic and basic subunits of glycinin. Also, the minor 93.9 kDa-band were identified as lipoxygenase. The SDS-electrophoresis profile of TC (Fig. 2a, line 2) showed that the lipoxygenase was disappeared, whereas the -α`-α- and β- subunits of β-conglycinin were registered as diffused bands. Nevertheless, β-conglycinin subunits content was 17.74% and similar to the value obtained for soy flour extracts (17.90%, Table 2). Also, due to the degradation of high molecular weight polypeptides (polypeptides of lipoxygenase, β- and γ-conglycinin, and β-amylase) a more intensive band of 57.1 kDa was observed. Moreover, glycinin sustained similar changes during the aqueous alcohol washing process. Namely, the content of acidic subunits was reduced, whereas the basic subunits were remarked as diffused zone represented with 18.96% of the extractable soluble proteins. The data shown in Table 2 indicate that acidic subunits were more susceptible to the alcohol wash than the basic ones, especially –A3- and -A5- polypeptides. Their content was reduced to 1.95% and 4.17%, respectively. Different influence of the alcohol treatment on the glycinin subunits was probably the consequence of its compact quaternary structure. It seems that the alcohol induced denaturation proceeds initially on the acid subunits and later on the basic ones. It is in agreement with the localization of the basic subunits in the interior of the glycinin molecule [32].
Figure 2.
SDS-PAGE analysis of modified traditional concentrates; RF-raw flour, 1.Molecular weight standards, 2.Traditional concentrate, 3. Functional concentrate,4. Pepsin-modified concentrate, 5,6,7 Traditional concentrate modified for 30, 60 and 90 minutes, respectively.
Figure 2.
SDS-PAGE analysis of modified traditional concentrates; RF-raw flour, 1.Molecular weight standards, 2.Traditional concentrate, 3. Functional concentrate,4. Pepsin-modified concentrate, 5,6,7 Traditional concentrate modified for 30, 60 and 90 minutes, respectively.
