Suvanine Sesterterpenes from a Tropical Sponge Coscinoderma sp. Inhibit Isocitrate Lyase in the Glyoxylate Cycle

Abstract
:1. Introduction
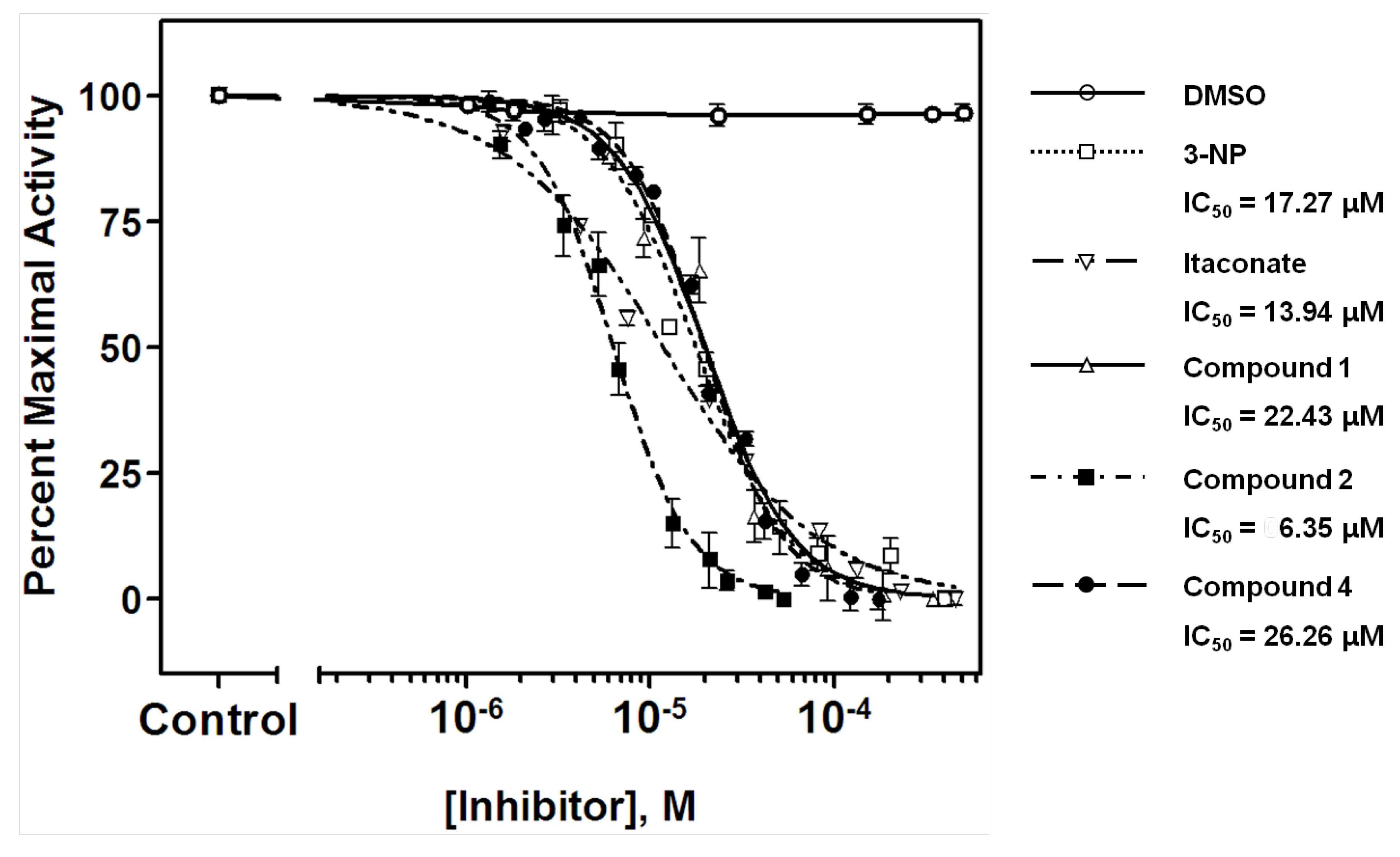
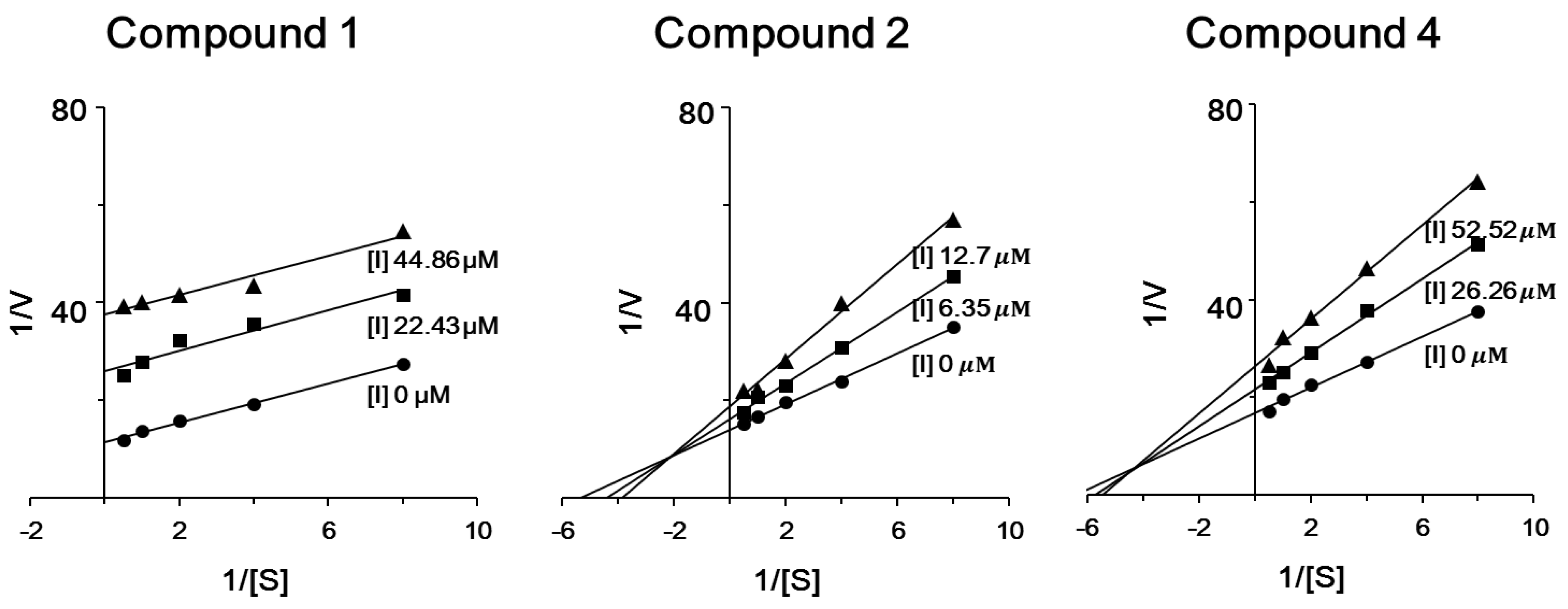
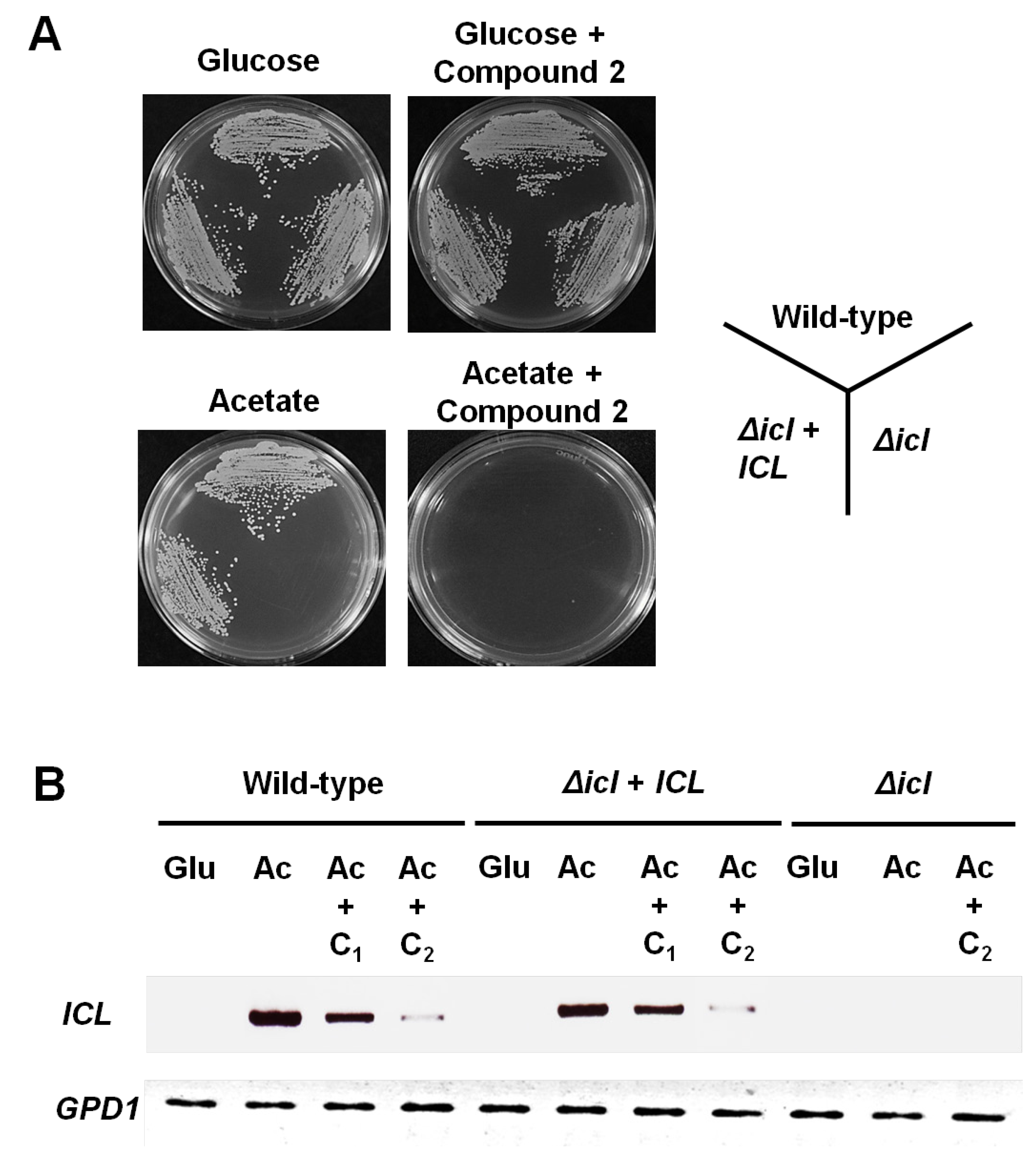
2. Results and Discussion



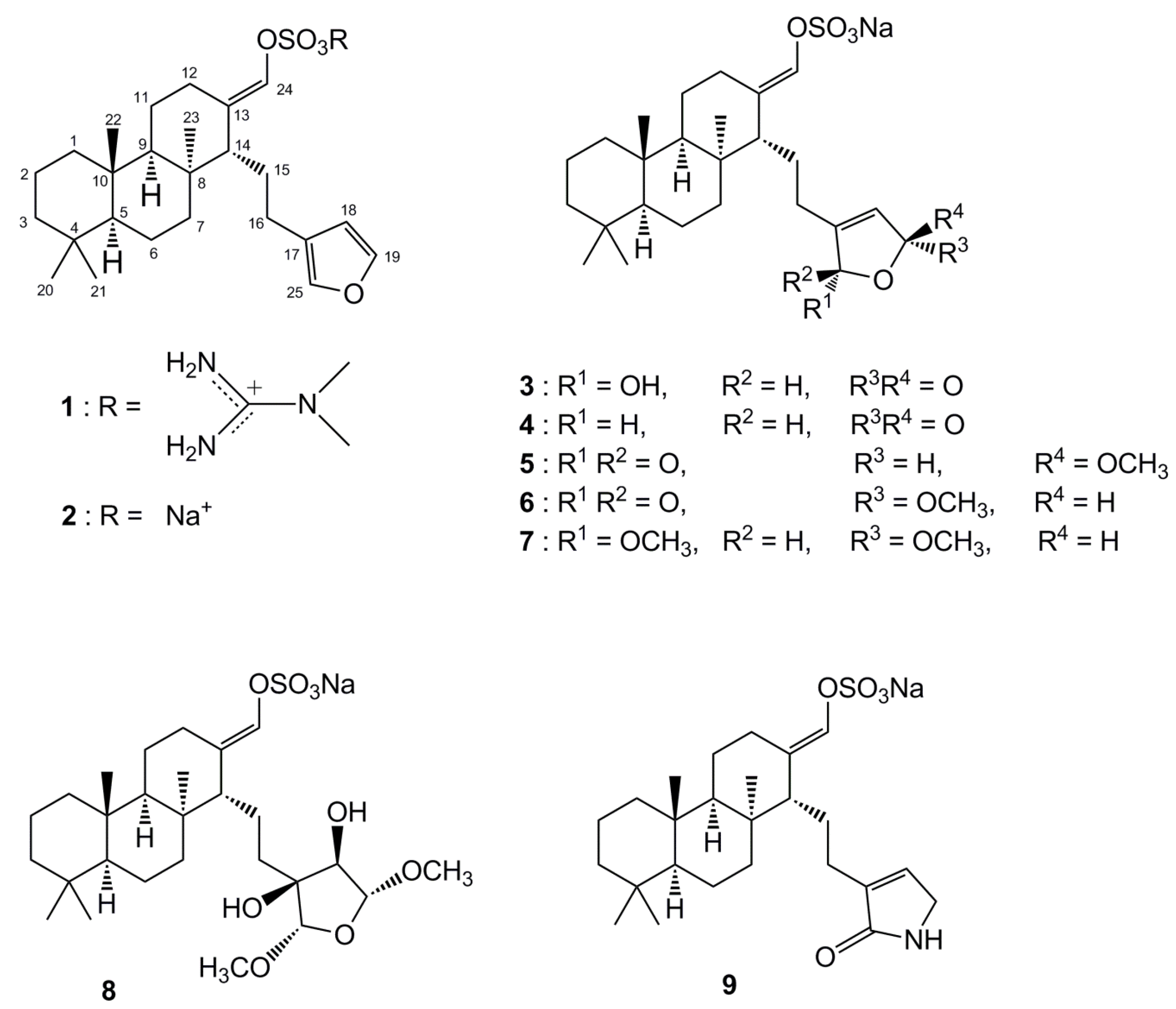{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | ICL IC50, µM (µg/mL) | MIC (µg/mL) | |
| Glucose | Acetate | ||
| 1 | 22.43 ± 1.49 (12.03 ± 0.80) | 100 | 25 |
| 2 | 6.35 ± 1.37 (3.00 ± 0.65) | 100 | 12.5 |
| 3 | 56.19 ± 7.10 (28.33 ± 3.58) | >100 | >100 |
| 4 | 26.26 ± 3.69 (12.82 ± 1.80) | >100 | 100 |
| 5 | 50.23 ± 6.27 (26.03 ± 3.25) | >100 | >100 |
| 6 | 96.15 ± 1.54 (49.83 ± 0.80) | >100 | 100 |
| 7 | 59.10 ± 3.45 (27.30 ± 1.85) | >100 | >100 |
| 8 | 67.64 ± 4.88 (38.44 ± 2.77) | >100 | >100 |
| 9 | 59.15 ± 1.11 (28.82 ± 0.54) | >100 | >100 |
| 3-NP | 17.27 ± 1.04 (2.06 ± 0.12) | >100 | >100 |
| Itaconate | 13.94 ± 0.64 (1.66 ± 0.08) | >100 | >100 |
| Amph B | ND | 1.56 | 0.39 |


3. Experimental Section
3.1. General Experimental Procedure
3.2. C. albicans Strains and Growth Media
3.3. Preparation of Recombinant ICL
3.4. ICL Inhibitor Potency Determination
3.5. In Vitro Growth Assays
