Airborne Laser Scanning Quantification of Disturbances from Hurricanes and Lightning Strikes to Mangrove Forests in Everglades National Park, USA

Abstract
:1. Introduction
2. Settings and Methods
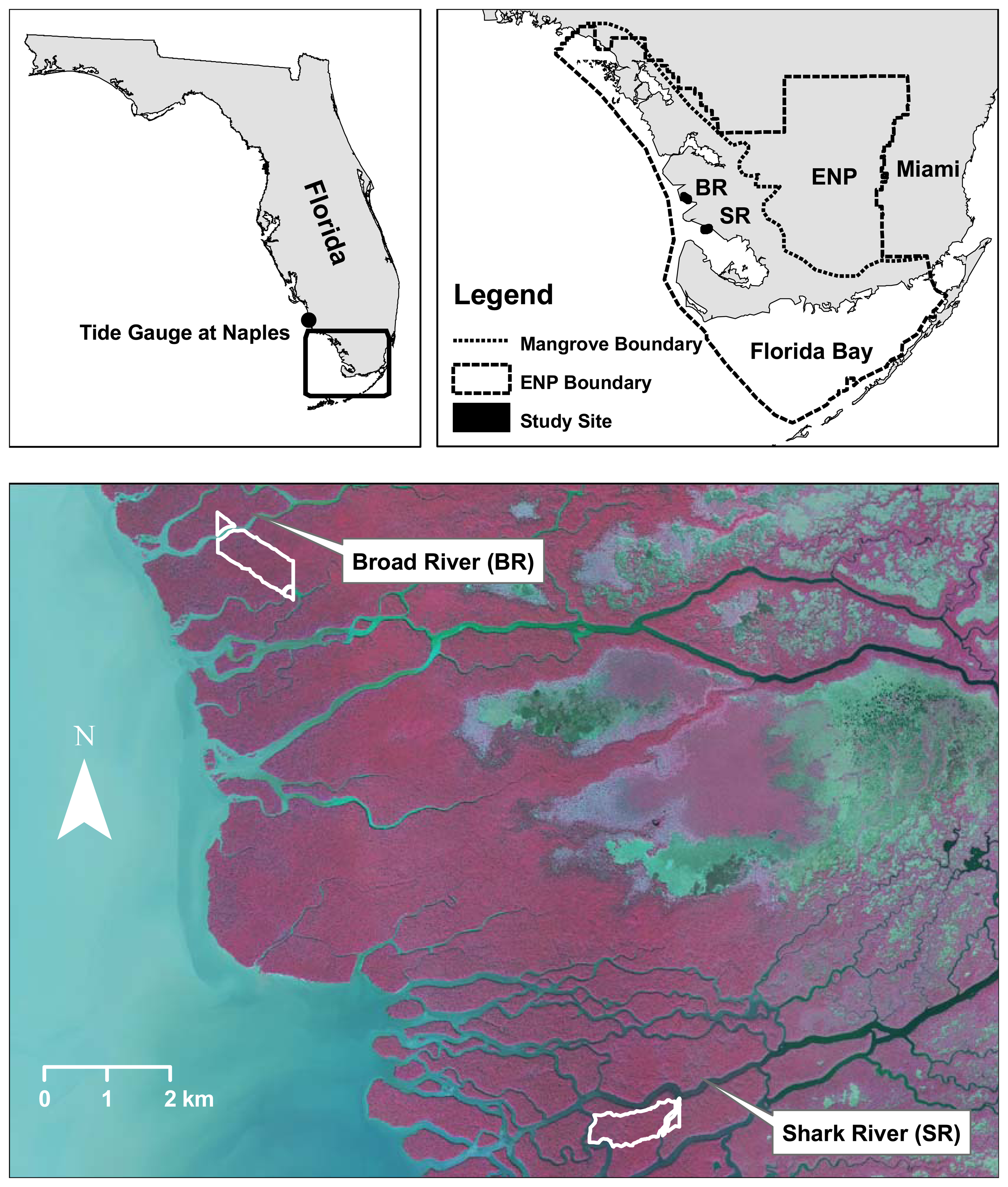
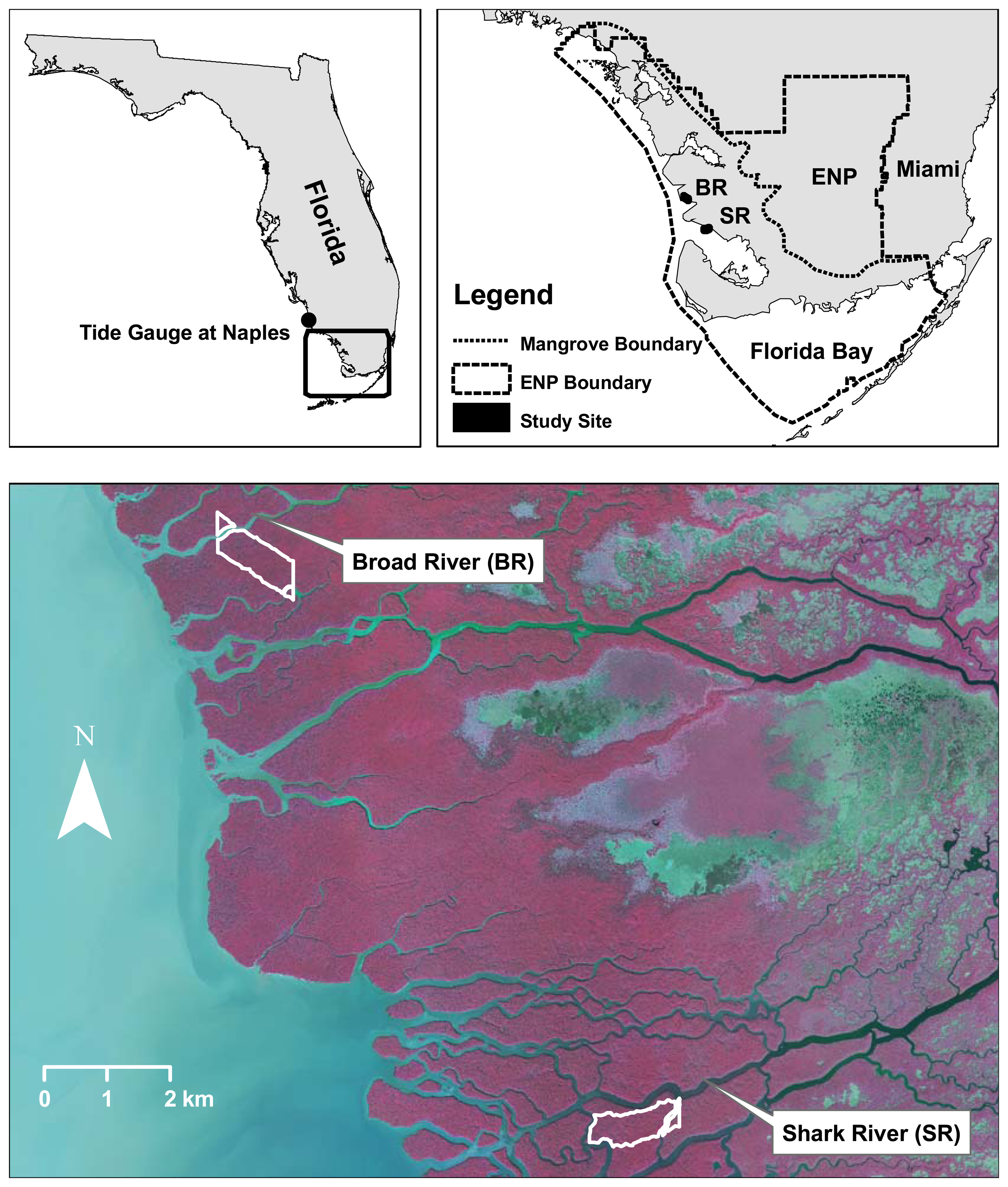
2.1. Mangrove forest in ENP
2.2. Hurricanes Katrina and Wilma
2.3. Historical hurricanes impacting the mangrove in ENP
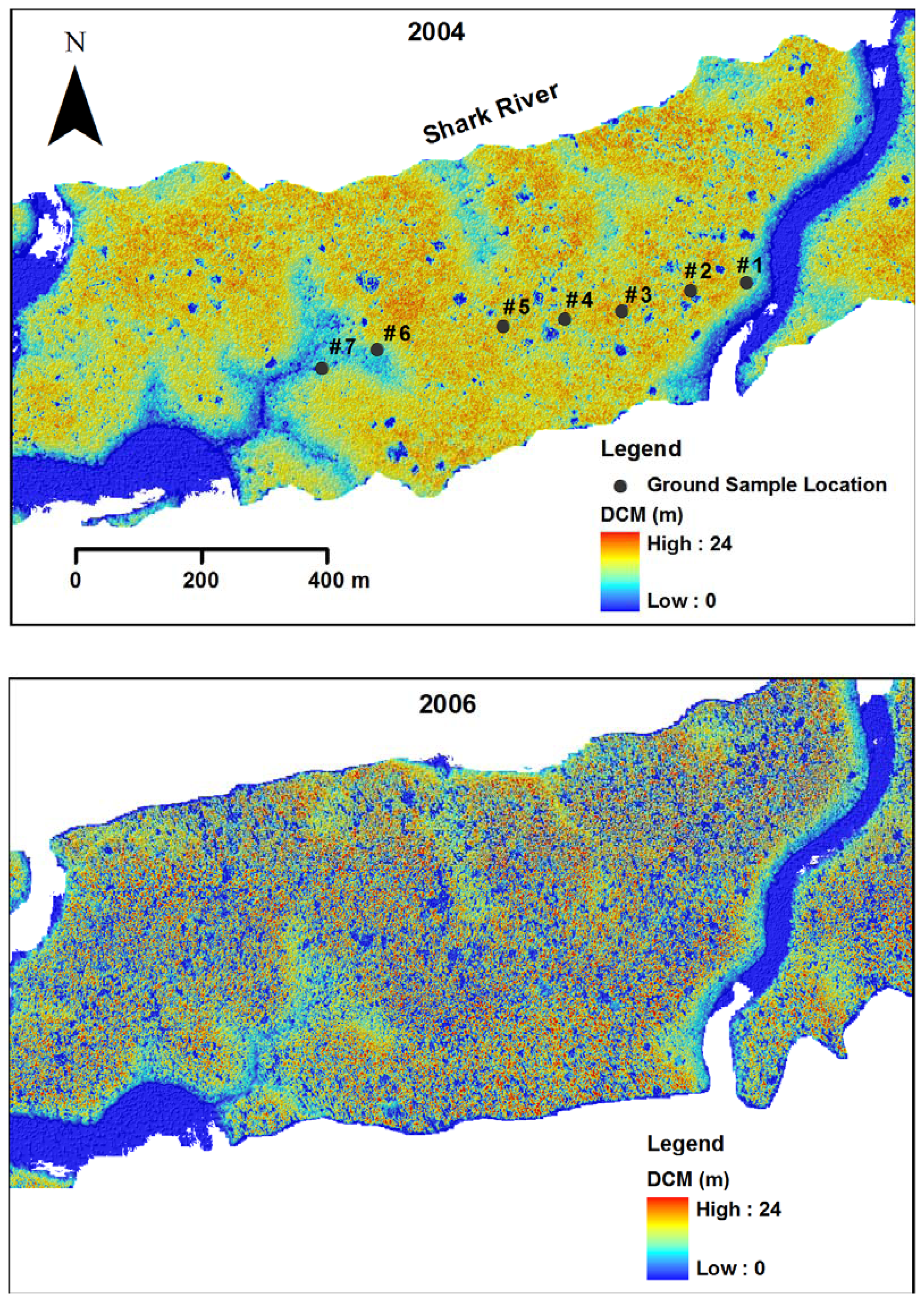
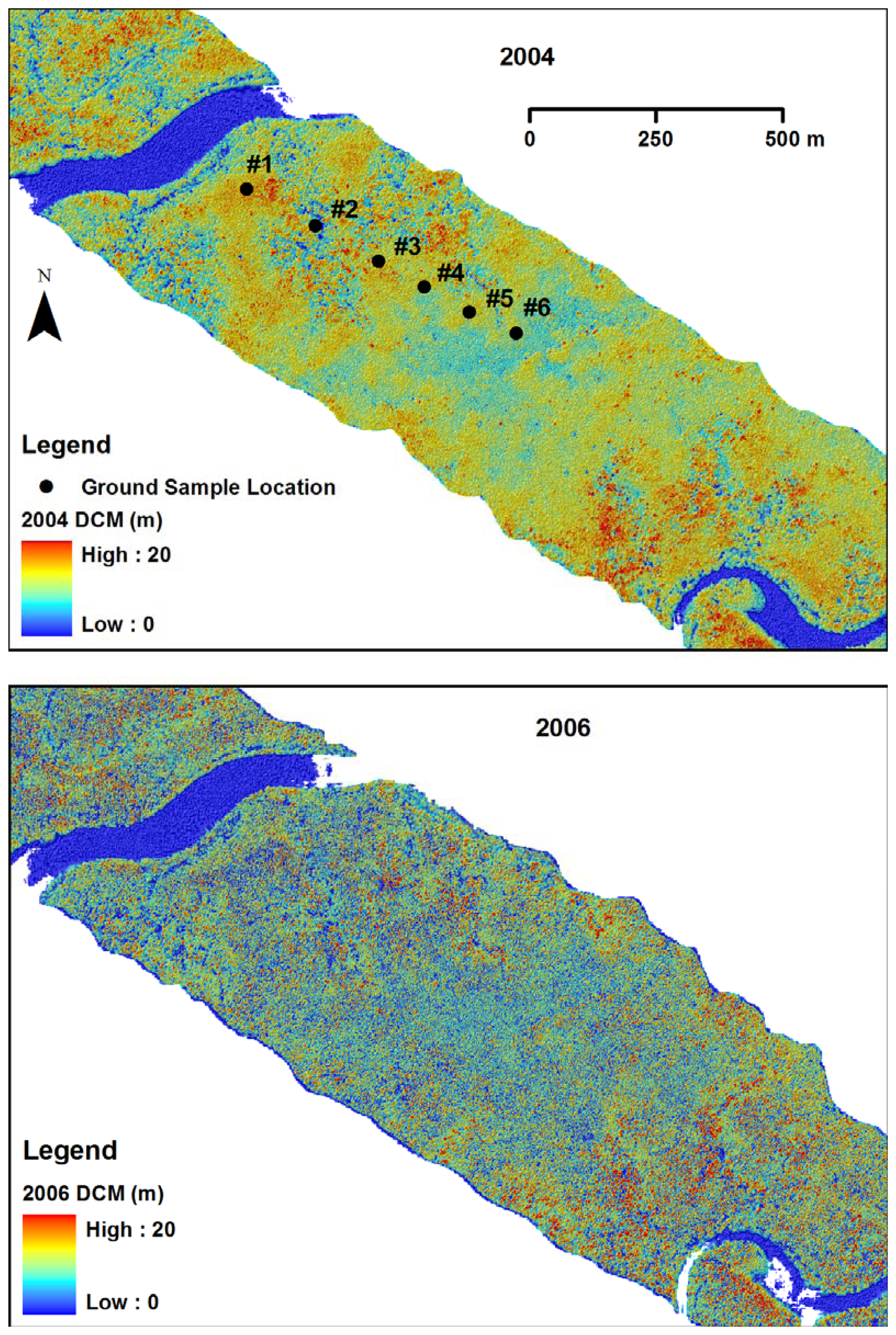
2.4. LIDAR data collection
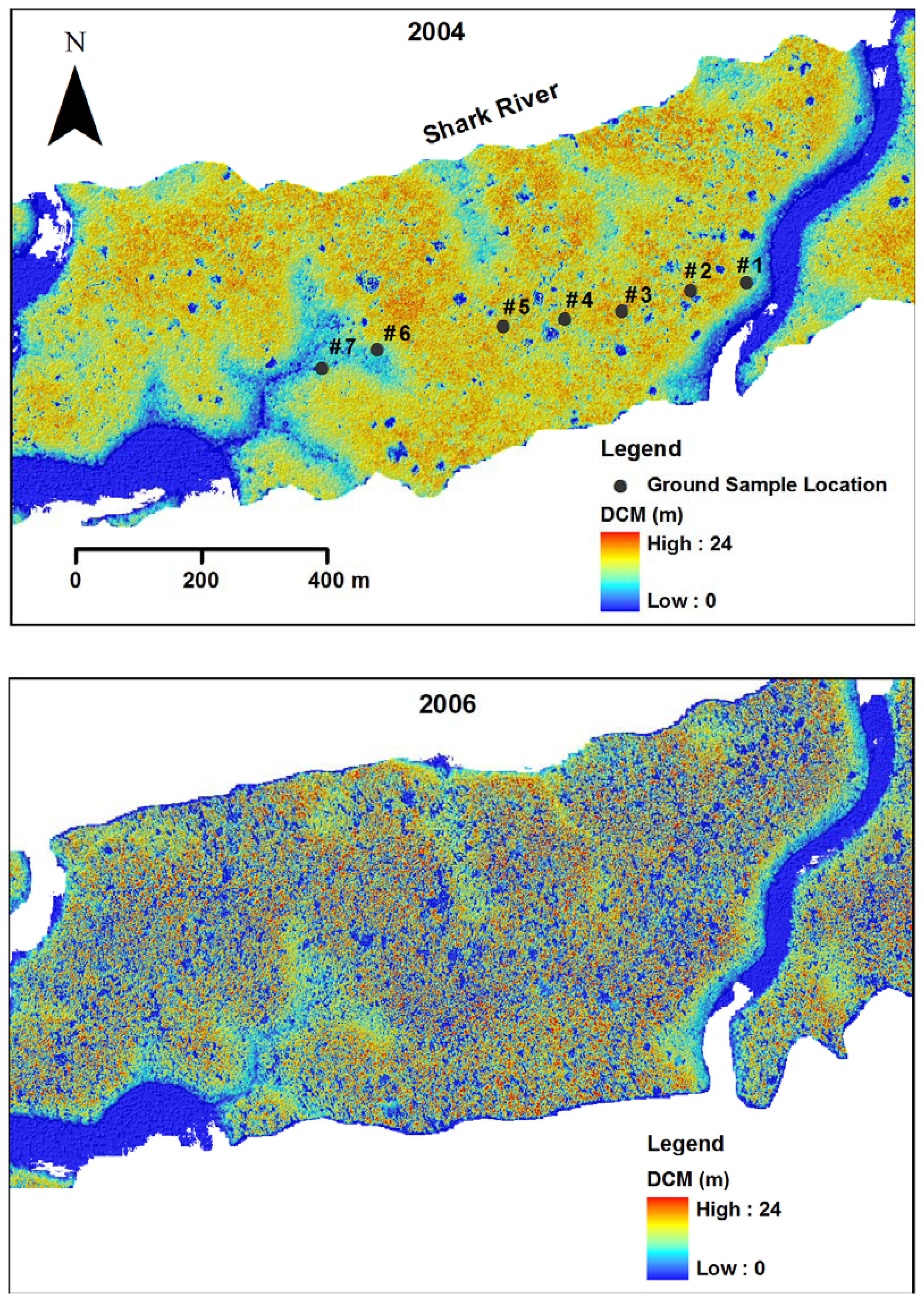
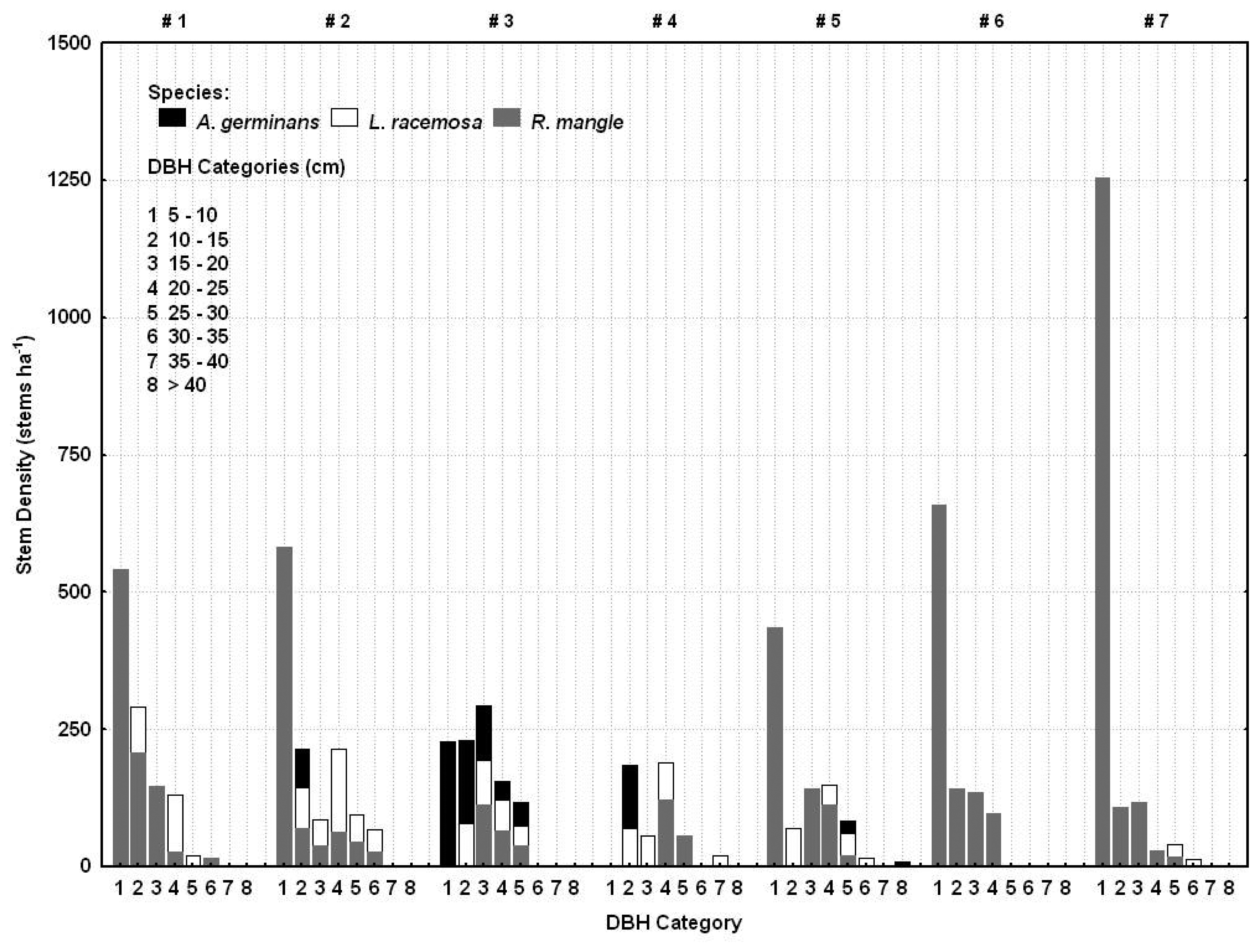
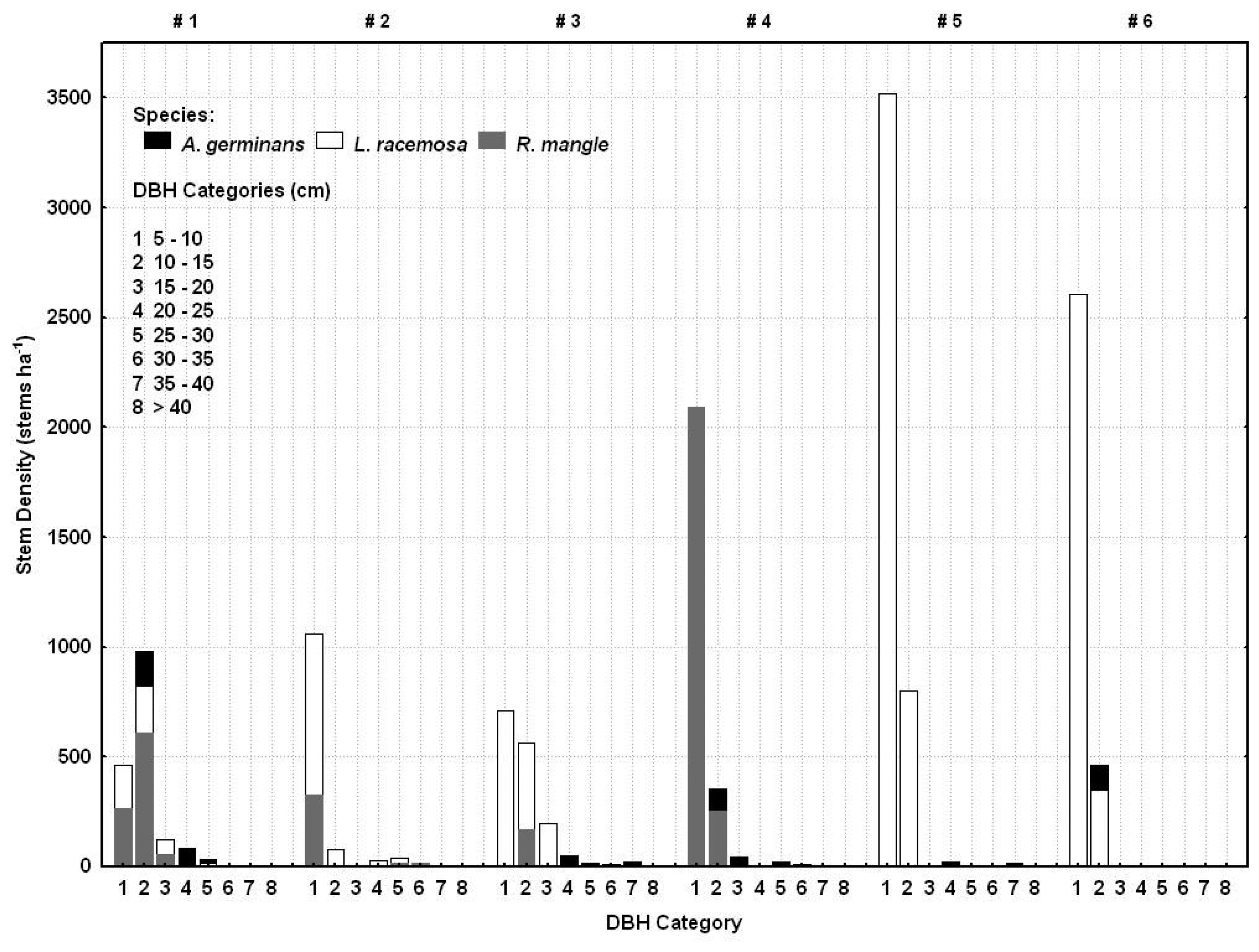
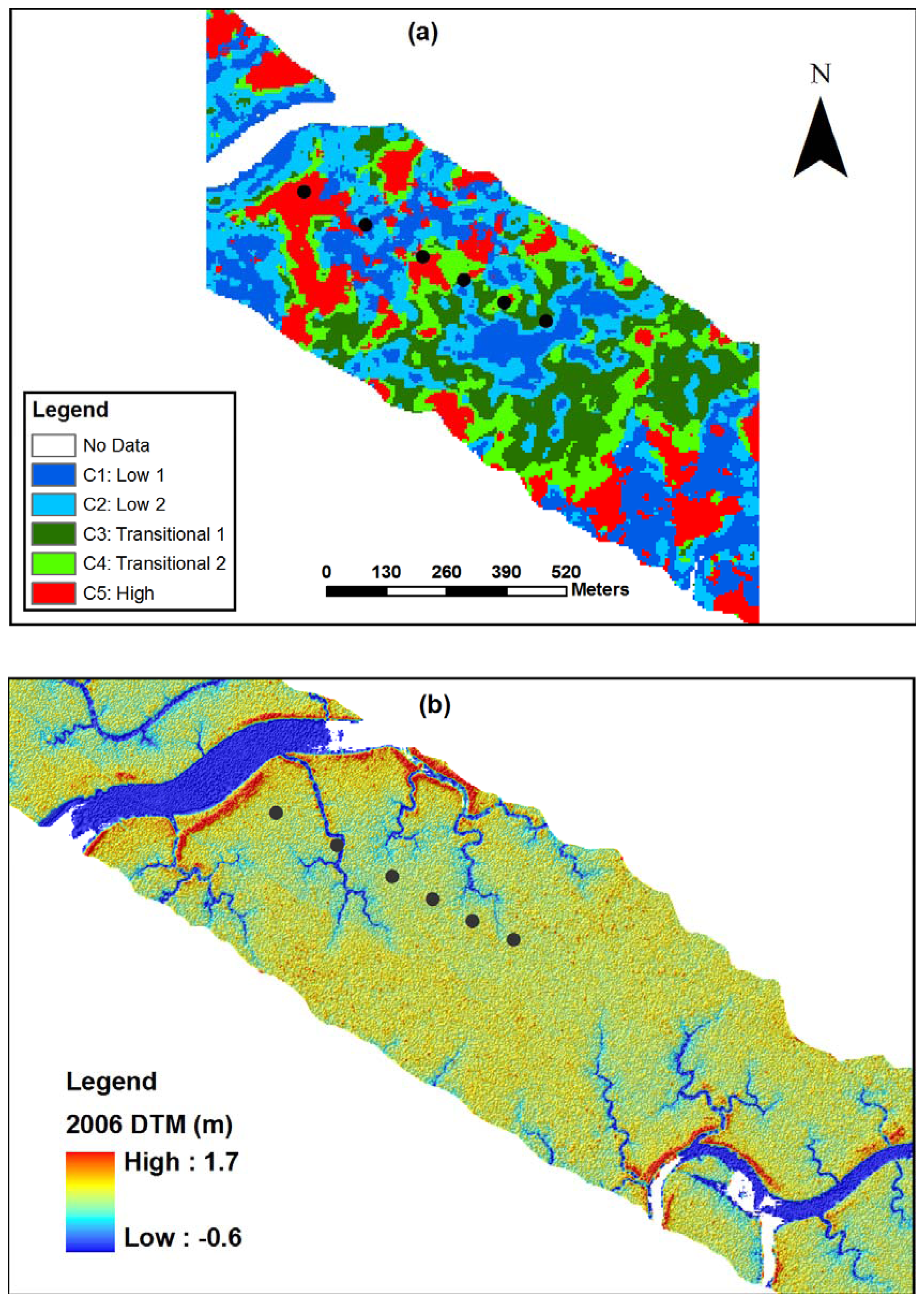
2.5. Ground survey of trees
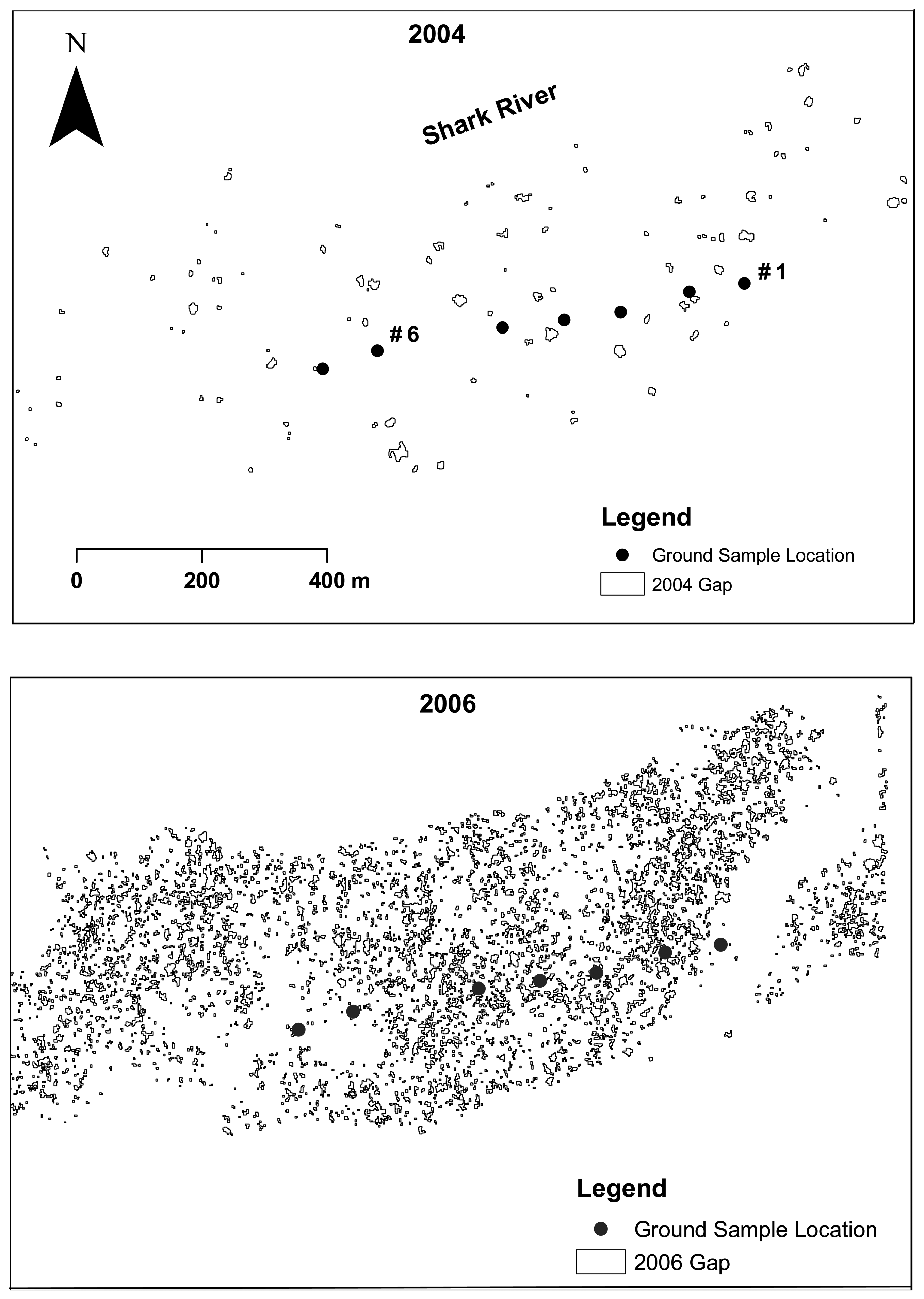
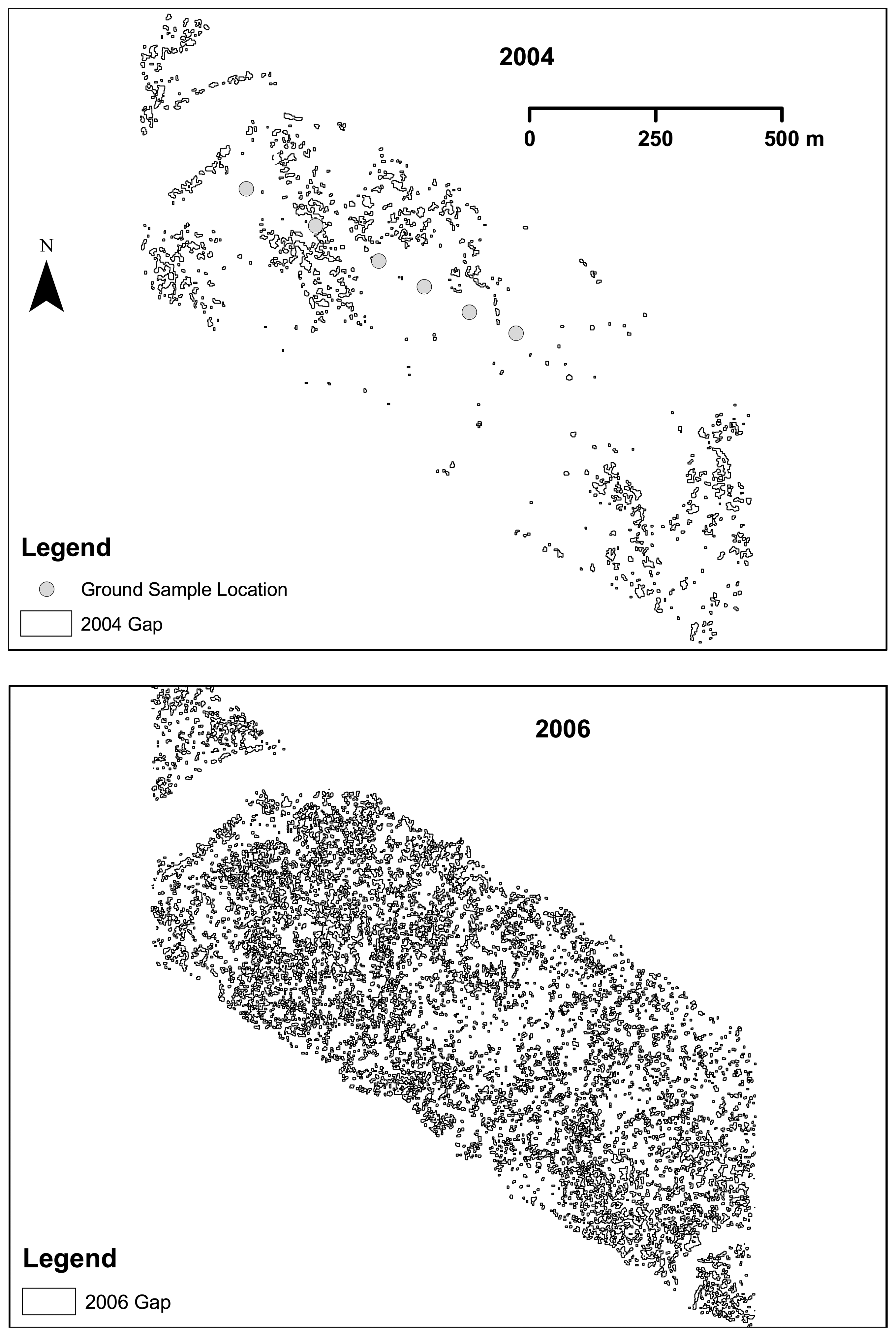
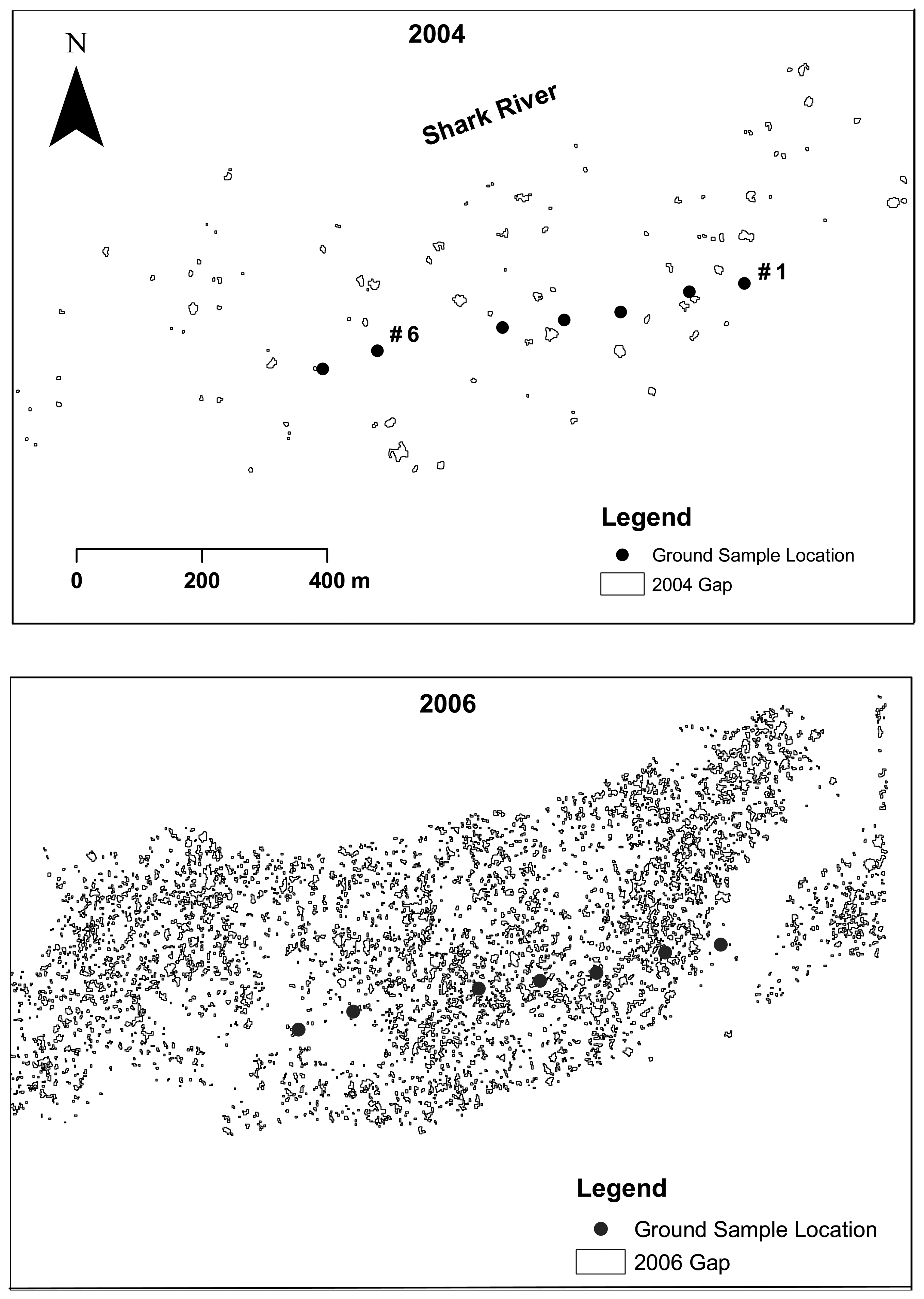
2.6. Gap and opening detection
2.7. Gap shape and size
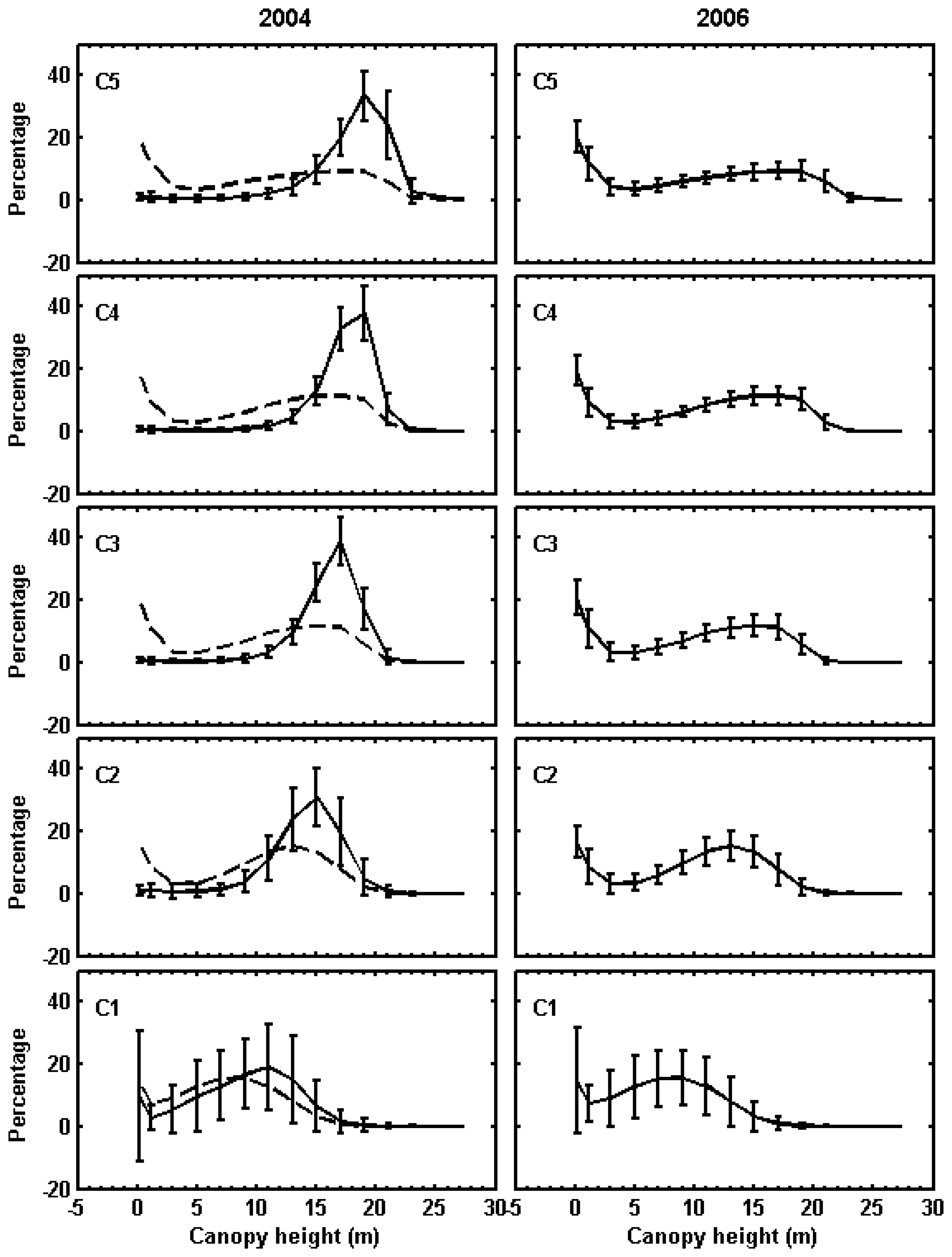
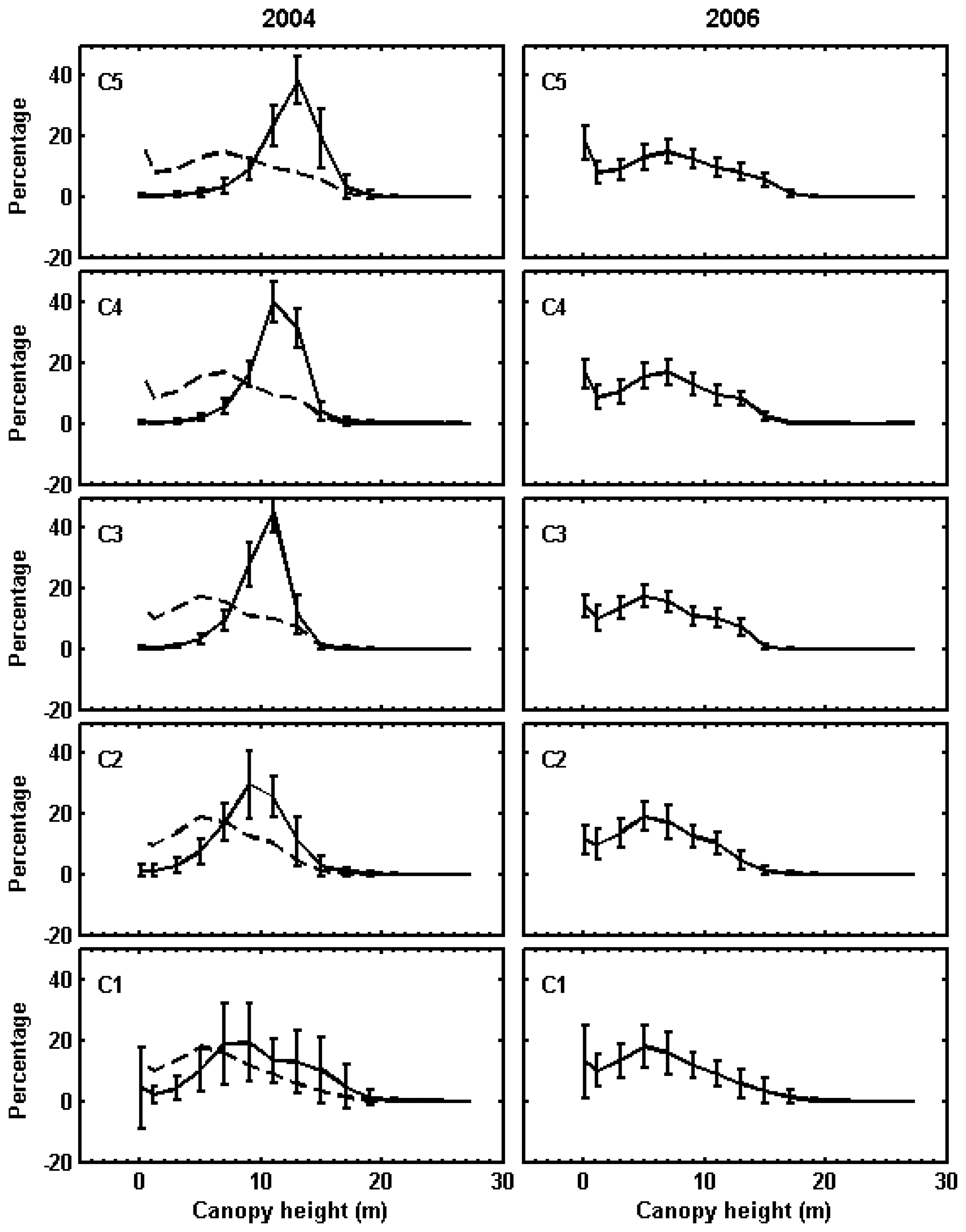
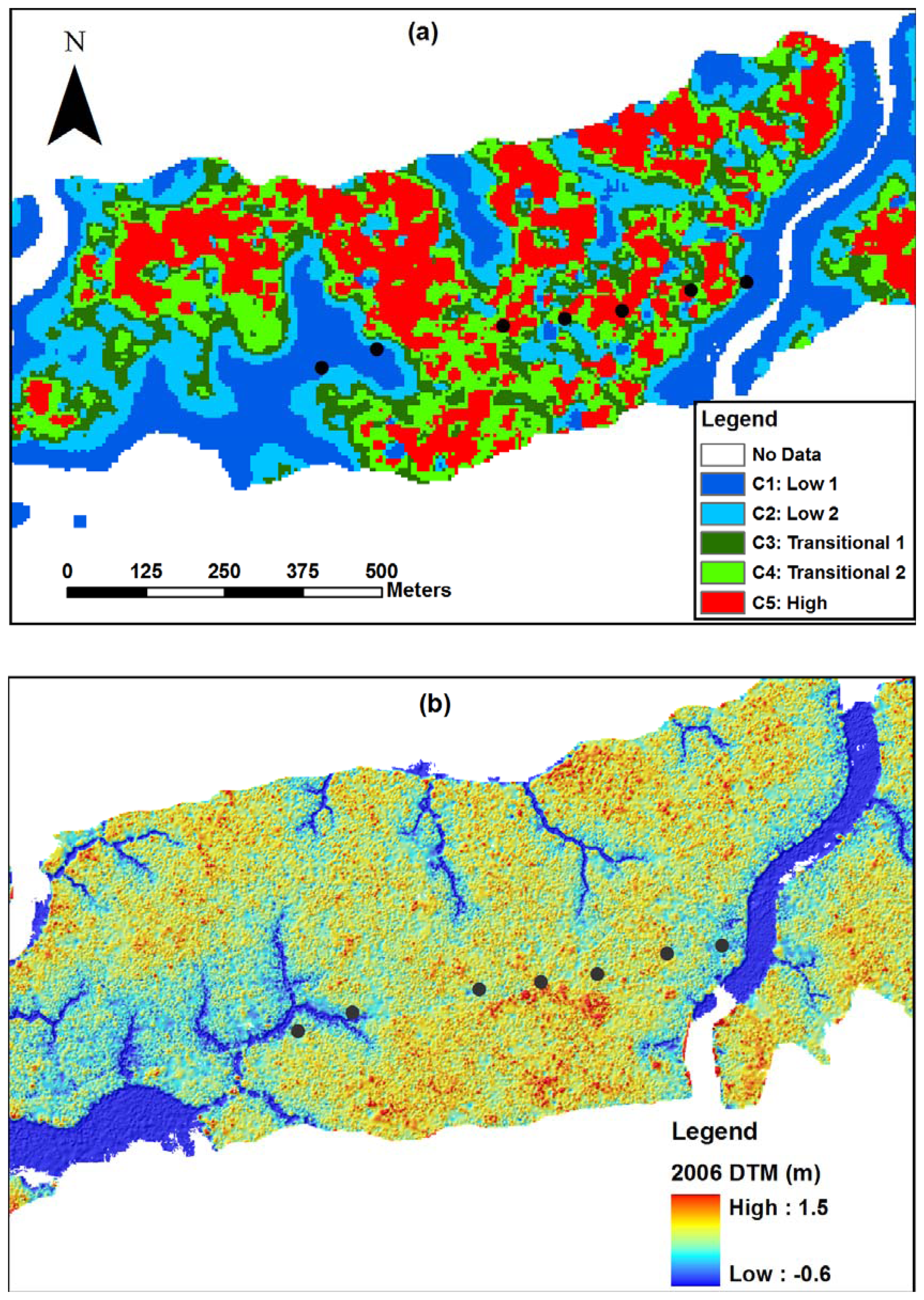
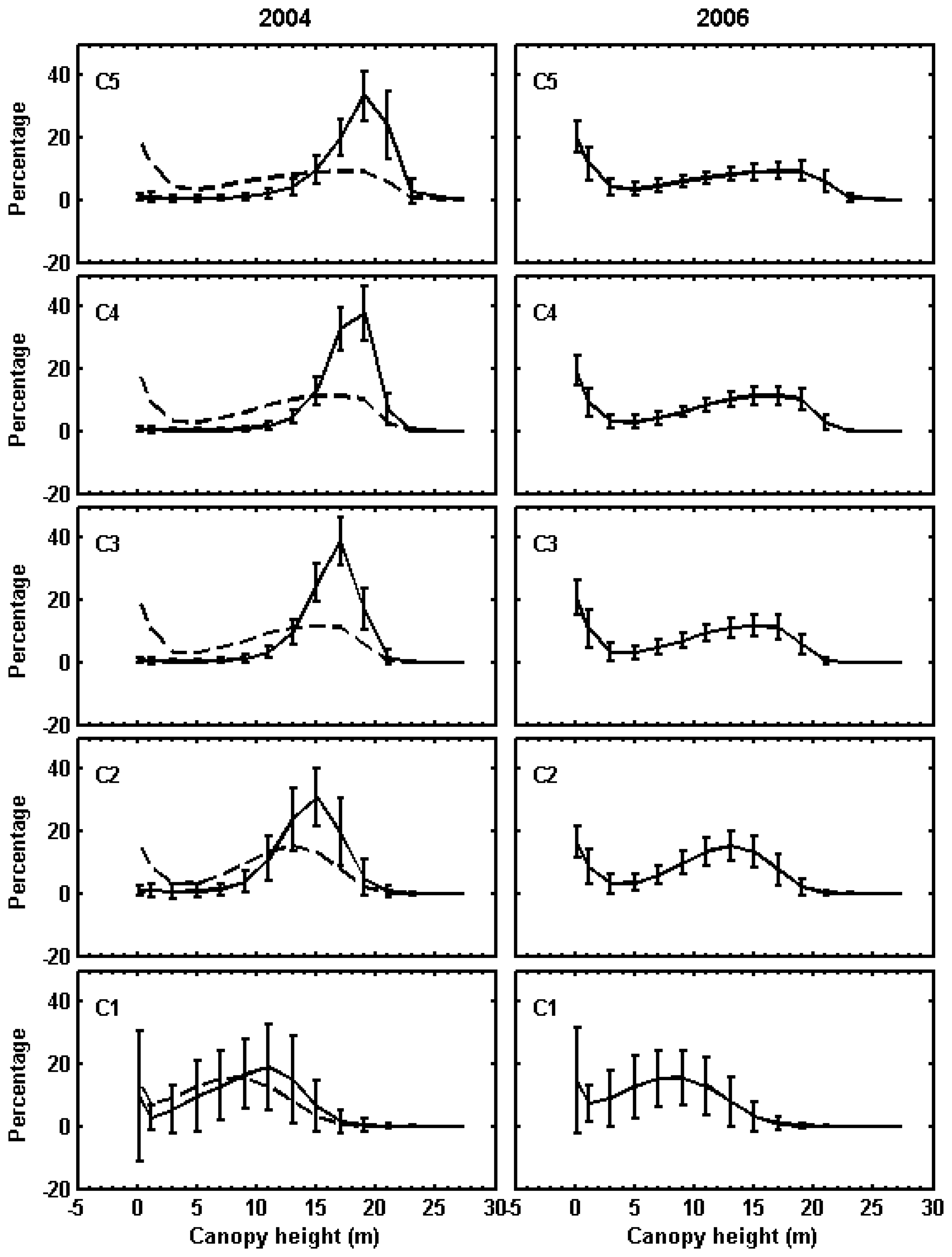
2.8. Classification of canopy changes
3. Results
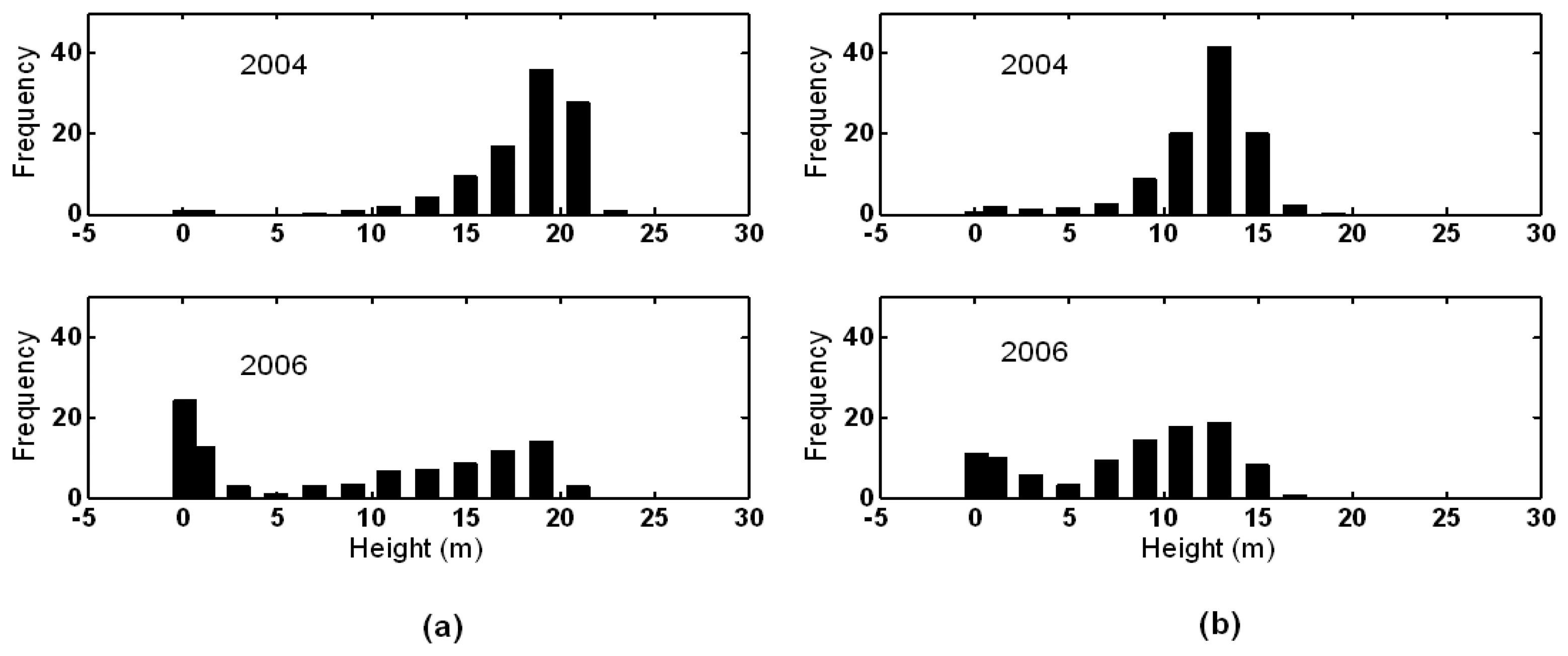
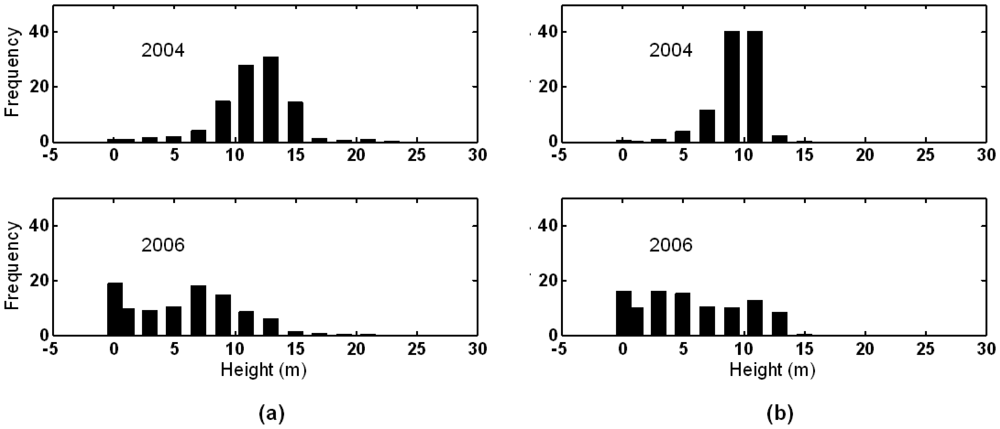
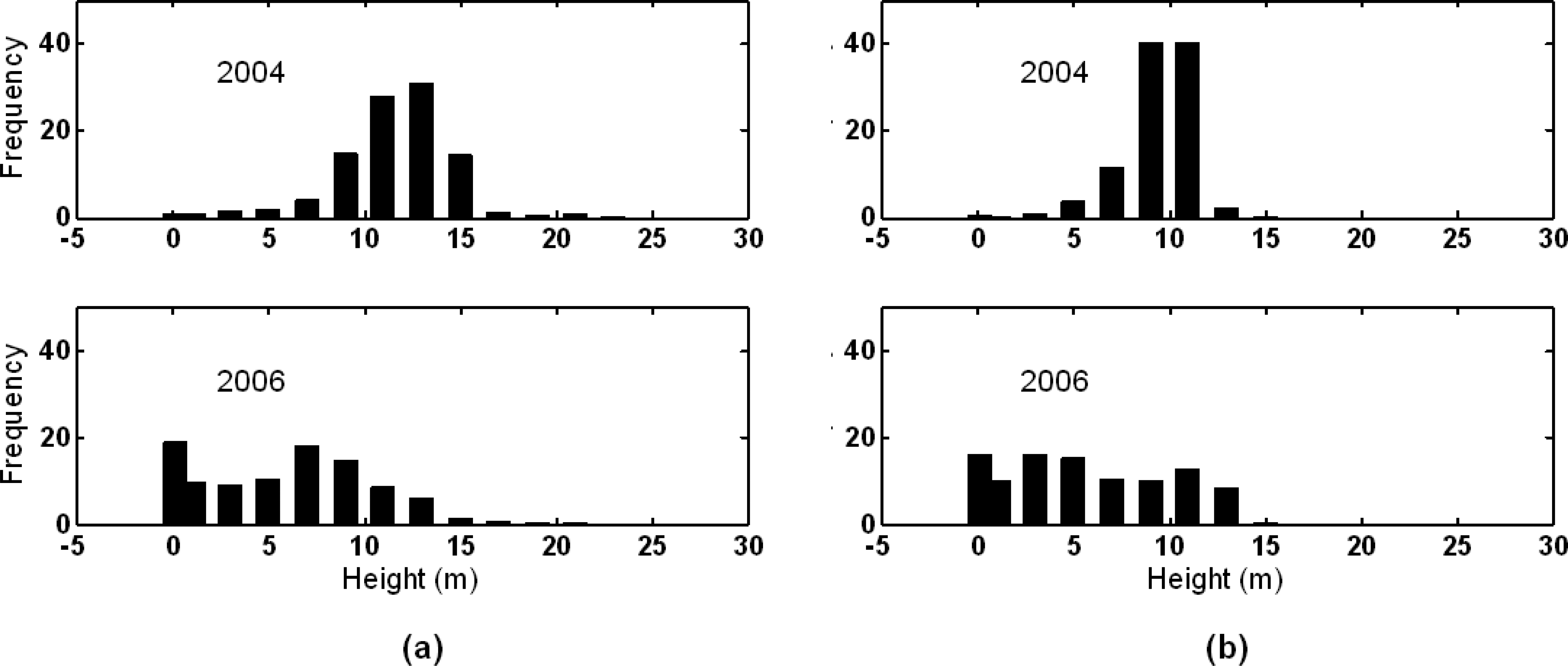
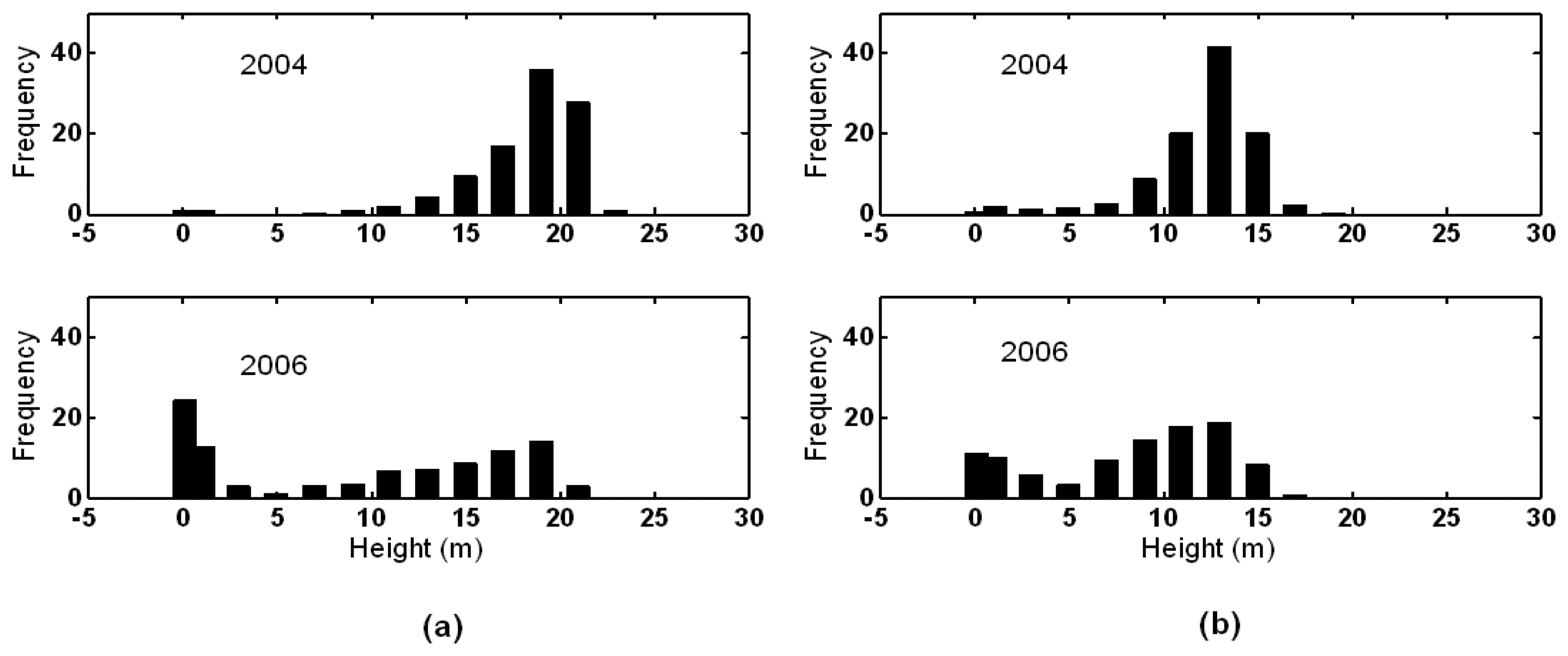
3.1. Shark River site
3.2. Broad River site
4. Discussion
5. Conclusion
Acknowledgments
References and Notes