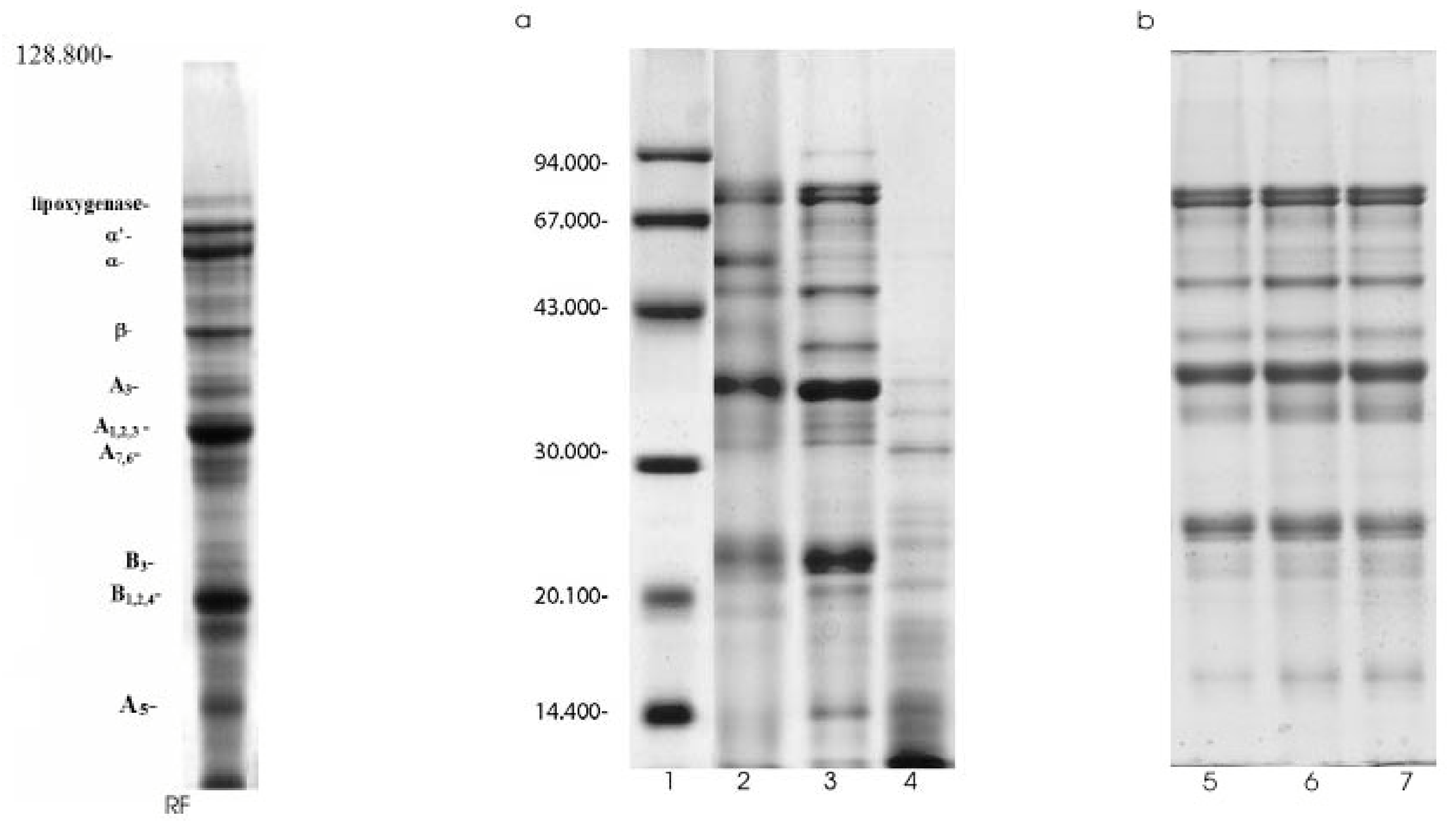
The Figure 2b shows the SDS-patterns of trypsin-modified concentrates, whereas the results of densitomeric analysis are presented in Table 2. The data obtained for TMC 30 and TMC 60 indicate that trypsin induce dissociation of unsoluble alcohol induced aggregates, resulting in an increase of β-conglycinin and glycinin content. Namely, their content increased from 17.74% to 28.02% and 41.53% to 54.58%, respectively. Further hydrolysis (90 min) generates an decrease of B subunits content and slightly increases the content of low molecular weight products.
Table 2.
The subunits composition of Tris-HCl extracts of soy flour, traditional and trypsin-modified concentrates.
| Protein | Sample | ||||
|---|---|---|---|---|---|
| subunit | Raw soy flour | TC | TMC30 | TMC60 | TMC90 |
| β-conglucinin | Concentration (%) | ||||
| α`- | 5.04 | 6.48 | 8.33 | 10.86 | 8.59 |
| α- | 6.07 | 5.74 | 9.96 | 10.00 | 10.89 |
| β- | 6.79 | 5.52 | 8.59 | 7.16 | 7.66 |
| ∑ | 17.9 | 17.74 | 26.88 | 28.02 | 27.14 |
| Glycinin | |||||
| A3- | 3.93 | 1.95 | 5.33 | 6.56 | 6.74 |
| A1,2,4- | 16.05 | 12.94 | 19.13 | 22.23 | 20.28 |
| A5- | 7.39 | 4.17 | 2.45 | 2.66 | 4.79 |
| A7,6- | 2.45 | 3.51 | 6.70 | 7.83 | 7.42 |
| B1,2,3,4- | 18.87 | 18.96 | 15.75 | 15.30 | 10.70 |
| ∑ | 48.69 | 41.53 | 49.36 | 54.58 | 49.93 |
TC-traditional soy protein concentrate; TMC30, TMC60 and TMC90 – samples modified for 30, 60 and 90 minutes with trypsin.* the values are the means of two densitometric determination.
The incubation of TC in the presence of pepsin resulted in the intensive disappearance of 7S and 11S protein subunits (Figure 2a, line 4). Minor protein bands of -α, -α`-, -β- subunits of β-conglycinin and acid -A 1.2.4- subunits of glycinin were only observed. Thus, the low molecular mass products (<27 kDa) dominated. Apparently, these polypeptides constitute high-molecular-mass aggregate (fraction A) detected on PAGE pattern of pepsin-treated sample (Figure 1b, line 2).