3.6. Semi-Quantitative RT-PCR Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Vanni, P.; Giachetti, E.; Pinzuati, G.; McFadden, B.A. Comparative structure, function and regulation of isocitrate lyase, an important assimilatory enzyme. Comp. Biochem. Physiol. B 1990, 95, 431–458. [Google Scholar]
- Dunn, M.F.; Ramirez-Trujillo, J.A.; Hernandez-Lucas, I. Major roles of isocitrate lyase and malate synthase in bacterial and fungal pathogenesis. Microbiology 2009, 155, 3166–3175. [Google Scholar] [CrossRef]
- Kunze, M.; Pracharoenwattana, I.; Smith, S.M.; Hartig, A. A central role for the peroxisomal membrane in glyoxylate cycle function. Biochim. Biophys. Acta 2006, 1763, 1441–1452. [Google Scholar] [CrossRef]
- Strijbis, K.; Distel, B. Intracellular acetyl unit transport in fungal carbon metabolism. Eukaryot. Cell 2010, 9, 1809–1815. [Google Scholar] [CrossRef]
- McKinney, J.D.; Honer Zu Bentrup, K.; Munoz-Elias, E.J.; Miczak, A.; Chen, B.; Chan, W.T.; Swenson, D.; Sacchettini, J.C.; Jacobs, W.R., Jr.; Russell, D.G. Persistence of Mycobacterium tuberculosis in macrophages and mice requires the glyoxylate shunt enzyme isocitrate lyase. Nature 2000, 406, 735–738. [Google Scholar] [CrossRef]