- Feller, I.C.; McKee, K.L. Small gap creation in Belizean mangrove forests by wood-boring insect. Biotropica 1999, 31, 607–617. [Google Scholar]
- Lugo, A.E.; Patterson-Zucca, C. The impact of low temprature stress on mangrove structure and growth. Tropical Ecology 1977, 18, 149–161. [Google Scholar]
- Smith, T.J.I.; Robbles, M.B.; Wanless, H.R.; Doyle, T.W. Mangroves, hurricanes, and lightning strikes. Bioscience 1994, 44, 256–262. [Google Scholar]
- Armentano, T.V.; Doren, R.F.; Platt, W.J.; Mullins, T. Effects of Hurricane Andrew on coastal and interior forests of Southern Florida: overview and synthesis. Journal of Coastal Research 1995, 21, 111–144. [Google Scholar]
- Craighead, F.C.; Gilbert, V.C. The effects of Hurricane Donna on the vegetation of southern Florida. Quarterly Journal of the Florida Academy of Sciences 1962, 25, 1–28. [Google Scholar]
- Doyle, T.W.; Smith, T.J.I.; Robblee, M.B. Wind damage effect of Hurricane Andrew on mangrove communities along the southwest coast of Florida, USA. Journal of Coastal Research 1995, 12, 159–168. [Google Scholar]
- Piou, C.; Feller, I.C.; Berger, U.; Chi, F. Zonation patterns of Belizean offshore mangrove forests 41 years after a catastrophic hurricane. Biotropica 2006, 38, 365–374. [Google Scholar]
- Roth, L.C. Hurricanes and mangrove regeneration: effects of Hurricane Joan, October 1988, on the vegetation of Isla del Venado, Bluefields, Nicaragua. Biotropica 1992, 24, 375–384. [Google Scholar]
