
Genome-Wide Analysis of VILLIN Gene Family Associated with Stress Responses in Cotton (Gossypium spp.)



Amity Institute of Biotechnology, Amity University Jharkhand, Ranchi 835303, India
*
Author to whom correspondence should be addressed.
Curr. Issues Mol. Biol. 2024, 46(3), 2278-2300; https://doi.org/10.3390/cimb46030146
Submission received: 1 February 2024
/
Revised: 3 March 2024
/
Accepted: 8 March 2024
/
Published: 11 March 2024
(This article belongs to the Section Molecular Plant Sciences)
Abstract
:The VILLIN (VLN) protein plays a crucial role in regulating the actin cytoskeleton, which is involved in numerous developmental processes, and is crucial for plant responses to both biotic and abiotic factors. Although various plants have been studied to understand the VLN gene family and its potential functions, there has been limited exploration of VLN genes in Gossypium and fiber crops. In the present study, we characterized 94 VLNs from Gossypium species and 101 VLNs from related higher plants such as Oryza sativa and Zea mays and some fungal, algal, and animal species. By combining these VLN sequences with other Gossypium spp., we classified the VLN gene family into three distinct groups, based on their phylogenetic relationships. A more in-depth examination of Gossypium hirsutum VLNs revealed that 14 GhVLNs were distributed across 12 of the 26 chromosomes. These genes exhibit specific structures and protein motifs corresponding to their respective groups. GhVLN promoters are enriched with cis-elements related to abiotic stress responses, hormonal signals, and developmental processes. Notably, a significant number of cis-elements were associated with the light responses. Additionally, our analysis of gene-expression patterns indicated that most GhVLNs were expressed in various tissues, with certain members exhibiting particularly high expression levels in sepals, stems, and tori, as well as in stress responses. The present study potentially provides fundamental insights into the VLN gene family and could serve as a valuable reference for further elucidating the diverse functions of VLN genes in cotton.
1. Introduction
In plant cells, the actin cytoskeleton is a complex and dynamic network that actively participates in several crucial activities, including signal transduction, vesicle trafficking, cell expansion, cell division, organelle movement, stomatal opening, and cytoplasmic streaming [1]. The diverse gelsolin/villin/fragmin superfamily, as well as nucleating proteins such as Formin and Arp2/3, monomer-binding protein profilin, severing/depolymerizing proteins ADF, and cofilin, are just a few actin-binding proteins (ABPs) that regulate cellular and cytoskeletal dynamics [2,3]. At certain time points and sites, ABPs control the assembly and disassembly of monomeric (G-actin) and filamentous actin (F-actin). Functional investigation of several ABPs has been performed in Arabidopsis. For instance, the loss of Actin-Depolymerizing Factor 5 (AtADF5) function has been shown to decrease drought tolerance and disrupt stomata closure due to abnormal actin dynamics [4]. The knockdown of FORMIN3 (AtFH3) via RNA interference has been observed to reduce actin-cable abundance and affects pollen-tube growth [5]. Profilins (PRFs) are cytosolic proteins consisting of 129–133 amino acids with a molecular weight of 12–15 kDa and play a crucial role in regulating the cell cytoskeleton architecture, primarily through the modulation of actin polymerization [6,7,8]. These actin-binding proteins possess conserved profilin–actin interacting regions (PAINRs), which are essential for actin polymerization or depolymerization processes [7,8]. Recent studies have investigated their functional roles in root elongation, leaf morphology, epidermal expansion, flowering time, and seed germination across various plant species [9,10,11]. Fimbrin, another well-known ABP, also plays a role in the regulation of actin dynamics. In the atfim4/atfim5 double mutant, there were significant changes in the morphology and length of root hair [12]. Collectively, these studies indicated that ABPs are crucial for plant development and exert their effects by regulating actin dynamics.
VLN, a member of the villin/gelsolin/fragmin actin-binding protein (ABP) superfamily, is a key regulator of both actin stability and dynamics [5,13,14,15]. The regulatory functions of the VLN proteins are intricately linked to their structural characteristics. Six gelsolin home domains (G1-G6) at the N-terminus and a headpiece domain (VHP) at the C-terminus, each containing two conserved actin-binding domains, distinguish VLN proteins from other actin-binding domains (ABDs). ABD2 in the VHP area bundles actin filaments together, whereas ABD1, which spans G1 and G2, binds to two neighboring actin monomers inside the filament [16,17,18]. Additionally, VLN proteins possess calcium ion (Ca2+)-binding sites, which vary in type and number among different VLN variants. VLNs can bind to F-actin either independently or in a Ca2+/calmodulin-dependent manner. These features allow VLN proteins to alter the dynamics and organization of actin filaments, which helps create highly fibrillar actin structures [19,20,21,22]. In plant biology, the first functionally characterized VLN homologs were identified in Lilium brownii and were named 115-ABP and 135-ABP. These proteins bind to the F-actin [23,24,25]. Further investigations have revealed that 135-ABP plays a critical role in regulating the arrangement of F-actin in pollen tubes as well as the cytoplasmic architecture and actin-filament organization in root hairs [19,25]. There are five VLN genes in Arabidopsis, which is a model plant species. Although these genes are widely expressed in various plant tissues [26,27,28,29], only a subset of them has been functionally characterized. Specifically, the loss of AtVLN1 and AtVLN4 function has been associated with longer and shorter root hairs, respectively, indicating their distinct roles in the regulation of root-hair growth in Arabidopsis [30]. AtVLN2 and AtVLN3 have been implicated in normal plant development and organ morphogenesis [31,32], while AtVLN5 is essential for pollen-tube growth [33]. Collectively, these studies underscore the vital role of VLNs in plant development through the regulation of various aspects of actin cytoskeleton and actin dynamics. To date, there has been limited research on VLN proteins in the Gossypium species. Several other actin-binding proteins (ABPs) and related proteins that may play a role in cytoskeletal dynamics and stress response activities have been investigated in Gossypium species, such as profilin, LIM, and DUF668 [3,34,35].
In the present study, 14 VLN genes were identified in the Gossypium hirsutum genome. Subsequent analyses encompassed the examination of gene structures, conserved domains, phylogenetic relationships, chromosomal locations, cis-regulatory elements, gene synteny, and expression patterns. The present study provides comprehensive and valuable systematic insights that might be useful for the extended exploration of the functional aspects of GhVLN in cotton.
2. Materials and Methods
2.1. Sequence Retrieval and Identification of Villin Gene Family
The VLN sequence was retrieved from Arabidopsis thaliana using the NCBI Protein Database (https://www.ncbi.nlm.nih.gov/protein) (accessed on 9 September 2023) [36] with the accession number BAA96955.1, as the sequence structure was analyzed by Sato et al. in the year 2000 [37]. To identify all the possible VLN Gossypium hirsutum, we used a BLAST model consisting of six gelsolins and one VHP of Arabidopsis thaliana in the Gossypium hirsutum protein database from the Phytozome 13 Web BLAST Search (Gossypium hirsutum v2.1, https://phytozome-next.jgi.doe.gov/) (accessed on 13 September 2023) [38], with the target type set as Proteome, Program—BLASTP. The Expect threshold was configured as (−1), and a total of 100 set alignments were specified. We retrieved 14 protein sequences, identified by their protein IDs, as hits. To predict the conserved domains within GhVLN, we conducted a Batch Conserved Domain search using the NCBI platform (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi) (accessed on 16 September 2023) [39]. This process also helped to eliminate redundant sequences, and we used the default settings throughout. We used the SMART tool (http://smart.embl-heidelberg.de/) (accessed on 13 September 2023) [40] to confirm the existence of both gelsolin and VHP domains.
2.2. Analysis of Protein Physiochemical Properties, Amino Acid Alignment, and Phylogenetic Investigation
We utilized the ExPASy online platform (https://web.expasy.org/protparam/) (accessed on 13 September 2023) [41] to estimate a range of physicochemical characteristics of Gossypium hirsutum VLN, such as chromosome number, molecular weight (MW), isoelectric point (pI), protein length (measured in amino acid count), and Grand Average of hydropathy (GRAVY). BaCelLo (https://busca.biocomp.unibo.it/bacello/) (accessed on 13 September 2023) [42] tool was used to predict the subcellular locations of the proteins. Subsequently, a subcellular localization heatmap was generated using WoLF PSORT (https://wolfpsort.hgc.jp/) (accessed on 16 September 2023) [43]. Amino acid alignment was performed using DNAMAN Version 10 (Lynnon Biosoft, 2018). Phylogenetic tree was generated by aligning sequences and applying the neighbor-joining (NJ) method within MEGA11 software [44]. This analysis incorporated 1000 bootstrap replicates and employed the Jones–Taylor–Thornton (JTT) model. The tree was further modified using Interactive Tree Of Life (iTOL) Version 6.8.1 (https://itol.embl.de/) (accessed on 10 November 2023) [45].
2.3. Analysis of Gene Structure, Identification of Conserved Motifs, and Conducting Conserved Domain Analysis
We obtained the gene sequences of the Gossypium hirsutum VLN family protein from the Phytozome website (Gossypium hirsutum v2.1, https://phytozome-next.jgi.doe.gov/) (accessed on 13 September 2023) [38], and these sequences were extracted using TBtools (version 2.003). To illustrate gene structures, we utilized GFF3 files. Additionally, online tool GSDS 2.0 (http://gsds.cbi.pku.edu.cn/) (accessed on 16 September 2023) [46] was employed to visually represent the exon/intron structures of the GhVLN genes. NCBI Batch Conserved Domain search (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi) (accessed on 16 September 2023) [39] was used to predict conserved domains. For motif analysis, we used the MEME Suite (https://meme-suite.org/meme/tools/meme) (accessed on 24 September 2023) [47] to identify 20 motifs. All data were visualized using TBtools.
2.4. Collinearity and Chromosomal Location Analysis of GhVLN Genes
We obtained complete genome data for Gossypium hirsutum and extracted Arabidopsis thaliana genome sequences based on their transcript IDs using the FASTA Extract tool in TBtools. A collinearity relationship was established between these two species using the One-Step MCScanX feature of TBtools, with BlastP configured with eight CPU threads, an E-value of 1 × 10−10, and a minimum of five blast hits. Subsequently, we combined the CTL, GFF, and collinearity files using the Dual Synteny Plot within MCScanX in TBtools. All the chromosome scaffolds were manually removed from the CTL file before using the Dual Synteny Plot. Finally, we constructed and visualized the collinear relationship between Gossypium hirsutum and Arabidopsis thaliana by using TBtools. Additionally, we used MG2C_v2.1 Web Suite (http://mg2c.iask.in/mg2c_v2.1/) (accessed on 24 October 2023) [48] to visualize the chromosomal location of GhVLN. This analysis was based on the transcript IDs, chromosome numbers, and chromosome lengths.
2.5. GhVLNs Structure Prediction and Protein–Protein Interation
The 3D structures of GhVLN proteins were predicted using the online tool ExPASy SWISS-MODEL (https://swissmodel.expasy.org/) (accessed on 9 October 2023) [49]. When predicting protein structures in three dimensions, several key parameters are essential to assess the quality of the generated models. These parameters included global model quality evaluation (GMQE), coverage, and sequence identity.
GMQE is a value that ranges between 0 and 1, where a score closer to 1 indicates a better-quality model. This reflected the overall reliability of the model used in the prediction process. Coverage, on the other hand, measures the extent to which the target protein sequence aligns with and is covered by the template protein sequence. Finally, the sequence identity indicates how well the amino acid sequences align and match the target and template proteins. A higher sequence identity signifies a more reliable model, contributing to a greater accuracy in predicting the 3D structure of the protein.
In our quest to understand the function of GhVLNs, we built protein interaction networks by utilizing the online tool STRING (https://string-db.org/) (accessed on 9 October 2023) [50], where we configured the Required Score to a medium confidence level of 0.4 and applied a medium FDR Stringency of 5 percent.