- Munoz-Elias, E.J.; McKinney, J.D. Mycobacterium tuberculosis isocitrate lyases 1 and 2 are jointly required for in vivo growth and virulence. Nat. Med. 2005, 11, 638–644. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Thornton, C.R.; Kershaw, M.J.; Debao, L.; Talbot, N.J. The glyoxylate cycle is required for temporal regulation of virulence by the plant pathogenic fungus Magnaporthe grisea. Mol. Microbiol. 2003, 47, 1601–1612. [Google Scholar] [CrossRef]
- Lorenz, M.C.; Fink, G.R. The glyoxylate cycle is required for fungal virulence. Nature 2001, 412, 83–86. [Google Scholar] [CrossRef]
- Ramirez, M.A.; Lorenz, M.C. Mutations in alternative carbon utilization pathways in Candida albicans attenuate virulence and confer pleiotropic phenotypes. Eukaryot. Cell 2007, 6, 280–290. [Google Scholar] [CrossRef]
- Goldstein, A.L.; McCusker, J.H. Development of Saccharomyces cerevisiae as a model pathogen. A system for the genetic identification of gene products required for survival in the mammalian host environment. Genetics 2001, 159, 499–513. [Google Scholar]
- Sharma, R.; Das, O.; Damle, S.G.; Sharma, A.K. Isocitrate lyase: A potential target for anti-tubercular drugs. Recent Pat. Inflamm. Allergy Drug Discov. 2013, 7, 114–123. [Google Scholar] [CrossRef]