- Lugo, A.E.; Snedaker, S.C. The ecology of mangroves. Annual Review Ecology and Systematics 1974, 5, 39–64. [Google Scholar]
- Lugo, A.E. Mangrove ecosystems: successional or steady state? Biotropica 1980, 12, 65–72. [Google Scholar]
- Duke, N.C. Gap creation and regenerative processes driving diversity and structure of mangrove ecosystems. Wetlands Ecology and Management 2001, 9, 257–269. [Google Scholar]
- Sherman, R.E.; Fahey, T.J.; Battles, J.J. Small-scale disturbance and regeneration dynamics in a neotropical mangrove forest. Journal of Ecology 2000, 88, 165–178. [Google Scholar]
- Shafer, P.E.; Fuelberg, H.E. A statistical procedure to forecast warm season lightning over portions of the Florida Peninsula. Weather and Forecasting 2006, 21, 851–868. [Google Scholar]
- Whelan, K. The successional dynamics of lightning-initiated canopy gaps in the mangrove forests of Shark River, Everglades National Park, USA; 2005; Biology, Florida International University: Miami; p. 214. [Google Scholar]
- Baldwin, A.; Egnotovich, M.; Ford, M.; Platt, W. Regeneration in fringe mangrove forests damaged by Hurricane Andrew. Plant Ecology 2001, 157, 149–162. [Google Scholar]
- Boutet, J.C.; Weishampel, J.F. Spatial pattern analysis of pre- and post-hurricane forest canopy strcuture in North Carolina, USA. Landscape Ecology 2003, 18, 553–559. [Google Scholar]