2.6. Measurement of Evolutionary Selection Pressure and Cis-Element Analysis
Pairs of genes with similar genetic relationships were chosen based on a phylogenetic tree alignment of sequences using the neighbor-joining (NJ) method within the MEGA11 software [44] and Interactive Tree Of Life (iTOL) Version 6.8.1 (https://itol.embl.de/) (accessed on 10 November 2023) [45]. Subsequently, the TBtools software was used to compute the values for Ka (non-synonymous rate), Ks (synonymous substitution), and Ka/Ks (evolutionary constraint). The divergence time (T) was determined using the formula T = Ks/(2 × 9.1 × 10−9) × 10−6 million years [51]. In general, if Ka/Ks < 1.0, it indicates purifying or negative selection, Ka/Ks = 1.0 represents neutral selection, and Ka/Ks > 1.0 signifies positive selection [52]. We conducted GhVLN cis-element prediction using Plant CARE (https://bioinformatics.psb.ugent.be/webtools/plantcare/html/) (accessed on 19 November 2023) [53] by analyzing a 3.5 kb sequence upstream of the coding region. The results were visualized using the BioSequence Structure Illustrator in the graphics section of TBtools.
2.7. Expression Patterns of GhVLNs in Different Tissues and Stress Conditions
We obtained the expression levels of Gossypium hirsutum RNA-Seq data, measured in Fragments Per Kilobase of transcript per million mapped reads (FPKM), for each GhVLN gene under various stress conditions, including drought, salt, heat, and cold stress, at different time points (0, 1, 3, 6, and 12 h). These data were extracted from the CottonRNA Database (PRJNA490626) (http://ipf.sustech.edu.cn/pub/cottonrna/) (accessed on 25 November 2023), and TBtools was used to visualize and present the gene-expression patterns based on the FPKM values.
3. Results
3.1. Sequence Retrieval of VLN Gene and Characterization of GhVLNs
To identify the Villin (VLN) proteins in cotton, we utilized protein sequences containing gelsolin and villin-headpiece (VHP) domains as query sequences from the PFAM database. These sequences comprised six gelsolin domains and a villin-headpiece domain (Figure 1A).
We initiated a BLAST search against the Gossypium hirsutum protein database available on the Phytozome 13 website, followed by an NCBI Batch Conserved Domain search to eliminate redundant sequences. A total of 14 VLN proteins were identified in Gossypium hirsutum.
To obtain a more profound understanding of GhVLNs, we extensively examined their diverse physicochemical characteristics. These included chromosome number, amino acid protein length, isoelectric point (pI), molecular weight (MW), subcellular distribution, and grand average of hydropathy (GRAVY). GhVLN proteins are 902–980 amino acids in length. Their estimated molecular masses varied from 100.72 to 107.37 kDa, with Gohir.D01G187300 having the longest protein length (980 A.A) and Gohir.D08G029300 having the highest molecular mass (107.37 kDa) (Figure 1B). The predicted isoelectric points of the GhVLNs ranged from 5.24 to 6.23, with all pI values greater than 5.25. Additionally, initial predictions indicated that most GhVLNs were localized to the nucleus (Figure 2).
Interestingly, we observed that certain paralogous of GhVLNs sequences, such as Gohir.D01G187300 and Gohir.A01G197100, had identical protein lengths and isoelectric points but differed in molecular weights. Similarly, Gohir.D03G069500 and Gohir.A02G107500 shared the same protein length but exhibited slight variations in molecular weight and isoelectric point (Figure 1B).
Additionally, by employing the VLN protein sequence from Arabidopsis thaliana as a model, we conducted a multiple-sequence-alignment analysis of Gossypium hirsutum. This analysis revealed that all 14 GhVLNs featured six distinctive gelsolin domains and one headpiece domain (Figure 3 and Figure 4). These observations suggest that the existence of both shared and distinct protein functions and characteristics within GhVLNs may contribute to diversifying the Villin gene function in Gossypium hirsutum.
3.2. Phylogenetic Analysis of Gossypium Species and Their Orthologs from Other Model Species
To investigate the evolutionary connections between the GhVLN across various Gossypium species and their counterparts in some different species, we conducted a phylogenetic analysis. To compile candidate VLN protein sequences, we examined protein databases encompassing both lower and higher plants, including monocotyledons and dicotyledons and also some fungal, algal, and animal species. Candidate sequences were identified by querying the respective databases using the initial query sequences. We ensured that the selected proteins included the conserved gelsolin domain and used the headpiece domain for phylogenetic analysis.
In total, we obtained 94 VLN protein sequences distributed across various Gossypium species (Supplementary Table S1).
The phylogenetic analysis successfully classified the 94 VLN proteins into three discernible clades, designated as A, B, and C. Clade A was characterized by a solitary VLN protein, whereas Clades B and C exhibited larger memberships, comprising 40 and 53 VLN proteins, respectively (Figure 5). The phylogenetic tree revealed that Gotom.D02G199000, derived from Gossypium tomentosum, is the sole member of Clade A, exhibiting a notable divergence from the midroot point. Meanwhile, the remaining sequences displayed an uneven distribution across Clades B and C, hinting at intriguing evolutionary relationships (Figure 5).
The phylogenetic analysis of GhVLNs, alongside VLN proteins from diverse representative species, resulted in the classification of 115 VLNs into four distinct clades: A, B, C, and D. Clade A comprised 22 VLN proteins, while Clades B, C, and D contained 3, 44, and 46 VLN protein sequences, respectively. These clades were systematically distributed based on their evolutionary resemblances (Figure 6). Villin protein sequences were sourced from various taxa, including Animalia (12 sequences), Fungi (3 sequences), Amoebozoa (3 sequences), Algae (4 sequences), and Plantae, further categorized into monocots (35 sequences) and dicots (58 sequences) (Supplementary Table S2). Interestingly, the number of VLN proteins varied among different species. Notably, a tree clade comprising Clades B, C, and D, which share a common ancestral origin, exhibited the maximum diversity in Villin gene evolution. This clade encompassed a range of species from lower to higher plants, as well as lower to higher classes of animals and fungi, underscoring the ancient origin of the Villin gene family. Clade B, the smallest clade with only three sequences, included the algal species Chlamydomonas reinhardtii, alongside animalia species such as Mus musculus and Danio rerio, suggesting a potential shared ancestry. Clade D emerged as the largest, comprising 46 sequences, predominantly from dicot plants, particularly within the Fabaceae family (16 species), and monocot plants, primarily within the Poaceae family (11 species). Notably, Clade D excluded species from the Animalia, Fungal, Algal, and Amoebozoa groups. Similarly, Clade C exhibited abundant representation from plant species, spanning monocot and dicot families (Figure 6). Within Clade A, there was a notable abundance of animalia species, including mammals (such as Homo sapiens, Mus musculus, and Bos taurus), Aves (such as Gallus gallus, Coturnix japonica, and Meleagris gallopavo), and aquatic species (such as Labeo rohita, Danio rerio, and Strongylocentrotus purpuratus). These species are found in similar subclades, suggesting a potential common ancestry with some Fungal and Amoebazoa species. Notably, only two plantae species (Striga asiatica and Marchantia polymorpha) are present in Clade A, and they share similar subclades with fungi, such as Aspergillus fishcheri, and certain algal species. Notably, it was found that sequences of Gossypium hirstusum were only found in Clades C and D (Figure 6), where all the higher plant species were present, and the Gossypium hirstusum sequences showed homology with its counterpart sequences only.
These results from the evolutionary analysis suggest that the VLN gene underwent divergence during the evolution of different species yet remained highly conserved in higher plants consisting of monocotyledons and dicotyledons.
3.3. Gene Structure Analysis of GhVLNs and the Conserved Domain and Motif Analysis of GhVLNs
To delve deeper into the structural characteristics of GhVLNs and the proteins they encode, we conducted a comprehensive examination of their gene structures, conserved motifs, and domains. Our analysis of GhVLN genes also involved investigating their exon/intron structures and identifying conserved motifs (Figure 7A–D). Notably, these genes exhibit a significant number of exons and introns, indicating extensive genomic sequences. The number of introns across the 14 GhVLNs varied from 21 to 24 (Figure 7D).
We also investigated the motifs and domains present in the GhVLN proteins to enhance our understanding of their conservation and diversification. Twenty distinct motifs were identified and labeled as Motifs 1–20. Additionally, our analysis showed conservation of the GhVLN domains, characterized by the presence of six gelsolin domains and a headpiece domain (VHP) (Figure 7C). Gene structural analyses collectively suggest that GhVLNs genes and their encoded proteins possess complementary structures, and these structures may explain their functional distinctions. This knowledge may provide a foundational structural framework for understanding the conversed gene functions.
The sequences corresponding to the 20 identified motifs vary in length and span from 21 to 50 amino acids (Table 1). Notably, the counts of these conserved motifs differed among GhVLN proteins, ranging from 16 to 19 motifs per protein (Figure 7B). Proteins identified by the transcript IDs Gohir.D08G029300, Gohir.A08G018700, Gohir.A05G089000, Gohir.A13G212400, Gohir.D13G216500, and Gohir.D05G090000 were found to be without motifs 10 and 18. However, these proteins exclusively contained motif 19. Additionally, Gohir.A13G212400 and Gohir.D13G216500 lacked motifs 9 and 15. Furthermore, Gohir.A05G089000 and Gohir.D05G090000 lacked motif 20, and Gohir.D11G329100 and Gohir.A11G312600 were devoid of motif 11. The DUF4045 superfamily domain was found in proteins Gohir.A02G107500 and Gohir.D03G069500 (Figure 7C). Most GhVLN proteins consist of six gelsolin homology domains (gelsolin s1 to s6-like) and a villin-headpiece domain (VHP), consistently, as mentioned elsewhere. However, it is important to note that exceptions exist, as Gohir.A13G212400 and Gohir.D13G216500 lacks the (G1) and (G1-2) homology domains, respectively, and two proteins, Gohir.D11G329100 and Gohir.A11G312600, do not possess the headpiece domain (VHP) (Figure 7C).
Hence, the gelsolin homology domains exhibited a remarkable degree of conservation within cotton VLNs. Furthermore, GhVLN orthologs that shared close evolutionary relationships displayed analogous motif architectures and GhVLNs exon/intron distribution patterns, highlighting similarities in their structural characteristics.
3.4. Collinearity and Chromosomal Location Analysis of GhVLNs Gene
To delve deeper into the functional mechanisms of Gossypium hirsutum VLNs, we conducted a collinearity analysis using the genomes of Arabidopsis thaliana and Gossypium hirsutum. This analysis revealed 12 orthologous pairs of VLNs between the two genomes (Figure 8A). Importantly, it was observed that several VLN genes in Gossypium hirsutum displayed synteny with the same VLN gene in Arabidopsis thaliana, suggesting a scenario where GhVLNs may have originated from a common ancestor through duplication events during evolution. These results suggest that VLNs were relatively conserved between Arabidopsis thaliana and Gossypium hirsutum during the evolution of higher plants.
In the analysis of collinearity of GhVLNs with Arabidopsis, we observed that the VLN genes were unevenly distributed across 12 of the 26 chromosomes in Gossypium hirsutum (Figure 8A). GhVLNs are located at the chromosomal termini of A01, D01, A02, A05, D05, A08, D08, A11, D11, A13, and D13. However, on chromosomes A01, D02, and D03, VLN locations were near the centromeres of the chromosomes (Figure 8B).
3.5. Three-Dimensional Structure Prediction and Protein–Protein Interaction Network
In the analysis of 3D-structure prediction, three key parameters—Global Model Quality Evaluation (GMQE), coverage, and sequence identity—were employed to assess and select templates. A higher value for these parameters signifies greater accuracy, and the template with the highest score was chosen. In our study, GMQE values consistently fell within the range of 0.73 to 0.79, with a 100% coverage rate and sequence identity ranging from 70.12 to 99.38. These values indicate a high level of reliability for the selected templates.