The results of this study indicate that the two processes, dissociation and degradation of the unsoluble aggregates, take place simultaneously during trypsin induced hydrolysis. In the initial phase of the treatment (30-60 min), the dissociation process dominated, resulting in the increase of the protein extractability. Noticeable decrease of the alcohol induced degradation products of 57.1kDa was detected in this stage of hydrolysis. After 60 min of trypsin acting, the degradation process became apparent. Basic polypeptides as well as low molecular weight products (10-2-15kDa) were registered as the diffuse zone. Relatively low percent of new protein bands were also observed. Different susceptibility of glycinin subunits to trypsin hydrolysis, as observed in the proceeding paper, could be explained by the facts that (i) trypsin preferentially hydrolyzes peptide bonds adjacent to the basic amino acids [33,34]; and (ii) the alkali-pretreatment facilitated protease to hydrolyze basic glycinin subunits [27]. Low percent of new protein bands suggests that during 90 min the “one- by- one” mechanism of hydrolysis is predominant. This mechanism proposes the first slow step in which the molecules are hydrolyzed one-by-one to yield the final products, and consequently no intermediate products of hydrolysis can be detected [35].
3.3 Emulsifying properties
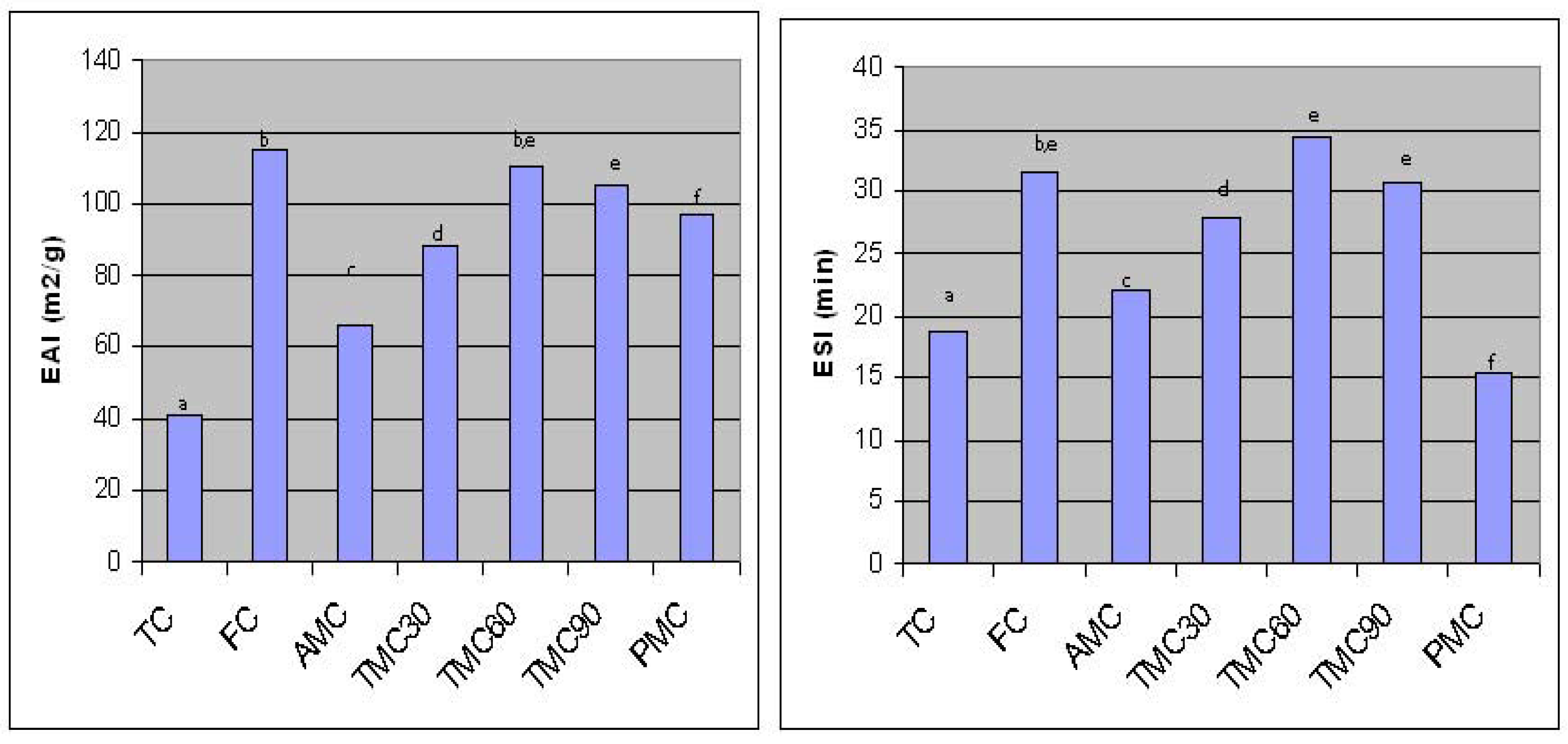
The emulsifying properties of TC and hydrolysates are shown in Figure 3. TC had the lower emulsifying activity and the low ability to form a stable emulsion. Trypsin and pepsin induced hydrolysis significantly (p<0.001) improved the emulsifying activity index of TC from 41 to 88, 110, 105 and 97 m2/g for TMC30, TMC60, TMC90 and PMC, respectively (Fig. 3). No significant difference was found between TMC60 and TMC90. The lower EAI of PMC compared to that of TMC60 and TMC90 could be due to the lower surface hydrophobicity of PMC. After partial hydrolysis, the protein structure was altered, as observed on PAGE and SDS-PAGE gels, which may improve the emulsifying properties. The hydrolysates with higher extractability and smaller molecular size might facilitate