- Weishampel, J.F.; Drake, J.B.; Cooper, A.; Blair, J.B.; Hofton, M. Forest canopy recovery from the 1938 hurricane and subsequent salvage damage measured with airborne LiDAR. Remote Sensing of Environment 2007, 109, 142–153. [Google Scholar]
- Simard, M.; Zhang, K.; Rivera-Monroy, V.H.; Ross, M.S.; Ruiz, P.L.; Castaneda-Moya, E.; Twilley, R.R.; Rodriguez, E. Mapping height and biomass of mangrove forests in Everglades National Park with SRTM elevation data. Photogrammetric Engineering and Remote Sensing 2006, 72, 299–311. [Google Scholar]
- Sklar, F.; McVoy, C.; VanZee, R.; Gawlik, D.E.; Tarboton, K.; Rudnick, D.; Miao, S. The effects of altered hydrology on the ecology of the Everglades. In The Everglades, Florida Bay, and Coral Reefs of the Florida Keys; Porter, J.W., Porter, K.G., Eds.; CRC Press: Boca Raton, Florida, 2002; pp. 39–82. [Google Scholar]
- Ross, M.S.; Meeder, J.F.; Sah, J.P.; Ruiz, P.L.; Telesnicki, G.J. The southeast saline Everglades revisited: 50 years of coastal vegetation change. Journal of Vegetation Science 2000, 11, 101–112. [Google Scholar]
- Knabb, R.D.; Rhome, J.R.; Brown, D.P. Hurricane Katrina 23- 20 August 2005; Tropical Cyclone Report. Miami, Florida, USA, 2005. http://www.nhc.noaa.gov.