Furthermore, in the same subgroup, the prediction templates remained consistent, highlighting their evolutionary conservation, except in Groups 1 and 4. This underscores the reliability of the selected prediction templates. Overall, these results suggest that GhVLN proteins within the same subgroup exhibit highly similar spatial structures (Figure 9). Villin belongs to the gelsolin superfamily and functions as a protein involved in F-actin nucleation, crosslinking, severing, and capping. The villin headpiece is crucial for anchoring villin to F-actin, facilitating the process of crosslinking [16]. Detailed information regarding protein structure predictions, including metrics like Coverage, GMQE (Global Model Quality Estimation), and identity scores, are presented in (Supplementary Table S3).
To gain a deeper understanding of the role of the GhVLN protein family in plant development, we predicted protein-interaction networks and three-dimensional structures of all GhVLN proteins.
Our analysis revealed that GhVLN proteins primarily interact with various cytoskeletal proteins and Golgi function-related proteins and play roles in stress responses, including interactions with arabinogalactan proteins (AGPs), the conserved oligomeric Golgi complex (COG), and leucine-rich repeat extensin proteins (LRXs) (Figure 10). Additionally, GhVLN proteins engage with proteins related to development, stress responses, and hormones, suggesting their involvement in signaling response pathways. Detailed information regarding these protein interactions is listed in (Supplementary Table S4). Moreover, there were several other proteins, including LOC107907026, whose protein families have not yet been identified. Initial functional predictions suggest that these proteins also play crucial roles in various cellular physiological processes (Figure 10).
3.6. Analysis of Evolutionary Selection Pressure
Calculating Ka (non-synonymous substitution), Ks (synonymous substitution), and Ka/Ks (evolutionary selection pressure) is of significant importance in determining phylogenetic relationships and gaining insights into evolutionary dynamics within and among species [7]. The Ka/Ks ratio serves as a valuable indicator for assessing the selection pressure in recurrent events. When Ka/Ks is less than one, it signifies purifying selection; when it equals one, it indicates neutral selection; and when it exceeds one, it suggests a positive selection [7].
Therefore, we computed Ka/Ks values for collinear gene pairs within Gossypium hirsutum (Table 2). The Ka/Ks ratios for GhVLN gene pairs ranged from 0.096 to 0.305, with an average of 0.201. Notably, all Ka/Ks values for GhVLN gene pairs were less than one, implying that these genes evolved predominantly under the influence of purifying selection (Table 2).
The estimated divergence time for these seven gene pairs ranged from 1.565 million years ago (MYA) for Gohir.D01G187300-Gohir. A01G197100 to 3.160 (MYA) for Gohir.A13G212400-Gohir.D13G216500 with an average divergence time of approximately 2.396 (MYA) (Table 2).
3.7. Cis-Element and Expression Analysis of GhVLNs
To further explore the potential biological roles of GhVLNs, we conducted a comprehensive analysis of cis-elements present in the predicted promoter regions of these genes. We focused on a 3.5 kb segment located upstream of the GhVLN start codon, using it as the promoter region for our investigation. The identification of cis-elements in the GhVLNs promoter region was carried out using the online Plant CARE tool, and sequence illustrative plots were generated using TBtools (Figure 11A–C).
Our analysis revealed that the promoter region of GhVLNs contains a variety of cis-elements that are associated with key hormonal biological processes. These include elements linked to Abscisic Acid (ABRE), MeJA (TGACG-motif, TATC-box, and CGTCA-motif), Gibberellin (GARE-motif and P-box), Ethylene (ERE), Salicylic Acid (TCA-elements), and Auxin (TGA-elements) responsiveness (Figure 11A). Moreover, we observed the presence of cis-elements related to environmental stress responses, such as Anaerobic Induction (ARE), elicitation (box S), wound (WUN-motif), pathogen (W box), low temperature (LTR), drought inducibility (MBS), and light, defense, and stress (TC-rich repeats) responsiveness (Figure 11B).
It is worth noting that some motifs, such as Heat stress (HSE), Fungal Elicitor (Box-W1, W3), Dehydration, and Salt stress (DRE)-responsive elements, were absent in the GhVLNs’ promoter region. Notably, the most prominent elements were related to the light response. Furthermore, the promoter region contained cis-elements associated with meristem expression, circadian control, endosperm expression, and the regulation of flavonoid biosynthetic genes (Figure 11C).
This comprehensive cis-element analysis suggests that GhVLNs may be involved in a range of signaling pathways and biological processes. However, it is important to emphasize that, while these findings are intriguing, further research is needed to substantiate these speculations. Analyzing the gene function of GhVLNs in the context of their responses to external- and internal-environmental signals holds great promise for advancing our understanding in this area.
3.8. Gene-Expression Pattern Analysis in Different Organs of GhVLNs
To investigate the potential functions of GhVLNs in various tissues of Gossypium hirsutum, we obtained tissue-specific expression data from the CottonRNA Database (PRJNA490626). The tissues examined included anthers, bracts, filaments, leaves, petals, pistils, roots, sepals, stems, ovules on different days post-anthesis (dpa), and fibers at different dpa stages (10, 15, 20, and 25 dpa). An analysis of the expression profiles of the 14 GhVLN genes revealed distinct spatial-expression patterns. For instance, Gohir.A05G089000 exhibited notably high expression levels in sepals, stems, and tori, whereas Gohir.D03G069500 showed elevated expression levels in filaments, petals, and roots (Figure 12C).
In ovule samples at different developmental stages, Gohir.A05G089000 showed the most prominent expression. In the case of fiber samples at various developmental stages, Gohir.A08G018700 had the highest expression at 10, 15, and 20 dpa, followed by Gohir.A05G089000, indicating its potential role in cell-wall thickening during fiber development. Gohir.A05G089000 was particularly expressed in 3-dpa ovules (Figure 12B).
Furthermore, Gohir.A02G107500, followed by Gohir.D03G069500, was found to be associated with stress responses, including cold, drought, heat, and salt environments (Figure 13). These genes could be considered candidates for future transformation experiments aimed at understanding their roles in cotton-fiber development.
4. Discussion
The VLN gene family plays a crucial role in various aspects of plant growth and development, and possesses an actin-binding domain. It acts as a group of regulators that control actin dynamics by polymerizing and depolymerizing actin filaments [54]. In different developmental stages and under the influence of environmental stress, VLN can modify actin filaments by either severing or bundling them [31,54]. Despite this, prior to our study, there has very limited investigation made for the VLN gene family in Gossypium hirsutum, a significant and widely cultivated fiber and cash crop. This genome-wide identification and characterization of the Gossypium hirsutum VLN gene family represents a vital initial step towards a deeper understanding of the functions of this gene family.
Several studies have investigated the functions of VLN genes in the regulation of plant architecture, which directly affects crop yields. These VLN genes control the development of various plant tissues by orchestrating the arrangement of actin filaments in Arabidopsis thaliana [5,31]. Notably, functional redundancies exist in AtVLN2 and AtVLN3, both of which jointly influence the plant architecture. The simultaneous mutation of VLN2 and VLN3 results in distorted roots, stems, leaves, pods, and inflorescences [31,31]. In agricultural settings, twisted organs can adversely affect photosynthesis, biomass, and harvest. Moreover, AtVLN5 plays a pivotal role in regulating pollen germination and pollen tube growth during the reproductive phase [31]. In light of these findings, it is evident that VLNs play a crucial role in the regulation of crop yield, offering insights into the genetic foundations of crop improvement.
In our investigation, we identified and characterized 14 Villin protein sequences in Gossypium hirsutum (Figure 1B). These genes have varying lengths, ranging from 902 to 980 amino acids, with molecular weights exceeding 100 kDa (Figure 1B). Subcellular localization analysis indicated their presence in the nucleus (Figure 2), which is consistent with the findings of previous studies on AtVLNs [55]. The amino acid alignment of the GhVLNs proteins represents the highly conserved nature of the protein sequences represented through distinct colors and through WebLogo (Figure 3 and Figure 4), respectively. The phylogenetic analysis of different species of Gossypium demonstrated that all VLNs could be grouped into three subcategories, A, B, and C (Figure 5), according to the clades dividing from the midroot point, which were unevenly spread across the tree.
In our analysis of phylogenetic relationships across various model species and families, there were five distinct subcategories, A, B, C, and D, divided according to the midroot point (Figure 6). It was observed that the majority of GhVLNs were clustered within subgroups C and D (Figure 6). This trend was consistent not only within Gossypium hirsutum but also among other higher plants, including Arabidopsis thaliana, Zea mays, and Oryza sativa. Interestingly, VLNs from the lower plant Chlamydomonas reinhardtii, although fewer in number, were predominantly distributed in subgroup B. Remarkably, animalia species were consistently grouped in subgroup A (Figure 6). This pattern suggests a certain level of functional conservation in the gene family across higher plants, encompassing both monocots and dicots, as well as animalia species, which share some common ancestry with fungal and algal species. Our investigation further revealed that the majority of Gossypium hirsutum VLNs contained six gelsolin domains (G1–G6) and a villin-headpiece domain (VHP) (Figure 1A) which was verified by the conserved domains of the GhVLNs (Figure 7C). Additionally, 20 sequences were identified, the majority of which belong to the ADF and gelsolin superfamily (Table 1) (Figure 7B). These structural configurations align with the findings of previous studies [56,57].
The collinearity analysis using the Arabidopsis genome suggested that the synteny between the GhVLN gene and Arabidopsis may share a common ancestor during its evolution (Figure 8A). The 3D-structure prediction suggested that the GhVLNs exhibit similar spatial structures which were consisted with previous studies on Glycine max [57]. Furthermore, we employed protein–protein interaction and cis-element analysis to assess the gene expression pattern within the promoter region [58]. This analysis revealed the existence of diverse hormone- and stress-responsive elements in the promoter regions of GhVLN genes (Figure 11). The protein–protein interaction revealed that GhVLN proteins interact with various cytoskeletal proteins, hence playing a crucial role in stress responses (Figure 10). This observation implies that these genes likely play crucial roles in response to stress, aligning with similar findings in other species [56,57]. This outcome is in line with findings from other studies, where AtVLNs exhibited distinct expression patterns featuring elevated expression levels and a preference for specific tissues [59]. Furthermore, GhVLNs are broadly expressed in various tissues and organs. This widespread expression implies a significant role in regulating the stress tolerance, growth, and development of Gossypium hirsutum. Enhancing stress responses is a pivotal strategy for enhancing crop agronomic traits and increasing yield. This includes bolstering resistance to high temperatures, cold, salt, and drought, and responsiveness to hormones. Notably, Gohir.A02G107500, Gohir.D03G069500, Gohir.A05G089000, and Gohir.D03G069500 were primarily linked to stress responses and were expressed during different developmental stages (Figure 12 and Figure 13). Our analysis of cis-elements indicated that GhVLNs may participate in various processes, including hormone signaling and responses to light, drought, and defense mechanisms (Figure 11). Furthermore, a particularly intriguing observation captured our attention: specifically, the presence of pathogen-related cis-elements (W box) and wound-response elements (WUN-motif) within the promoter regions of certain GhVLNs (Figure 11), all of which were significantly induced under various biotic and abiotic stress treatments. Moreover, previous research has documented their involvement in the response to salt and drought stress, as well as their essential role in conferring tolerance to Verticillium infection in cotton. [60]. Notably, the high expression of Gohir.A025G089000 in sepals, stems, and torus suggests its potential involvement in the regulation of growth and elongation traits, which are crucial factors for Gossypium hirsutum yield regulation (Figure 12). The roles of the five Villins in Arabidopsis have been thoroughly characterized, and the GhVLNs investigated in this study are phylogenetically related to Arabidopsis VLN4, which is known to be involved in root-hair growth through the regulation of actin organization [5]. GhVLN4 exhibits ubiquitous expression in cotton tissues, with its predominant presence in elongating young fibers. Previous reports have highlighted the crucial roles of GhVLN4 in both tip and diffuse growth through the regulation of actin organization [27]. It has also been shown that VLN is directly involved in the developmental regulation of root hairs in plants under osmotic stress [30,61]. Our study revealed a high expression of Gohir.A08G018700 during fiber development from 15DPA to 25DPA (Figure 12A), indicating its involvement in the fiber-elongation phase, which is also consistent with some existing studies [56]. Given that cotton is primarily cultivated for its fiber yield, Villin genes play a crucial role in fiber elongation and development. To breed high-quality, high-yield Gossypium hirsutum varieties in the future, it is imperative to explore the function of the VLN gene family in shaping plant architecture and crop yield. Despite numerous studies on plant VLNs related to their role in regulating plant growth, development, and response to environmental stresses, our understanding of Gossypium hirsutum VLNs remains at an early stage.