the diffusion and spread at oil-water interfaces. In addition, partial hydrolysis could lead to unfolding of protein molecules. Nonpolar amino acid groups, buried inside protein molecules, could be exposed on the surface of protein molecules after unfolding. These exposed hydrophobic groups enhanced the interaction between protein and lipids. Improvement in emulsifying activity due to higher surface hydrophobicity of protein molecules is well documented [36, 37, 38].
Figure 3.
Emulsifying properties (EAI and ESI) of traditional and enzyme-modified concentrates* TC-traditional soy protein concentrate; FC-functional concentrate; TMC30, TMC60 and TMC90 – samples modified with trypsin during 30, 60 and 90 minutes; PMC – samples modified with pepsin. *Means with different letters are significantly different.
Figure 3.
Emulsifying properties (EAI and ESI) of traditional and enzyme-modified concentrates* TC-traditional soy protein concentrate; FC-functional concentrate; TMC30, TMC60 and TMC90 – samples modified with trypsin during 30, 60 and 90 minutes; PMC – samples modified with pepsin. *Means with different letters are significantly different.
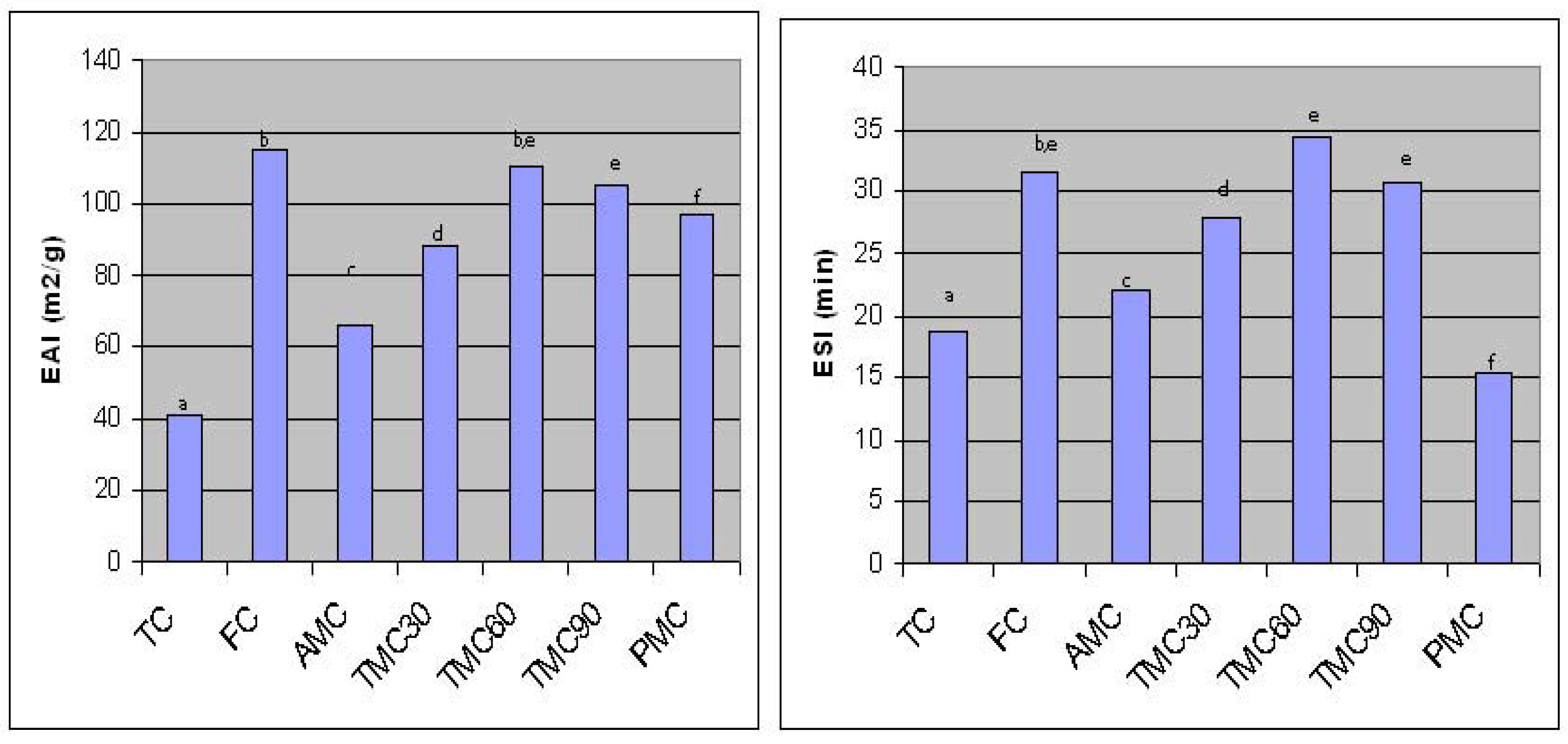
Trypsin modification also significantly (p<0.05) improved the ESI of TMC30, TMC60 and TMC90 from 18.72 min to 27.85, 34.37 and 30.72 min, respectively (Fig.3). No significant difference in ESI (p < 0.05) was found between TMC60 and TMC90. Emulsions prepared with 60 min-hydrolyzed samples were more stable than those prepared with the functional concentrates. On the other side, although pepsin acting increased EAI of traditional concentrate, the emulsions prepared with these samples were less stable than those prepared with the unmodified concentrate. These results indicate that enzyme induced protein alterations had significant influence on emulsifying stability. The smaller protein spaces, produced during the dissociation and degradation processes of the unsoluble aggregates, might have higher molecular flexibility, which improve the ability of proteins to adsorb at the interface and form stable emulsions. However, the decrease in emulsion stability of PMC could