- Kratky, M.; Vinsova, J. Advances in mycobacterial isocitrate lyase targeting and inhibitors. Curr. Med. Chem. 2012, 19, 6126–6137. [Google Scholar] [CrossRef]
- Lee, D.; Shin, J.; Yoon, K.M.; Kim, T.I.; Lee, S.H.; Lee, H.S.; Oh, K.B. Inhibition of Candida albicans isocitrate lyase activity by sesterterpene sulfates from the tropical sponge Dysidea sp. Bioorg. Med. Chem. Lett. 2008, 18, 5377–5380. [Google Scholar] [CrossRef]
- Lee, H.S.; Lee, T.H.; Lee, J.H.; Chae, C.S.; Chung, S.C.; Shin, D.S.; Shin, J.; Oh, K.B. Inhibition of the pathogenicity of Magnaporthe grisea by bromophenols, isocitrate lyase inhibitors, from the red alga Odonthalia corymbifera. J. Argic. Food. Chem. 2007, 55, 6923–6928. [Google Scholar] [CrossRef]
- Kernan, M.R.; Faulkner, D.J. Sesterterpene sulfates from a sponge of the family Halichondriidae. J. Org. Chem. 1988, 53, 4574–4578. [Google Scholar] [CrossRef]
- Shin, D.S.; Lee, T.H.; Lee, H.S.; Shin, J.; Oh, K.B. Inhibition of infection of the rice blast fungus by halisulfate 1, an isocitrate lyase inhibitor. FEMS Microbiol. Lett. 2007, 272, 43–47. [Google Scholar] [CrossRef]