- Powell, M.D.; Houston, S.H.; Amat, L.R.; Morisseau-Leroy, N. The HRD real-time hurricane wind analysis system. Journal of Wind Engineering Industrial Aerodynamics 1998, 77&78, 53–64. [Google Scholar]
- South Florida Weather Forecast Office. Hurricane Wilma; 2005; National Weather Service: Miami, Florida, USA. [Google Scholar]
- Willoughby, H.E.; Rahn, M.E. Parametric representation of the primary hurricane vortex. Part I: Observations and evaluations of the Holland (1980) model. Monthly Weather Review 2004, 132, 3033–3048. [Google Scholar]
- Bitterlich, W. Die Winkelzahlprobe. Allgemeine Forst-und Holzwirtschaftliche Zeitang 1948, 59, 4–5. [Google Scholar]
- Grosenbaugh, L. Plotless timber estimates – new, fast, and easy. Journal of Forestry 1952, 50, 32–37. [Google Scholar]
- Zhang, K. Identification of gaps in mangrove forests with airborne LIDAR. Remote Sensing of Environment 2008, in press. [Google Scholar]
- Zhang, K.; Chen, S.C.; Whitman, D.; Shyu, M.L.; Yan, J.; Zhang, C. A progressive morphological filter for removing non-ground measurements from airborne LIDAR data. IEEE Transactions on Geoscience and Remote Sensing 2003, 41, 872–882. [Google Scholar]
- Chen, R.; Twilley, R.R. A gap dynamic model of mangrove forest development along gradients of soil salinity and nutrient resources. Journal of Ecology 1998, 86, 37–51. [Google Scholar]