be due to production of more small protein spaces in PMC as compared to trypsin modified hydrolysates (87.67% of the extractable soluble polypeptides, as observed in SDS-PAGE). These small polypeptides could have lower surface hydrophobicity than larger ones. The effect of protein hydrolysis on surface hydrophobicity is well known [27]. This observation indicated that the emulsion stability could be decreased by excessive enzyme hydrolysis.
3.4 Trypsin-inhibitor activity and phytic acid content
The trypsin-inhibitor activity is determined in the concentrates and hydrolysates, as show in Table 3. Residual inhibitor activity registered in the traditional concentrate was 17.34% of row flour. According to the data reported in the available literature [39], this activity has no antinutritive effect in nutrition. Intensive band of extractable KTI registered on PAGE pattern (Fig. 1, line 3), as well as the low residual activities suggest that most of KTI exists in partially disrupted native conformation and inactivated form. The presence of an inactive form of inhibitor is important from the stand of their amino acids composition and potential health benefits. Further decrease of the inhibitor activity during the enzyme modifications (15.15-17.31%) has no practical significance.
As shown in Table 3, no significant differences (p>0.05) were found between TC and FC in phytic acid content. However, the enzyme induced modifications significantly reduce the phytic acid content to 4.03 ° 0.03 %– 3.47 ° 0.05 %. No significant differences were found among hydrolisates. In the traditional nutritional theory the phytic acid has been considered as an antinutritient. Namely, phytic acid could interact with proteins and decreases their digestibility [40] and can reduce the bioavailability of minerals at high concentrations [41]. On the contrary, it is well known today that the low concentration of PA has some beneficial effects; these include controlling dental caries; improving the oxygen-providing ability of red blood cells; the cancer preventing activity [40,41]. Thus, the high content of phytic acid in modified soy protein concentrates may be desirable.