- De Marino, S.; Festa, C.; D’Auria, M.V.; Bourguet-Kondracki, M.L.; Petek, S.; Debitus, C.; Andres, R.M.; Terencio, M.C.; Paya, M.; Zampella, A. Coscinolactams A and B: New nitrogen-containing sesterterpenoids from the marine sponge Coscinoderma mathewsi exerting anti-inflammatory properties. Tetrahedron 2009, 65, 2905–2909. [Google Scholar]
- Kimura, J.; Ishizuka, E.; Nakao, Y.; Yoshida, W.Y.; Scheuer, P.J.; Kelly-Borges, M. Isolation of 1-methylherbipoline salts of halisulfate-1 and of suvanine as serine protease inhibitors from a marine sponge, Coscinoderma mathewsi. J. Nat. Prod. 1998, 61, 248–250. [Google Scholar] [CrossRef]
- Loukaci, A.; Le Saout, I.; Samadi, M.; Leclerc, S.; Damiens, E.; Meijer, L.; Debitus, C.; Guyot, M. Coscinosulfate, a CDC25 phosphatase inhibitor from the sponge Coscinoderma mathewsi. Bioorg. Med. Chem. 2001, 9, 3049–3054. [Google Scholar] [CrossRef]
- Cassiano, C.; Monti, M.C.; Festa, C.; Zampella, A.; Riccio, R.; Casapullo, A. Chemical proteomics reveals heat shock protein 60 to be the main cellular target of the marine bioactive sesterterpene suvanine. ChemBioChem 2012, 13, 1953–1958. [Google Scholar] [CrossRef]
- Bae, J.; Jeon, J.; Lee, Y.J.; Lee, H.S.; Sim, C.J.; Oh, K.B.; Shin, J. Sesterterpenes from the tropical sponge Coscinoderma sp. J. Nat. Prod. 2011, 74, 1805–1811. [Google Scholar] [CrossRef]