- Patton, D.R. A diversity index for quantifying habitat ‘edge’. Wildlife Society Bulletin 1975, 3, 171–173. [Google Scholar]
- Zhang, K.; Houle, P.A.; Ross, M.S.; Ruiz, P.L.; Simard, M. Airborne laser mapping of mangroves on the Biscayne Bay coast, Miami, Florida. Proceedings of 2006 IEEE International Geoscience and Remote Sensing Symposium & 27th Canadian Symposium on Remote Sensing, Denver, Colorado, USA; 2006; pp. 3750–3754. [Google Scholar]
- Jensen, J.R. Introductory Digital Image Processing, Third Edition ed; Prentice Hall: New Jersey, USA, 2004; p. 544. [Google Scholar]
- Craighead, F.C. The trees of South Florida, vol. 1. The natural environments and their succession; University of Miami Press: Miami, Florida, USA, 1971; p. 212. [Google Scholar]
- Goldenberg, S.B.; Landsea, C.W.; Mestas-Nuñez, A.M.; Gray, W.M. The recent increase in Atlantic hurricane activity: causes and implications. Science 2001, 293, 474–479. [Google Scholar]
- Clarke, P.J.; Kerrigan, R.A. Do forest gaps influence the population structure and species composition of mangrove stands in Northern Australia? Biotropica 2000, 32, 642–652. [Google Scholar]
- Ross, M.S.; Ruiz, P.L.; Sah, J.P.; Reed, D.L.; Walters, J.; Meeder, J.F. Early stand development following disturbance along a mangrove forest productivity gradient in South Florida. Plant Ecology 2006, 185, 283–297. [Google Scholar]