Table 3.
Trypsin inhibitor activity and phytic acid content of traditional and modified concentrates*
| Sample | Residual Ti-activity | Phytic acid** % | |
| TUI/g** | % | ||
| Row flour | 178.50 ±0.71 a | 100 | 4.10 ±0.07 a |
| TC | 31.57 ±0.80 b | 17.34 | 4.95 ±0.03b |
| FC | 135.62 ±0.08 c | 76.01 | 4.81±0.05 b |
| AMC | 29.61± 1.86b,d | 15.51 | 3.89±0.13a,c,d |
| TMC30 | 29.50 ±0.79d | 17.31 | 4.03 ±0.03 a |
| TMC60 | 26.78 ±0.76b,e | 15.15 | 3.65 ±0.05 e,c |
| TMC90 | 27.50 ±1.80 e,d | 15.56 | 3.47±0.05c,d,e |
| PMC | 27.47±0.60e,d | 15.66 | 3.60±0.03e,c |
*TC-traditional protein concentrate; FC- functional concentrate; AMC-samples treated at 37±C and pH 8.0 with no enzyme added; TMC30, TMC60 and TMC90- samples treated with trypsin for 30, 60 and 90 minutes, respectively; PMC- samples treated with pepsin. **Means with different letters are significantly different (p<0.05)
4. Conclusion
Processing method had significant effect on the extractability of soy protein concentrates. The limited enzymatic hydrolysis is a simple method for improvement of the extractability of the traditional soy protein concentrate. The hydrolysates prepared with trypsin are characterized with high content of extractable proteins, better emulsifying properties than the traditional concentrate, acceptable level of trypsin-inhibitor activity and phytic acid content. The improvement of these properties is the result of the enzyme induced alterations of major soybean proteins.
References
- Sorgentini, A. D.; Wagner, J. R.; Anon, M. C. Effect of thermal treatment soy protein isolate on the characteristics and structure- function relationship of soluble and insoluble fractions. J. Agric. Food Chem. 1995, 43, 2471–2479. [Google Scholar] [CrossRef]
- Beery, K. E. Preparation of soy protein concentrate products and their application in food systems. In Proceedings of the World Congress of Vegetable Proteins Utilization in Human Food and Animal Food Stuffs; Applewhite, TH, AOCS, Champaign, USA, 1989; pp. 62–67. [Google Scholar]
- Kinsella, J. E.; German, B.; Damodoran, s. Physicochemical and functional properties of oilseed proteins with emphasis on soy proteins. In New Protein Foods; Altschul, A.M., Wilke, H.L., Eds.; Academic Press: New York, 1985; Vol. 5, pp. 107–179. [Google Scholar]
- Kinsella, J. E. Functional properties of soy proteins. J. Am. Oil Chem. Soc. 1979, 56, 242–258. [Google Scholar] [CrossRef]
- Rhee, K. C. Functionality of soy proteins. In Protein Functionality in Food Systems; Hettiarachchy, N. S., Ziegler, G. R., Eds.; Dekker, New York, 1994; pp. 311–324. [Google Scholar]
- Chobert, J. M; Sitohy, M.Z.; Whitaker, D. J.R. Specific limited hudrolysis and phosphorylation of food proteins for improvement of functional and nutritional properties. J.Am. Oil Chem. Soc 1987, 36, 1704–1711. [Google Scholar] [CrossRef]
- Petrucelli, S.; Anon, M. C. The relationship between the method of obtention and the structural and functional properties of soy protein isolates 1. Structural and hydration properties. J. Agric. Food Chem 1994a, 42, 2161–2169. [Google Scholar] [CrossRef]
- Petrucelli, S.; Anon, M. C. The relationship between the method of obtention and the strustural and functional properties of soy protein isolates 2. Surface properties. J. Agric. Food Chem 1994b, 42, 2170–2176. [Google Scholar] [CrossRef]
- Wagner, J. R.; Anon, M. C. Influence of denaturation degree, hydrophobicity and sulphydryl content on solubility and water absorbing capacity of soy protein isolates. J. Agric. Food Chem. 1990, 50, 765–770. [Google Scholar]