Therefore, more investigations are required to assess and validate the effect of VLNs on the growth and development of Gossypium hirsutum. The present study of a genome-wide analysis may lay the groundwork for further exploration into the functions of Gossypium hirsutum VLNs, potentially offering valuable insights that could contribute to cotton breeding efforts.
5. Conclusions
The present study investigated the phylogeny and characteristics of the VLN genes in Gossypium hirsutum from various perspectives. All 14 GhVLN protein sequences were categorized into three distinct groups within the Gossypium species. Sequences within the same group exhibit comparable evolutionary features, such as three-dimensional structure, gene structure, motifs, and conserved domains, suggesting similar potential functions. Moreover, these GhVLN genes were distributed across 12 of the 26 chromosomes in the Gossypium hirsutum genome, and their amino acid sequences displayed a high degree of similarity. The collinearity analysis suggested that GhVLNs share functional similarities with VLN genes in the model species Arabidopsis. An examination of expression patterns across various tissues indicated that GhVLN genes are extensively expressed in different tissues, and that there may be functional overlaps among different GhVLN members. An analysis of protein-interaction networks and cis-elements suggests that GhVLNs may be involved in various physiological processes, including responses to hormones, stress, and developmental signals. The insights obtained from our study lay a robust foundation for future studies on the function of VLN genes in Gossypium and other higher plant species.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/cimb46030146/s1, Table S1: Protein sequences of all 94 VLNs in Gossypium species (Gossypium hirsutum, Gossypium arboreum, Gossypium barbadense, Gossypium darwinii, Gossypium herbaceum, Gossypium mustelinum, Gossypium rainmondii, and Gossypium tomentosum); Table S2: Protein sequences of all 115 VLNs, including Gossypium hirsutum and VLNs from other representative models from plants, algae, fungi, and animal species; Table S3: Detailed information regarding protein structure predictions, including metrics such as coverage, Global Model Quality Estimation (GMQE), and identity scores for the three-dimensional prediction of GhVLNs proteins using SWISS-MODEL; Table S4: Detailed information regarding protein–protein interactions, including predicted locus, function, and score metrics.
Author Contributions
Conceptualization, D.K.P.; Methodology, A.D. and D.K.P.; Validation, A.D. and D.K.P.; Investigation, A.D. and D.K.P.; Writing—original draft preparation, A.D.; Writing—review and editing, A.D. and D.K.P.; Supervision, D.K.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All supporting data are either provided with main text or as Supplementary Files.
Acknowledgments
The authors are thankful to Amity University Jharkhand for the support provided under NTCC program.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Yuan, G.; Gao, H.; Yang, T. Exploring the Role of the Plant Actin Cytoskeleton: From Signaling to Cellular Functions. Int. J. Mol. Sci. 2023, 24, 15480. [Google Scholar] [CrossRef]
- Pollard, T.D. Actin and Actin-Binding Proteins. Cold Spring Harb. Perspect. Biol. 2016, 8, a018226. [Google Scholar] [CrossRef]
- Pandey, D.K.; Chaudhary, B. Transcriptional Loss of Domestication-Driven Cytoskeletal GhPRF1 Gene Causes Defective Floral and Fiber Development in Cotton (Gossypium). Plant Mol. Biol. 2021, 107, 519–532. [Google Scholar] [CrossRef]
- Qian, D.; Zhang, Z.; He, J.; Zhang, P.; Ou, X.; Li, T.; Niu, L.; Nan, Q.; Niu, Y.; He, W.; et al. Arabidopsis ADF5 Promotes Stomatal Closure by Regulating Actin Cytoskeleton Remodeling in Response to ABA and Drought Stress. J. Exp. Bot. 2019, 70, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xiao, Y.; Du, F.; Cao, L.; Dong, H.; Ren, H. Arabidopsis VILLIN4 Is Involved in Root Hair Growth through Regulating Actin Organization in a Ca2+-Dependent Manner. New Phytol. 2011, 190, 667–682. [Google Scholar] [CrossRef] [PubMed]
- Christensen, H.E.M.; Ramachandran, S.; Tan, C.; Surana, U.; Dong, C.; Chua, N. Arabidopsis Profilins Are Functionally Similar to Yeast Profilins: Identification of a Vascular Bundle-specific Profilin and a Pollen-specific Profilin. Plant J. 1996, 10, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Pandey, D.K.; Chaudhary, B. Evolutionary Expansion and Structural Functionalism of the Ancient Family of Profilin Proteins. Gene 2017, 626, 70–86. [Google Scholar] [CrossRef]
- Pandey, D.K.; Chaudhary, B. Evolution of Functional Diversity Among Actin-Binding Profilin Genes in Land Plants. Front. Cell Dev. Biol. 2020, 8, 588689. [Google Scholar] [CrossRef]
- Ramachandran, S.; Christensen, H.E.M.; Ishimaru, Y.; Dong, C.-H.; Chao-Ming, W.; Cleary, A.L.; Chua, N.-H. Profilin Plays a Role in Cell Elongation, Cell Shape Maintenance, and Flowering in Arabidopsis. Plant Physiol. 2000, 124, 1637–1647. [Google Scholar] [CrossRef]
- Müssar, K.J.; Kandasamy, M.K.; McKinney, E.C.; Meagher, R.B. Arabidopsis Plants Deficient in Constitutive Class Profilins Reveal Independent and Quantitative Genetic Effects. BMC Plant Biol. 2015, 15, 177. [Google Scholar] [CrossRef]
- Pandey, D.K.; Chaudhary, B. Domestication-Driven Gossypium Profilin 1 (GhPRF1) Gene Transduces Early Flowering Phenotype in Tobacco by Spatial Alteration of Apical/Floral-Meristem Related Gene Expression. BMC Plant Biol. 2016, 16, 112. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Zhang, S.; Liu, J.; Liu, S.; Su, H. Arabidopsis FIM4 and FIM5 Regulates the Growth of Root Hairs in an Auxin-Insensitive Way. Plant Signal. Behav. 2018, 13, e1473667. [Google Scholar] [CrossRef] [PubMed]
- Hesterberg, L.K.; Weber, K. Demonstration of Three Distinct Calcium-Binding Sites in Villin, a Modulator of Actin Assembly. J. Biol. Chem. 1983, 258, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Tomar, A.; Parrill, A.L.; Khurana, S. Functional Dissection and Molecular Characterization of Calcium-Sensitive Actin-Capping and Actin-Depolymerizing Sites in Villin. J. Biol. Chem. 2004, 279, 45036–45046. [Google Scholar] [CrossRef]
- Zhai, L.; Zhao, P.; Panebra, A.; Guerrerio, A.L.; Khurana, S. Tyrosine Phosphorylation of Villin Regulates the Organization of the Actin Cytoskeleton. J. Biol. Chem. 2001, 276, 36163–36167. [Google Scholar] [CrossRef]
- Hampton, C.M.; Liu, J.; Taylor, D.W.; DeRosier, D.J.; Taylor, K.A. The 3D Structure of Villin as an Unusual F-Actin Crosslinker. Structure 2008, 16, 1882–1891. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.; Tholl, S.; Moes, D.; Dieterle, M.; Papuga, J.; Moreau, F.; Steinmetz, A. Actin Bundling in Plants. Cell Motil. 2009, 66, 940–957. [Google Scholar] [CrossRef]
- Vardar, D.; Buckley, D.A.; Frank, B.S.; McKnight, C.J. NMR Structure of an F-Actin-Binding “Headpiece” Motif from Villin. J. Mol. Biol. 1999, 294, 1299–1310. [Google Scholar] [CrossRef]
- Dos Remedios, C.G.; Chhabra, D.; Kekic, M.; Dedova, I.V.; Tsubakihara, M.; Berry, D.A.; Nosworthy, N.J. Actin Binding Proteins: Regulation of Cytoskeletal Microfilaments. Physiol. Rev. 2003, 83, 433–473. [Google Scholar] [CrossRef]
- Huang, S.; Qu, X.; Zhang, R. Plant Villins: Versatile Actin Regulatory Proteins. J. Integr. Plant Biol. 2015, 57, 40–49. [Google Scholar] [CrossRef]
- Su, H.; Wang, T.; Dong, H.; Ren, H. The Villin/Gelsolin/Fragmin Superfamily Proteins in Plants. J. Integr. Plant Biol. 2007, 49, 1183–1191. [Google Scholar] [CrossRef]
- Tominaga, M.; Yokota, E.; Vidali, L.; Sonobe, S.; Hepler, P.K.; Shimmen, T. The Role of Plant Villin in the Organization of the Actin Cytoskeleton, Cytoplasmic Streaming and the Architecture of the Transvacuolar Strand in Root Hair Cells of Hydrocharis. Planta 2000, 210, 836–843. [Google Scholar] [CrossRef] [PubMed]
- Vidali, L.; Yokota, E.; Cheung, A.Y.; Shimmen, T.; Hepler, P.K. The 135 kDa Actin-Bundling Protein fromLilium Longiflorum Pollen Is the Plant Homologue of Villin. Protoplasma 1999, 209, 283–291. [Google Scholar] [CrossRef]
- Yokota, E.; Vidali, L.; Tominaga, M.; Tahara, H.; Orii, H.; Morizane, Y.; Hepler, P.K.; Shimmen, T. Plant 115-kDa Actin-Filament Bundling Protein, P-115-ABP, Is a Homologue of Plant Villin and Is Widely Distributed in Cells. Plant Cell Physiol. 2003, 44, 1088–1099. [Google Scholar] [CrossRef] [PubMed]
- Yokota, E.; Shimmen, T. The 135-kDa Actin-Bundling Protein from Lily Pollen Tubes Arranges F-Actin into Bundles with Uniform Polarity. Planta 1999, 209, 264–266. [Google Scholar] [CrossRef] [PubMed]