- McKee, K.L. Interspecific variation in growth, biomass partition, and defensive characteristics of neotropical mangrove seedlings: response to light and nutrient availability. American Journal of Botany 1995, 82, 299–307. [Google Scholar]
- Cardona-Olarte, P.; Twilley, R.R.; Krauss, K.W.; Rivera-Monroy, V. Responses of neotropical mangrove seedlings grown in monoculture and mixed culture under treatments of hydroperiod and salinity. Hydrobiologia 2006, 569, 325–341. [Google Scholar]
- Krauss, K.W.; Doyle, T.W.; Twilley, R.R.; Smith, T.J.I.; Whelan, K.R.T.; Sullivan, J.K. Woody debris in the mangrove forests of South Florida. Biotropica 2005, 9–15. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | 1 | 2 | 3 | 4 | 5 |
|---|---|---|---|---|---|
| Vmax (m/s) | 37.6 | 45.8 | 53.8 | 64.0 | 69.5 |
| Rmax (km) | 40 | 35 | 31 | 26 | 24 |
| Name | 2004 | 2006 |
|---|---|---|
| Cell size (c) of DCM | 1 (m) | 1 (m) |
| Number of windows for ASF (N) | 8 | 8 |
| Increment of window size (s) | 2 (c) | 2 (c) |
| Gap depth threshold (λ) | 0.60 | 0.60 |
| Minimum canopy height (hmin) | 5 (m) | 5 (m) |
| Minimum gap area (amin) | 10 (m2) | 10 (m2) |
| Window size for filling holes (wf) | 3 | NA |
| Site | Gap Number | Gap Number per km2 | Gap Area (km2) | Gap Area / Total Area (%) | Total Forest Area (km2) |
|---|---|---|---|---|---|
| Shark River | 311/2661 | 510/4279 | 0.007/0.074 | 1.1/12.1 | 0.610 |