- Arrese, E.L.; Sorgentini, D. A.; Wagner, .J.R.; Annon, M.C. Electrophoretic, solubility, and functional properties of commercial soy protein isolates. J. Agric .Food Chem 1991, 39, 1029–1032. [Google Scholar] [CrossRef]
- Wagner, J. R.; Sorgentini, D. A; Anon, M. C. Relation between solubility and surface hydrophobicity as an indicator of modifications during preparation processes of commercial and laboratory-prepared soy protein isolates. J. Agric. Food Chem. 2000, 48, 3159–3165. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Park, P.S.W.; Rhee, K.C. Functional properties of proteolytic enzyme modified soy protein isolate. J. Agric. Food Chem. 1990, 33, 651–656. [Google Scholar] [CrossRef]
- Kamata, Y.; Otsuka, S.; Sato, M.; Shibasaki, K. Limited proteolysis of soybean beta-conglycinin. Agric. Biol. Chem. 1982, 46, 2829–2834. [Google Scholar] [CrossRef]
- Shutov, A.D.; Pineda, V. I.; Senyuk, V.; Reva, V. A; Vaintraub, I. A. Action of trypsin on glycinin. Eur.J.Biochem. 1991, 199, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Were, L.; Hettiarachchy, N. S.; Kalapathy, U. In vitro accessibility of untreated, toasted and extruded soybean meals for proteases and carbohydrases. J. Food Sci. 1997, 62, 821–823. [Google Scholar] [CrossRef]
- Calderon de la Barca, A. M.; Ruiz-Salazar, R. A.; Jara-Marini, M. E. Enzymatic hydrolysis and synthesis of soy protein to improve its amino acid composition and functional properties. J. Food Sci. 2000, 65, 246–253. [Google Scholar] [CrossRef]
- Molina Ortiz, S.E.; Wagner, J.R. Hydrolysates of native and modified soy protein isolates: structural characteristics, solubility and foaming properties. Food Res. Int. 2002, 35, 511–518. [Google Scholar] [CrossRef]
- Hrackova, M.; Rusnakova, M.; Zemanovic, J. Enzymatic hydrolysis of defatted soy four by three different proteases and their effect on the functional properties of resulting protein hydrolysates. Czech Journal of Food Science 2002, 20, 7–14. [Google Scholar]
- Achouri, A.; Zhang, W.; Xu, S. Enzymatic hydrolysis of soy protein isolate and effect of succinylation on the functional properties of resulting hydrolysates. Food Res. Int. 1998, 31, 617–623. [Google Scholar] [CrossRef]
- Achouri, A.; Zhang, W. Effect of succinylation on the physicochemical properties of soy protein hydrolysate. Food Res. Int. 2001, 56, 507–514. [Google Scholar] [CrossRef]
- Fischer, M.; Gruppen, H.; Piersma, S. R.; Kofod, L. V.; Schols, H. A.; Voragen, A. G. J. Aggregation of peptides during hydrolysis as a cause of reduced enzymatic extractability of soybean meal proteins. J. Agric. Food Chem. 2002, 50, 4512–4519. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Association of Official Analytical Chemists, Official methods of analysis, 12 edn. 1975; Washington, DC. [Google Scholar]
- Thanh, V. H.; Shibasaki, K. Major proteins of soybean seeds. A straightforward fractionation and their characterization. J. Agric. Food Chem. 1976, 24, 1117–1121. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R. J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Davis, J. Disc Electrophoresis- II Method and application to human serum proteins. Annals N. Y. Ac. Sci. 1964, 121, 404–427. [Google Scholar] [CrossRef]
- Fling, S. P.; Gregerson, D. S. Peptide and protein molecular weight determination by electrophoresis using a high-molarity Tris- buffer system without urea. Anal. Biochem. 1986, 155, 83–88. [Google Scholar] [CrossRef]