- Khurana, S. Role of Actin Cytoskeleton in Regulation of Ion Transport: Examples from Epithelial Cells. J. Membr. Biol. 2000, 178, 73–87. [Google Scholar] [CrossRef]
- Lv, F.; Han, M.; Ge, D.; Dong, H.; Zhang, X.; Li, L.; Zhang, P.; Zhang, Z.; Sun, J.; Liu, K.; et al. GhVLN4 Is Involved in Cell Elongation via Regulation of Actin Organization. Planta 2017, 246, 687–700. [Google Scholar] [CrossRef]
- Pina, C.; Pinto, F.; Feijó, J.A.; Becker, J.D. Gene Family Analysis of the Arabidopsis Pollen Transcriptome Reveals Biological Implications for Cell Growth, Division Control, and Gene Expression Regulation. Plant Physiol. 2005, 138, 744–756. [Google Scholar] [CrossRef]
- Staiger, C.J.; Hussey, P.J. Actin and Actin-Modulating Proteins. In Annual Plant Reviews Online; Roberts, J.A., Ed.; Wiley: Hoboken, NJ, USA, 2018; pp. 32–80. ISBN 978-1-119-31299-4. [Google Scholar]
- Wang, X.; Bi, S.; Wang, L.; Li, H.; Gao, B.; Huang, S.; Qu, X.; Cheng, J.; Wang, S.; Liu, C.; et al. GLABRA2 Regulates Actin Bundling Protein VILLIN1 in Root Hair Growth in Response to Osmotic Stress. Plant Physiol. 2020, 184, 176–193. [Google Scholar] [CrossRef]
- Bao, C.; Wang, J.; Zhang, R.; Zhang, B.; Zhang, H.; Zhou, Y.; Huang, S. Arabidopsis VILLIN2 and VILLIN3 Act Redundantly in Sclerenchyma Development via Bundling of Actin Filaments. Plant J. 2012, 71, 962–975. [Google Scholar] [CrossRef]
- Van Der Honing, H.S.; Kieft, H.; Emons, A.M.C.; Ketelaar, T. Arabidopsis VILLIN2 and VILLIN3 Are Required for the Generation of Thick Actin Filament Bundles and for Directional Organ Growth. Plant Physiol. 2012, 158, 1426–1438. [Google Scholar] [CrossRef]
- Zhang, H.; Qu, X.; Bao, C.; Khurana, P.; Wang, Q.; Xie, Y.; Zheng, Y.; Chen, N.; Blanchoin, L.; Staiger, C.J.; et al. Arabidopsis VILLIN5, an Actin Filament Bundling and Severing Protein, Is Necessary for Normal Pollen Tube Growth. Plant Cell 2010, 22, 2749–2767. [Google Scholar] [CrossRef]
- Raghavendra, K.P.; Das, J.; Kumar, R.; Gawande, S.P.; Santosh, H.B.; Sheeba, J.A.; Kranthi, S.; Kranthi, K.R.; Waghmare, V.N. Genome-Wide Identification and Expression Analysis of the Plant Specific LIM Genes in Gossypium Arboreum under Phytohormone, Salt and Pathogen Stress. Sci. Rep. 2021, 11, 9177. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, P.; Gao, W.; Long, Y.; Wang, Y.; Geng, S.; Su, X.; Jiao, Y.; Chen, Q.; Qu, Y. Genome-Wide Identification of the DUF668 Gene Family in Cotton and Expression Profiling Analysis of GhDUF668 in Gossypium Hirsutum under Adverse Stress. BMC Genom. 2021, 22, 395. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The Protein Families Database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef] [PubMed]
- Sato, S. Structural Analysis of Arabidopsis Thaliana Chromosome 5. X. Sequence Features of the Regions of 3,076,755 Bp Covered by Sixty P1 and TAC Clones. DNA Res. 2000, 7, 31–63. [Google Scholar] [CrossRef] [PubMed]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A Comparative Platform for Green Plant Genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Derbyshire, M.K.; Yamashita, R.A.; Marchler-Bauer, A. NCBI’s Conserved Domain Database and Tools for Protein Domain Analysis. Curr. Protoc. Bioinform. 2020, 69, e90. [Google Scholar] [CrossRef]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: Recent Updates, New Developments and Status in 2020. Nucleic Acids Res. 2021, 49, D458–D460. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server. In The Proteomics Protocols Handbook; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. ISBN 978-1-58829-343-5. [Google Scholar]
- Pierleoni, A.; Martelli, P.L.; Fariselli, P.; Casadio, R. BaCelLo: A Balanced Subcellular Localization Predictor. Bioinformatics 2006, 22, e408–e416. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Li, C.; Li, Y.; An, Y.; Ruan, X.; Guo, Y.; Dong, X.; Ruan, Y. Genome-Wide Identification and Characterization of the VQ Motif-Containing Gene Family Based on Their Evolution and Expression Analysis under Abiotic Stress and Hormone Treatments in Foxtail Millet (Setaria Italica L.). Genes 2023, 14, 1032. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An Upgraded Gene Feature Visualization Server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef]
- Chao, J.; Li, Z.; Sun, Y.; Aluko, O.O.; Wu, X.; Wang, Q.; Liu, G. MG2C: A User-Friendly Online Tool for Drawing Genetic Maps. Mol. Hortic. 2021, 1, 16. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology Modelling of Protein Structures and Complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING Database in 2021: Customizable Protein–Protein Networks, and Functional Characterization of User-Uploaded Gene/Measurement Sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Y.; Liu, B.; Wang, L.; Zhang, L.; Hu, J.; Chen, J.; Zheng, H.; Lu, M. Characterization of the Populus Rab Family Genes and the Function of PtRabE1b in Salt Tolerance. BMC Plant Biol. 2018, 18, 124. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, J.; Zhao, X.-Q.; Wang, J.; Wong, G.K.-S.; Yu, J. KaKs_Calculator: Calculating Ka and Ks Through Model Selection and Model Averaging. Genom. Proteom. Bioinform. 2006, 4, 259–263. [Google Scholar] [CrossRef]
- Lescot, M. PlantCARE, a Database of Plant Cis-Acting Regulatory Elements and a Portal to Tools for in Silico Analysis of Promoter Sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Khurana, S.; George, S.P. Regulation of Cell Structure and Function by Actin-Binding Proteins: Villin’s Perspective. FEBS Lett. 2008, 582, 2128–2139. [Google Scholar] [CrossRef]
- Khurana, P.; Henty, J.L.; Huang, S.; Staiger, A.M.; Blanchoin, L.; Staiger, C.J. Arabidopsis VILLIN1 and VILLIN3 Have Overlapping and Distinct Activities in Actin Bundle Formation and Turnover. Plant Cell 2010, 22, 2727–2748. [Google Scholar] [CrossRef] [PubMed]
- Lv, F.; Wang, S.; Tian, R.; Wang, P.; Liu, K. Villin Family Members Associated with Multiple Stress Responses in Cotton. Phyton-Int. J. Exp. Bot. 2021, 90, 1645–1660. [Google Scholar] [CrossRef]
- Zhou, Y.; He, L.; Zhou, S.; Wu, Q.; Zhou, X.; Mao, Y.; Zhao, B.; Wang, D.; Zhao, W.; Wang, R.; et al. Genome-Wide Identification and Expression Analysis of the VILLIN Gene Family in Soybean. Plants 2023, 12, 2101. [Google Scholar] [CrossRef] [PubMed]
- Biłas, R.; Szafran, K.; Hnatuszko-Konka, K.; Kononowicz, A.K. Cis-Regulatory Elements Used to Control Gene Expression in Plants. Plant Cell Tissue Organ Cult. PCTOC 2016, 127, 269–287. [Google Scholar] [CrossRef]
- Klahre, U.; Friederich, E.; Kost, B.; Louvard, D.; Chua, N.-H. Villin-Like Actin-Binding Proteins Are Expressed Ubiquitously in Arabidopsis. Plant Physiol. 2000, 122, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Ge, D.; Pan, T.; Zhang, P.; Wang, L.; Zhang, J.; Zhang, Z.; Dong, H.; Sun, J.; Liu, K.; Lv, F. GhVLN4 Is Involved in Multiple Stress Responses and Required for Resistance to Verticillium Wilt. Plant Sci. 2021, 302, 110629. [Google Scholar] [CrossRef]
- Bi, S.; Li, M.; Liu, C.; Liu, X.; Cheng, J.; Wang, L.; Wang, J.; Lv, Y.; He, M.; Cheng, X.; et al. Actin Depolymerizing Factor ADF7 Inhibits Actin Bundling Protein VILLIN1 to Regulate Root Hair Formation in Response to Osmotic Stress in Arabidopsis. PLoS Genet. 2022, 18, e1010338. [Google Scholar] [CrossRef]
Figure 1.
Isolation and identification of GhVLNs: (A) Gelsolin domain (PF00626) and the Villin-headpiece domain (PF02209)-conserved domains in GhVLNs. (B) Comprehensive assessment of physical and chemical attributes of GhVLNs. (C) Evolutionary linkage among between VLNs found in Gossypium hirsutum and Arabidopsis thaliana.
Figure 1.
Isolation and identification of GhVLNs: (A) Gelsolin domain (PF00626) and the Villin-headpiece domain (PF02209)-conserved domains in GhVLNs. (B) Comprehensive assessment of physical and chemical attributes of GhVLNs. (C) Evolutionary linkage among between VLNs found in Gossypium hirsutum and Arabidopsis thaliana.
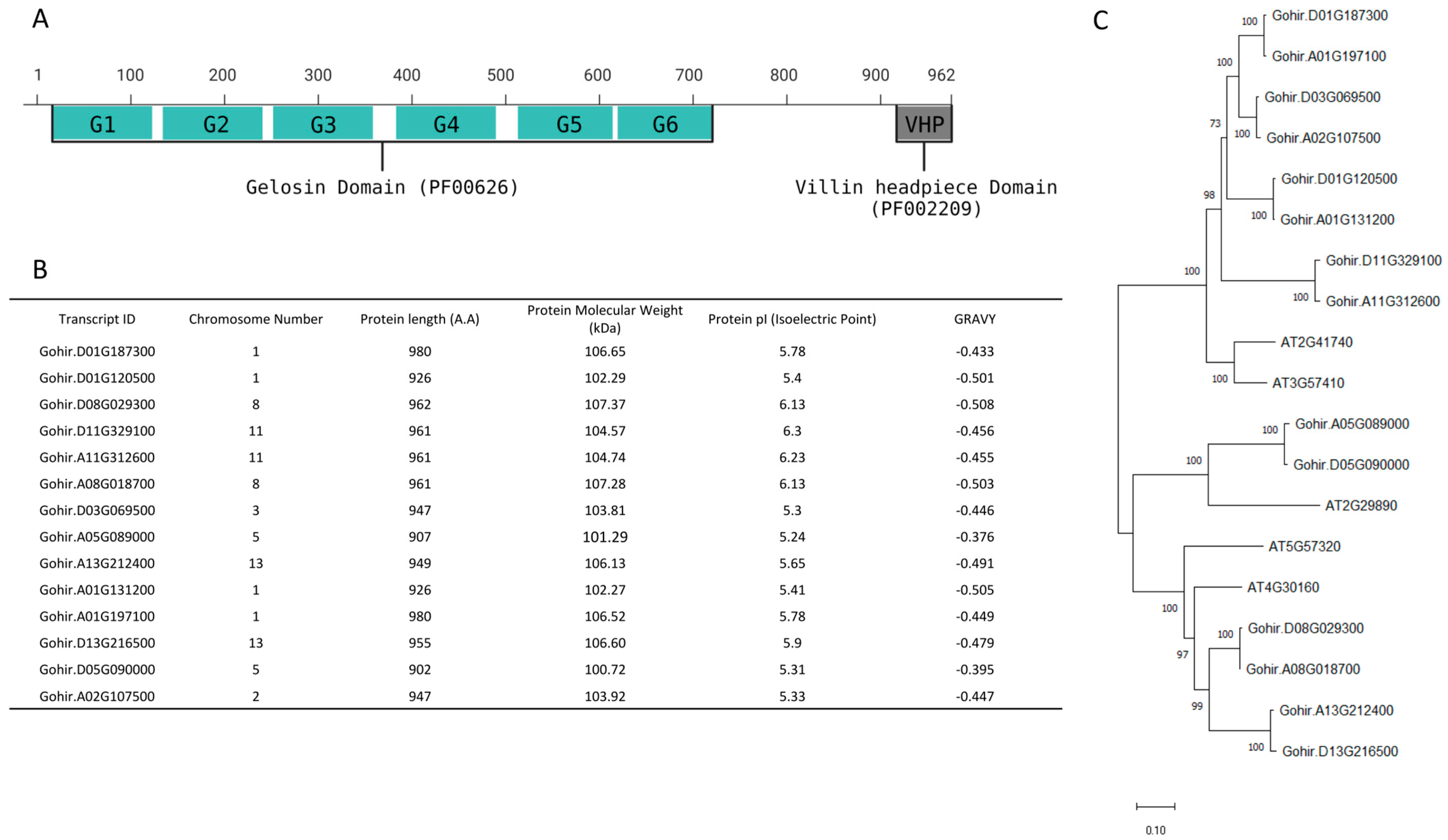
Figure 2.
Prediction scores for WoLF PSORT plant data sets are presented, indicating subcellular localization probabilities for various compartments. Abbreviations such as ‘Nuc’ for Nuclear, ‘Cyt’ for cytosol, ‘Mit’ for mitochondria, ‘Vac’ for vacuole, ‘Cys’ for cytoskeleton, ‘Chl’ for chloroplast, ‘E.R’ for endoplasmic reticulum, ‘Pla’ for plasma membrane, ‘Gol’ for Golgi apparatus, ‘Per’ for peroxisome, and ‘Ext’ for extracellular, are utilized to denote specific cellular locations.