| Broad River | 290/2527 | 424/3694 | 0.013/0.083 | 1.9/12.2 | 0.684 |
| PCAs | 1st PCA | 2nd PCA | 3rd PCA | 4th PCA | 5th PCA | Total |
|---|---|---|---|---|---|---|
| Shark River | 44.48% | 21.90% | 10.95% | 6.28% | 5.19% | 88.80% |
| Broad River | 43.47% | 29.26% | 9.36% | 5.54% | 4.76% | 92.39% |
© 2008 by MDPI (http://www.mdpi.org). Reproduction is permitted for noncommercial purposes.
Share and Cite
Zhang, K.; Simard, M.; Ross, M.; Rivera-Monroy, V.H.; Houle, P.; Ruiz, P.; Twilley, R.R.; Whelan, K. Airborne Laser Scanning Quantification of Disturbances from Hurricanes and Lightning Strikes to Mangrove Forests in Everglades National Park, USA. Sensors 2008, 8, 2262-2292. https://doi.org/10.3390/s8042262
Zhang K, Simard M, Ross M, Rivera-Monroy VH, Houle P, Ruiz P, Twilley RR, Whelan K. Airborne Laser Scanning Quantification of Disturbances from Hurricanes and Lightning Strikes to Mangrove Forests in Everglades National Park, USA. Sensors. 2008; 8(4):2262-2292. https://doi.org/10.3390/s8042262
Chicago/Turabian StyleZhang, Keqi, Marc Simard, Michael Ross, Victor H. Rivera-Monroy, Patricia Houle, Pablo Ruiz, Robert R. Twilley, and Kevin Whelan. 2008. "Airborne Laser Scanning Quantification of Disturbances from Hurricanes and Lightning Strikes to Mangrove Forests in Everglades National Park, USA" Sensors 8, no. 4: 2262-2292. https://doi.org/10.3390/s8042262