- Wu, W. U.; Hettiararchy, N. S.; Qui, M. Hydrophobicity, solubility, and emulsifying of soy protein prepared by papain modification and ultrafiltration. J.Am. Oil Chem. Soc. 1998, 75, 845–850. [Google Scholar] [CrossRef]
- Liu, M.; Markakis, P. An improved colorimetric method for determination antitryptic activity in soybean products. Cereal Chem. 1989, 66, 415–421. [Google Scholar]
- Thomson, D.B.; Erdman, J. W. Phytic acid determination in soybeans. J.Food Sci. 1982, 47, 513–519. [Google Scholar] [CrossRef]
- Velickovic, D.; Vucelic-Radovic, B.; Blagojevic, S.; Barac, M.; Stanojevic, S.; Ljubicic, M. A modification of a method for phytic acid determination. J. Serb. Chem. Soc 1999, 64, 303–310. [Google Scholar]
- Barac, M.; Stanojevic, s. The effect of microwave roasting on soybean protein composition and components with trypsin inhibitor activity. Acta Alimentaria 2005, 34, 23–31. [Google Scholar] [CrossRef]
- Marcone, M.F.; Kakuda, Y.; Yada, R.Y. Imunochemical examination of the surface physico-chemical properties of various dicotyledonous and monocotyledonous globulin seed storage proteins. Food Chem 1998, 63, 85–95. [Google Scholar] [CrossRef]
- Moreira, M. A.; Hermodson, M. A.; Larkins, B. A.; Nielsen, N.C. Comparison of the primary structure of the acidic polypeptides of glycinin. Arch. Biochem. Biophys. 1981, 210, 633–642. [Google Scholar] [CrossRef]
- Fukazawa, C.; Momma, T.; Hirano, H.; Harada, K.; Udaka, K. Glycinin A3B4 mRNA: cloning and sequencing of double stranded cDNA complementary to a soybean storage protein. J. Biol. Chem. 1985, 260, 6234–6239. [Google Scholar] [PubMed]
- Molina Ortiz, S.E.; Anon, M.C. Analysis of products, mechanisms of reaction, and some functional properties of soy protein hydrolysates. J. Am.Oil Chem. Soc 2000, 77, 1293–1302. [Google Scholar] [CrossRef]
- Kato, A.; Nakai, S. Hydrophobicity Determined by Fluorescence probe Method and Its Correlation with Surface Properties of Proteins. Biochim.Biophys.Acta 1980, 624, 13–20. [Google Scholar] [CrossRef]
- Nakai, S. Structure-Functional Relationship of Food Proteins with Emphasis on the Importance of the Protein Hydrophobicity. J.Agric.Food Chem. 1983, 31, 676–683. [Google Scholar] [CrossRef]
- Pesic, M.; Vucelic-Radovic, B.; Barac, M.; Stanojevic, S. The Influence of Genotypic Variation in Protein Composition on Emulsifying Properties of Soy Proteins. J.Am.Oil.Chem. 2005, 82(9), 667–672. [Google Scholar] [CrossRef]
- Sessa, D. J.; Wolf, W.J. Bowman-Birk inhibitors in soybean seed coats. Ind. Crops Prod 2001, 14, 73–83. [Google Scholar] [CrossRef]
- Thomson, L. Reduction of phytic acid concentration in protein isolates by acylation techniques. J. Am. Oil. Chem. Soc 1987, 64, 1712. [Google Scholar] [CrossRef]
- Wang, C.; Wixon, R. Phytochemicals in soybean – Their potential helth benefits. Inform 1999, 10, 315–320. [Google Scholar]
© 2006 by MDPI (http://www.mdpi.org). Reproduction is permitted for noncommercial purposes.
Share and Cite
MDPI and ACS Style
Barac, M.B.; Jovanovic, S.T.; Stanojevic, S.P.; Pesic, M.B. Effect of Limited Hydrolysis on Traditional Soy Protein Concentrate. Sensors 2006, 6, 1087-1101. https://doi.org/10.3390/s6091087
AMA Style
Barac MB, Jovanovic ST, Stanojevic SP, Pesic MB. Effect of Limited Hydrolysis on Traditional Soy Protein Concentrate. Sensors. 2006; 6(9):1087-1101. https://doi.org/10.3390/s6091087
Chicago/Turabian StyleBarac, Miroljub B., Snezana T. Jovanovic, Sladjana P. Stanojevic, and Mirjana B. Pesic. 2006. "Effect of Limited Hydrolysis on Traditional Soy Protein Concentrate" Sensors 6, no. 9: 1087-1101. https://doi.org/10.3390/s6091087