Figure 2.
Prediction scores for WoLF PSORT plant data sets are presented, indicating subcellular localization probabilities for various compartments. Abbreviations such as ‘Nuc’ for Nuclear, ‘Cyt’ for cytosol, ‘Mit’ for mitochondria, ‘Vac’ for vacuole, ‘Cys’ for cytoskeleton, ‘Chl’ for chloroplast, ‘E.R’ for endoplasmic reticulum, ‘Pla’ for plasma membrane, ‘Gol’ for Golgi apparatus, ‘Per’ for peroxisome, and ‘Ext’ for extracellular, are utilized to denote specific cellular locations.

Figure 3.
Amino acid alignment of GhVLN proteins reveals conserved domains, including G1–G6 and a VHP domain, depicted by distinct lines matching these domains across all sequences. This visual representation highlights the conservation of essential motifs within the GhVLN protein family. Different colors delineate the degrees of similarity among 14 VLN protein sequences: cyan shades denote similarity ranging from 50% to 74%, cherry red signifies similarity from 75% to 99%, and deep blue represents 100% similarity.
Figure 3.
Amino acid alignment of GhVLN proteins reveals conserved domains, including G1–G6 and a VHP domain, depicted by distinct lines matching these domains across all sequences. This visual representation highlights the conservation of essential motifs within the GhVLN protein family. Different colors delineate the degrees of similarity among 14 VLN protein sequences: cyan shades denote similarity ranging from 50% to 74%, cherry red signifies similarity from 75% to 99%, and deep blue represents 100% similarity.

Figure 4.
Conserved motifs identified within GhVLNs as G1–G6 and VHP are depicted. These conserved motifs serve as key structural and functional elements within the GhVLN protein family, potentially influencing various biological processes. The colors of amino acids correspond to their chemical properties; polar, basic, acidic, and hydrophobic amino acids are represented by green, blue, red, and black respectively.
Figure 4.
Conserved motifs identified within GhVLNs as G1–G6 and VHP are depicted. These conserved motifs serve as key structural and functional elements within the GhVLN protein family, potentially influencing various biological processes. The colors of amino acids correspond to their chemical properties; polar, basic, acidic, and hydrophobic amino acids are represented by green, blue, red, and black respectively.
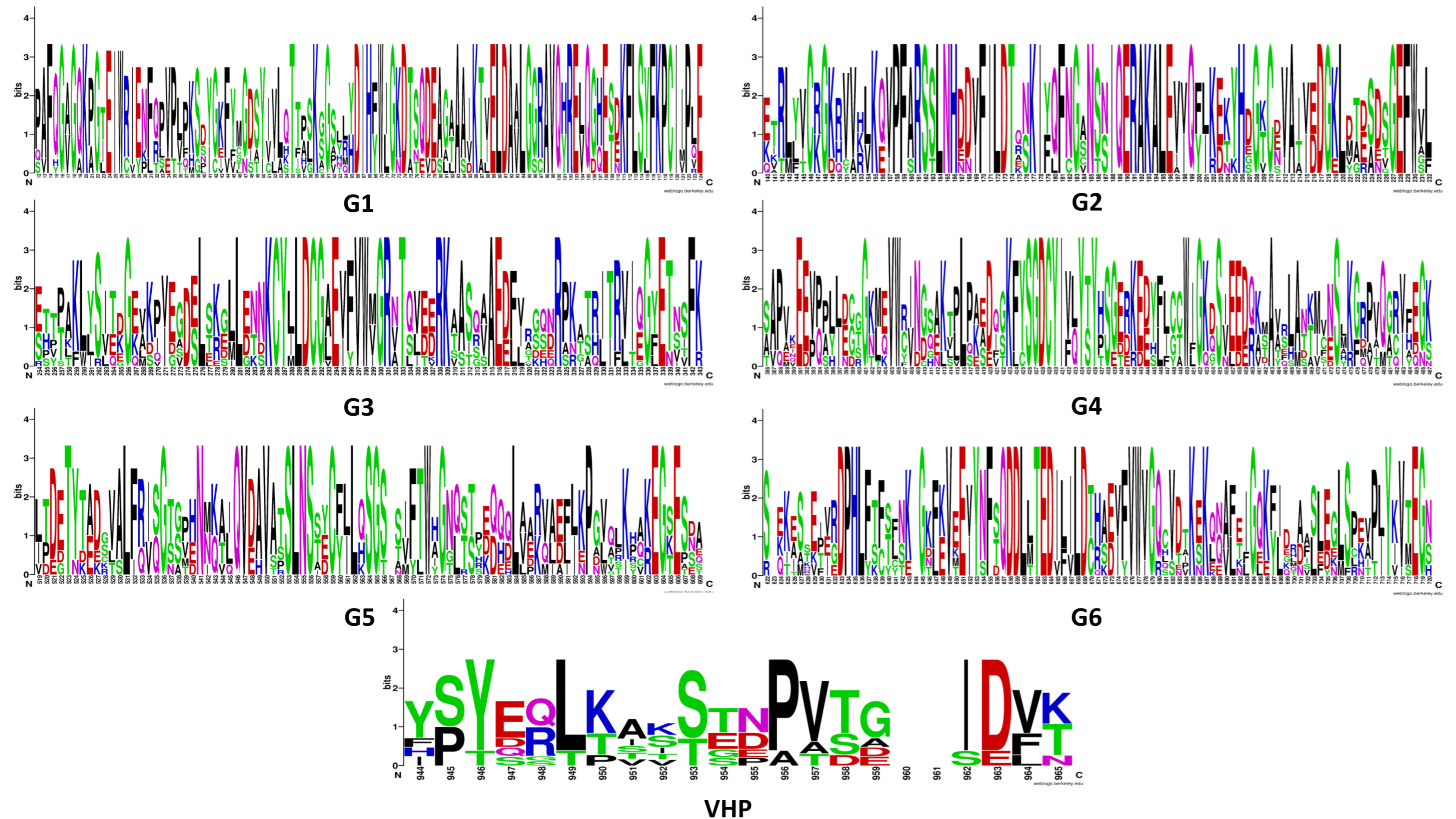
Figure 5.
Phylogenetic analysis of VLN possessing sequences in different Gossypium species including 14 from Gossypium hirsutum (Gohir), 8 from Gossypium arboretum (Cotton_A), 14 from Gossypium barbadense (Gobar), 14 from Gossypium darwinii (Godar), 7 from Gossypium herbaceum (Ghe), 15 from Gossypium mustelinum (Gomus), 7 from Gossypium rainmondii (Gorai), and 15 from Gossypium tomentosum (Gotom) distributed into three distinct clades (A–C).
Figure 5.
Phylogenetic analysis of VLN possessing sequences in different Gossypium species including 14 from Gossypium hirsutum (Gohir), 8 from Gossypium arboretum (Cotton_A), 14 from Gossypium barbadense (Gobar), 14 from Gossypium darwinii (Godar), 7 from Gossypium herbaceum (Ghe), 15 from Gossypium mustelinum (Gomus), 7 from Gossypium rainmondii (Gorai), and 15 from Gossypium tomentosum (Gotom) distributed into three distinct clades (A–C).
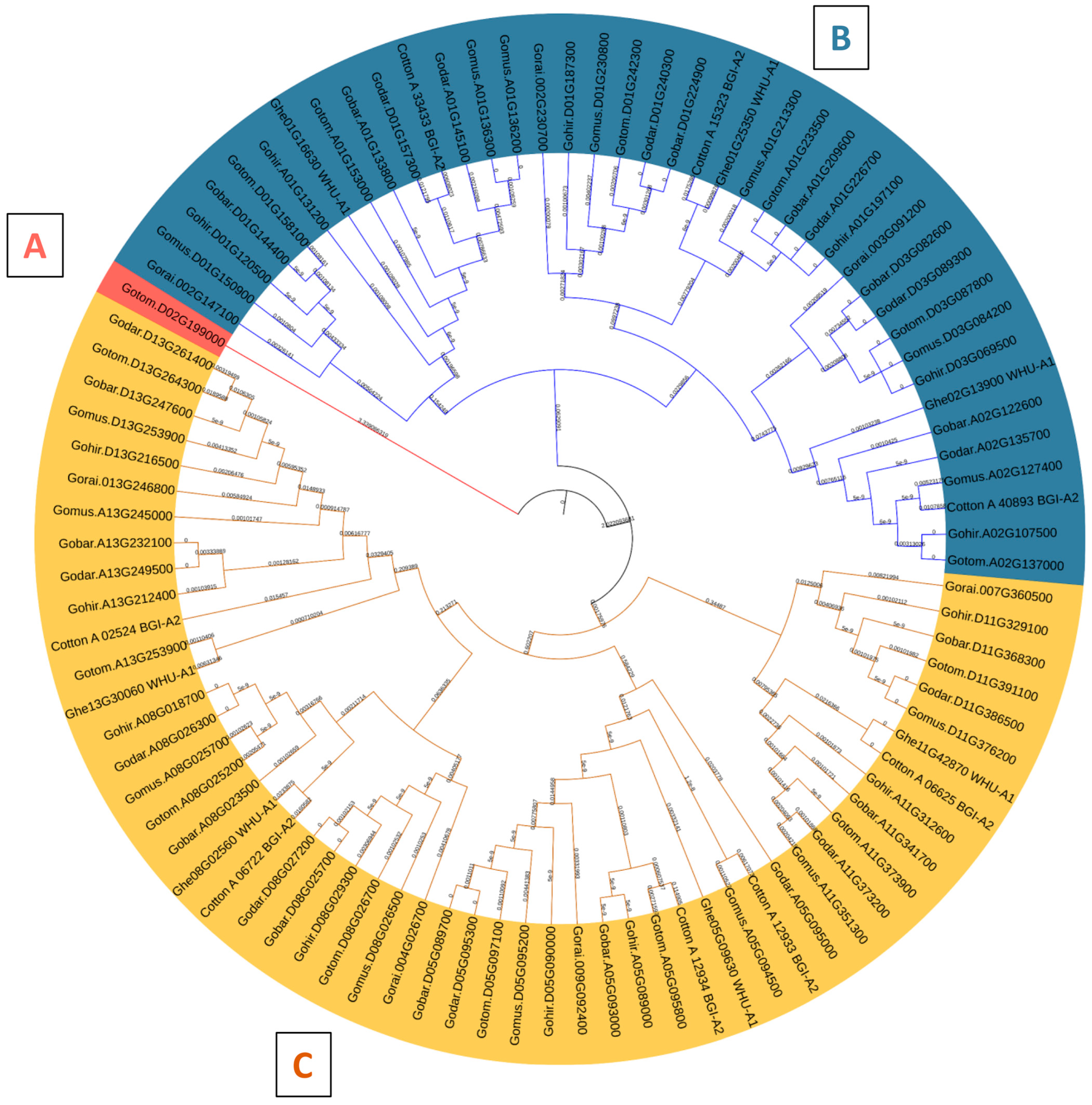
Figure 6.
Phylogenetic analysis depicting the evolutionary relationships of the VLN protein from Gossypium hirsutum alongside diverse plant species, organized into four distinct clades (A–D), providing valuable insights into the evolutionary trajectory of this gene.
Figure 6.
Phylogenetic analysis depicting the evolutionary relationships of the VLN protein from Gossypium hirsutum alongside diverse plant species, organized into four distinct clades (A–D), providing valuable insights into the evolutionary trajectory of this gene.
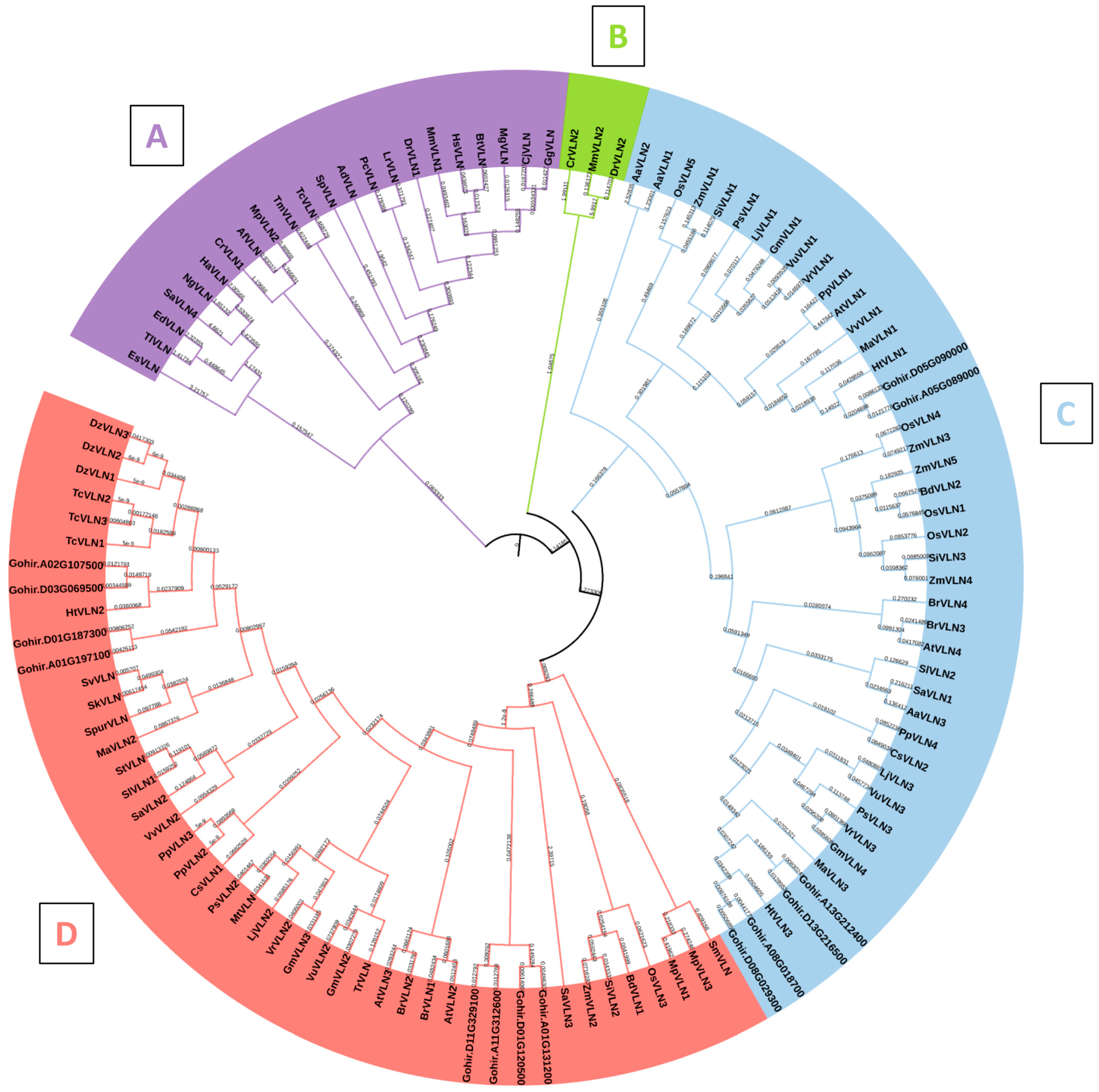
Figure 7.
Gene structure of GhVLNs, motif, and conservative domain analysis of GhVLNs. (A) Intra-species phylogenetic tree of GhVLNs. (B) The motif composition of GhVLNs, with conserved motifs in Gossypium VLN proteins indicated by colored boxes. (C) Conserved domains of GhVLNs, with colored boxes representing the conserved protein domain. (D) The gene structure of GhVLNs; green boxes, yellow boxes, and black lines represent the UTR, exons, and introns, respectively.
Figure 7.
Gene structure of GhVLNs, motif, and conservative domain analysis of GhVLNs. (A) Intra-species phylogenetic tree of GhVLNs. (B) The motif composition of GhVLNs, with conserved motifs in Gossypium VLN proteins indicated by colored boxes. (C) Conserved domains of GhVLNs, with colored boxes representing the conserved protein domain. (D) The gene structure of GhVLNs; green boxes, yellow boxes, and black lines represent the UTR, exons, and introns, respectively.
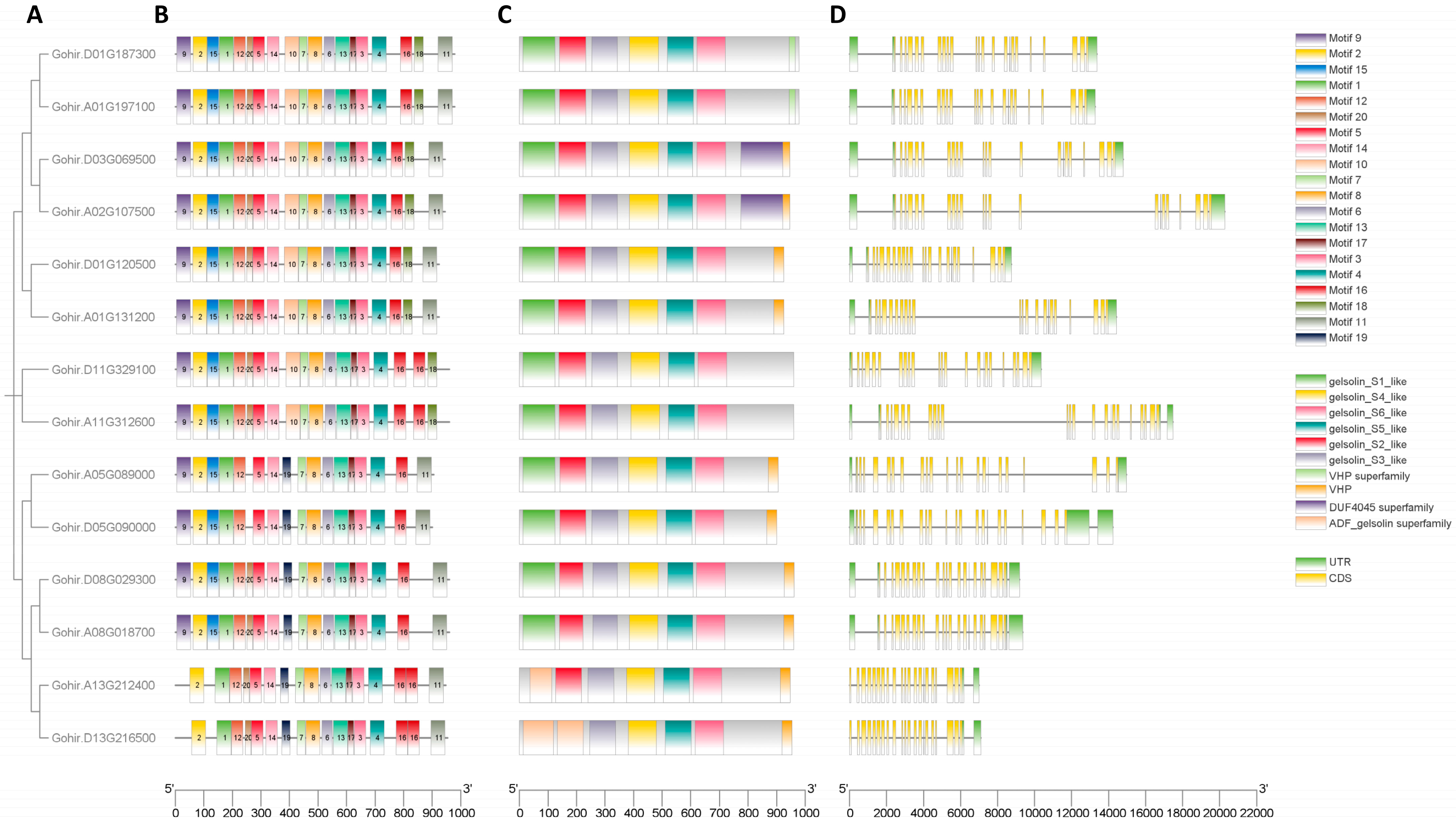
Figure 8.
Analysis of collinearity and chromosomal positions of GhVLNs: (A) We chose the model plant Arabidopsis thaliana for establishing collinearity with Gossypium hirsutum. In the graphical representation, the gray lines in the backdrop indicate collinear blocks shared between Arabidopsis thaliana and Gossypium hirsutum, whereas the red lines denote collinear blocks associated with VLN genes. (B) This diagram displays the physical positions of GhVLNs on the chromosomes, with the scale on the left indicating the genomic length in megabases (Mb).
Figure 8.
Analysis of collinearity and chromosomal positions of GhVLNs: (A) We chose the model plant Arabidopsis thaliana for establishing collinearity with Gossypium hirsutum. In the graphical representation, the gray lines in the backdrop indicate collinear blocks shared between Arabidopsis thaliana and Gossypium hirsutum, whereas the red lines denote collinear blocks associated with VLN genes. (B) This diagram displays the physical positions of GhVLNs on the chromosomes, with the scale on the left indicating the genomic length in megabases (Mb).

Figure 9.
Three-dimensional prediction of GhVLN proteins by SWISS-MODEL. The rainbow color scheme provides a visual representation of different structural features within the protein, aiding in the interpretation of its predicted three-dimensional structure.
Figure 9.
Three-dimensional prediction of GhVLN proteins by SWISS-MODEL. The rainbow color scheme provides a visual representation of different structural features within the protein, aiding in the interpretation of its predicted three-dimensional structure.
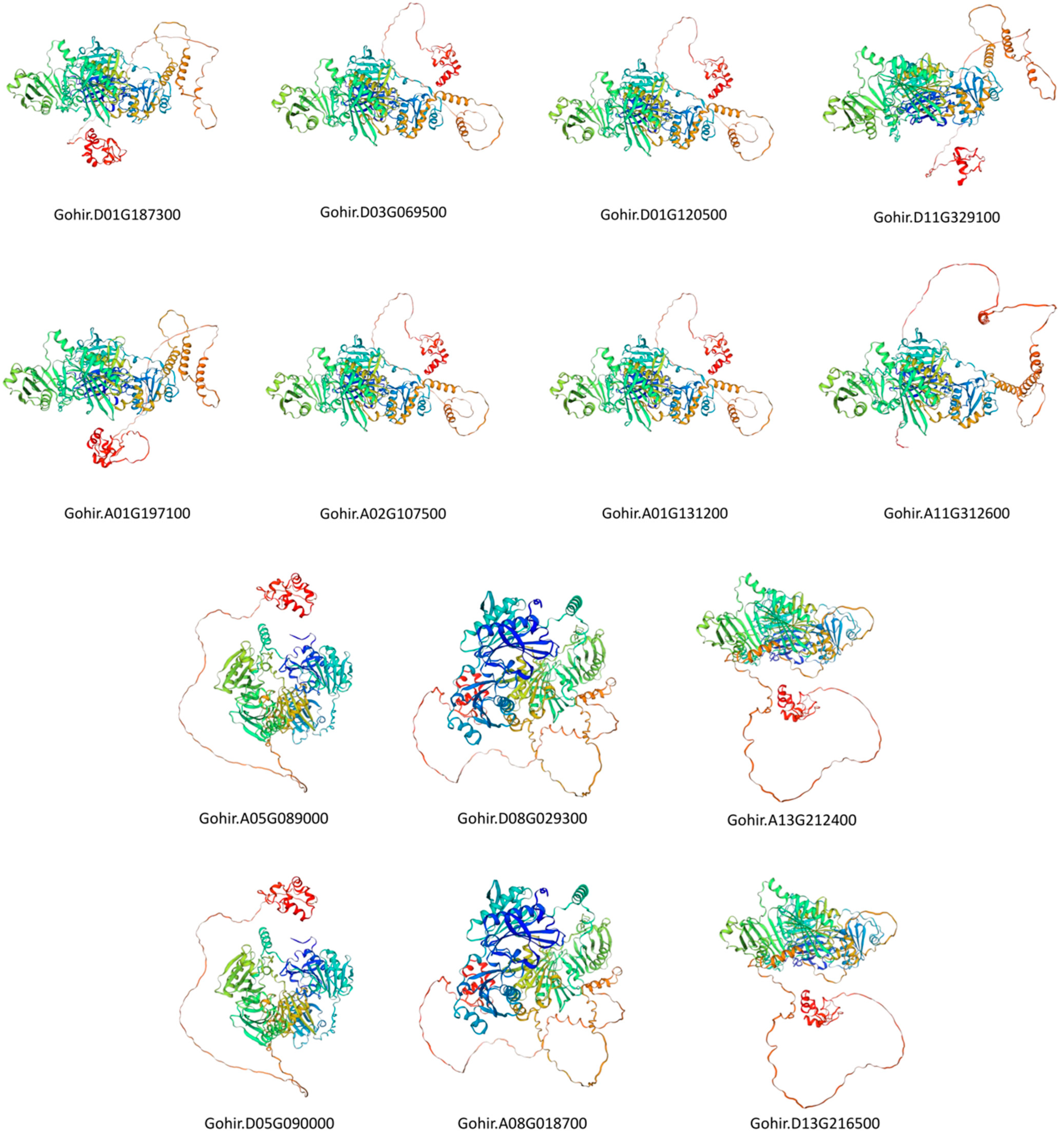
Figure 10.
Protein–protein interactions among GhVLN proteins, where each node of different color represents an individual protein, and each edge signifies an interaction. The red loci represent the query GhVLN protein. Edges in the visualization are assigned colors corresponding to different types of evidence: known interactions from curated databases and experimentally determined interactions are depicted by cyan and pink lines, respectively. Predicted interactions derived from gene neighborhood, gene fusions, and gene co-occurrence are represented by green, red, and blue lines. Additionally, other evidence types obtained through text-mining, co-expression, and protein homology are indicated by dark yellow, black, and violet lines, respectively.
Figure 10.
Protein–protein interactions among GhVLN proteins, where each node of different color represents an individual protein, and each edge signifies an interaction. The red loci represent the query GhVLN protein. Edges in the visualization are assigned colors corresponding to different types of evidence: known interactions from curated databases and experimentally determined interactions are depicted by cyan and pink lines, respectively. Predicted interactions derived from gene neighborhood, gene fusions, and gene co-occurrence are represented by green, red, and blue lines. Additionally, other evidence types obtained through text-mining, co-expression, and protein homology are indicated by dark yellow, black, and violet lines, respectively.

Figure 11.
Cis-element analysis of (A) environmental stress responses, (B) hormonal biological processes, (C) meristem expression, circadian control, endosperm expression, and regulation of flavonoid biosynthetic genes.
Figure 11.
Cis-element analysis of (A) environmental stress responses, (B) hormonal biological processes, (C) meristem expression, circadian control, endosperm expression, and regulation of flavonoid biosynthetic genes.

Figure 12.
Expression profiles of GhVLNs in different tissues and organs in Gossypium hirsutum represented on a heatmap through TBtools. (A) fiber and (B) ovule at different (dpa), and (C) expression of GHVLNs in different tissues.
Figure 12.
Expression profiles of GhVLNs in different tissues and organs in Gossypium hirsutum represented on a heatmap through TBtools. (A) fiber and (B) ovule at different (dpa), and (C) expression of GHVLNs in different tissues.
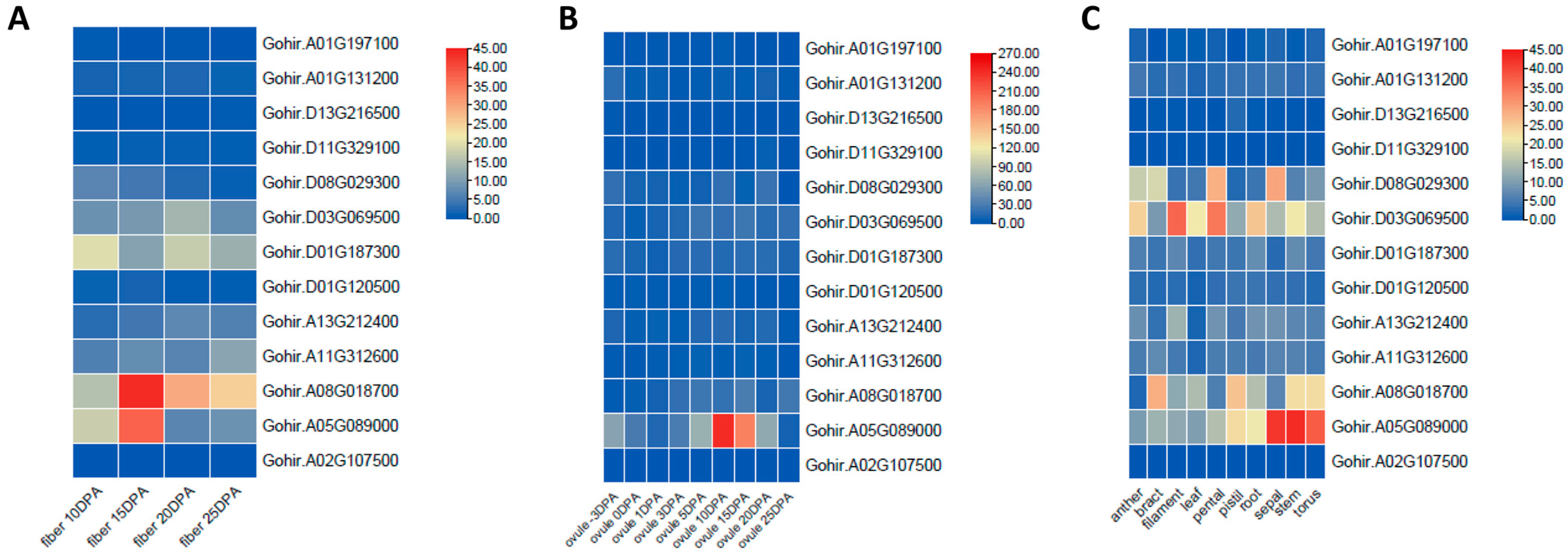
Figure 13.
Expression patterns of GhVLN genes in response to different stresses analyzed from FPKM values at five time points (1, 3, 6, 12, and 24 h) after stress treatment. (A) Cold stress; (B) drought stress; (C) Salt stress; and (D) Heat stress.
Figure 13.
Expression patterns of GhVLN genes in response to different stresses analyzed from FPKM values at five time points (1, 3, 6, 12, and 24 h) after stress treatment. (A) Cold stress; (B) drought stress; (C) Salt stress; and (D) Heat stress.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Characteristics of the 20 conserved motifs in GhVLNs.
| Motif | Motif Sequences | Length (A.A) | Domain |
|---|---|---|---|
| 1 | VPFARSSLNHDDVFILDTQNKIYQFNGANSNIQERAKALEVVQFJKEKYH | 50 | ADF_gelsolin super family |
| 2 | YDIHFWJGKDTSQDEAGAAAIKTVELDAVLGGRAVQHRELQGHESDKFLS | 50 | ADF_gelsolin super family |
| 3 | FKVEEVYNFSQDDLLTEDILILDTHAEVFVWVGQCVDTKEK | 41 | ADF_gelsolin super family |
| 4 | SLEGLSPEVPJYKVTEGNEPCFFTTFFSWDSTKATVHGNSFQKKLALLFG | 50 | ADF_gelsolin super family |
| 5 | KGLLENNKCYLLDCGAEVFVWVGRNTQVEERKAASQAAEDF | 41 | ADF_gelsolin super family |
| 6 | LFRISGTSPHNMKAJQVDAVATSLNSSECFJLQSGSS | 37 | ADF_gelsolin super family |
| 7 | DYFLCCWIGKDSIEEDQKTAVRLANKMVN | 29 | Not identified |
| 8 | KGRPVQGRVFEGKEPPQFIAJFQPMVVLKGGLSAGYKKSIAEKGJTDETY | 50 | ADF_gelsolin super family |
| 9 | KVLDPAFQGAGQKPGTEIWRIENFQPVPLPKSDYGKFYMGDSYIVLQTTP | 50 | ADF_gelsolin super family |
| 10 | PPLLEGGGKMEVWCINGSAKTPLPKEDIGKFYSGDCYIVLYTYHSGERKE | 50 | ADF_gelsolin super family |
| 11 | YERLKASSTBPVTGIDVKRREAYLSDEEFKEKFGMEKEAFYKLPKWKQBK | 50 | Villin-headpiece domain |
| 12 | GTCEVAIVEDGKLDTESDSGEFWVLFGGFAPJPKKTASEDD | 41 | Not identified |
| 13 | WHGNQSTYEQQQLVARVAEFJKPGVQLKHAKEGSESNAFWSALGGKTEYT | 50 | ADF_gelsolin super family |
| 14 | RITRVIZGYETNSFKSKFDSWPQGSNAPGGEEGRGKVAALL | 41 | Not identified |
| 15 | YFKPCIIPLEGGVASGFKKPEEEEFETRLYVCRGKRVVKLK | 41 | Not identified |
| 16 | GPRQRAPALAALASAFNPSSASKTSAPKPVSRKQGSQRAAA | 41 | Not identified |
| 17 | EKESSEIVRDPHLFTFSFNKG | 21 | Not identified |
| 18 | TAEKKKQSPDGSPIKSTSSTPAVTSPPTEAKS | 32 | Not identified |
| 19 | KEEPQPYIDCTGNLQVWRVNGQEKVLLPA | 29 | ADF_gelsolin super family |
| 20 | TPAKLYSITDGEVKPVEGELS | 21 | Not identified |
Table 2.
Ka/Ks values for GhVLN collinear gene pairs.
| Locus_1 | Locus_2 | Ka | Ks | Ka/Ks | Time (MYA) | Synteny | Selection |
|---|---|---|---|---|---|---|---|
| Gohir.D01G187300 | Gohir.A01G197100 | 0.006674869 | 0.028485622 | 0.234324135 | 1.56514409 | D01/A01 | Purifying |
| Gohir.D03G069500 | Gohir.A02G107500 | 0.008274678 | 0.054163656 | 0.152771775 | 2.976025047 | D03/A02 | Purifying |
| Gohir.D01G120500 | Gohir.A01G131200 | 0.005862153 | 0.046331088 | 0.126527414 | 2.545664167 | D01/A01 | Purifying |
| Gohir.D11G329100 | Gohir.A11G312600 | 0.013325515 | 0.043621315 | 0.305481738 | 2.396775561 | D11/A11 | Purifying |
| Gohir.A05G089000 | Gohir.D05G090000 | 0.010134474 | 0.03314541 | 0.305757995 | 1.821176385 | A05/D05 | Purifying |
| Gohir.D08G029300 | Gohir.A08G018700 | 0.004066581 | 0.042012538 | 0.096794467 | 2.308381209 | D08/A08 | Purifying |
| Gohir.A13G212400 | Gohir.D13G216500 | 0.011020325 | 0.057520859 | 0.191588331 | 3.160486746 | A13/D13 | Purifying |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Deep, A.; Pandey, D.K. Genome-Wide Analysis of VILLIN Gene Family Associated with Stress Responses in Cotton (Gossypium spp.). Curr. Issues Mol. Biol. 2024, 46, 2278-2300. https://doi.org/10.3390/cimb46030146
AMA Style
Deep A, Pandey DK. Genome-Wide Analysis of VILLIN Gene Family Associated with Stress Responses in Cotton (Gossypium spp.). Current Issues in Molecular Biology. 2024; 46(3):2278-2300. https://doi.org/10.3390/cimb46030146
Chicago/Turabian StyleDeep, Akash, and Dhananjay K. Pandey. 2024. "Genome-Wide Analysis of VILLIN Gene Family Associated with Stress Responses in Cotton (Gossypium spp.)" Current Issues in Molecular Biology 46, no. 3: 2278-2300. https://doi.org/10.3390/cimb46030146