- Shin, D.S.; Kim, S.; Yang, H.C.; Oh, K.B. Cloning and expression of isocitrate lyase, a key enzyme of the glyoxylate cycle, of Candida albicans for development of antifungal drugs. J. Microbiol. Biotechnol. 2005, 15, 652–655. [Google Scholar]
- Dixon, G.H.; Kornberg, H.L. Assay methods for key enzymes of the glyoxalate assay. Biochem. J. 1959, 72, 3P. [Google Scholar]
- Hautzel, R.; Anke, H.; Sheldrick, W.S. Mycenon, a new metabolite from a Mycena species TA 87202 (Basidiomycetes) as an inhibitor of isocitrate lyase. J. Antibiot. 1990, 43, 1240–1244. [Google Scholar] [CrossRef]
- Miller, B.; Friedman, A.J.; Choi, H.; Hogan, J.; McCammon, J.A.; Hook, V.; Gerwick, W.H. The marine cyanobacterial metabolite gallinamide A is a potent and selective inhibitor of human cathepsin L. J. Nat. Prod. 2014, 77, 92–99. [Google Scholar] [CrossRef]
- Sharma, V.; Sharma, S.; Hoener Zu Bentrup, K.; McKinney, J.D.; Russell, D.G.; Jacobs, W.R., Jr.; Sacchettini, J.C. Structure of isocitrate lyase, a persistence factor of Mycobacterium tuberculosis. Nat. Struct. Biol. 2000, 7, 663–668. [Google Scholar] [CrossRef]
- Lineweaver, H.; Burk, D. The determination of enzyme dissociation constants. J. Am. Chem. Soc. 1934, 56, 658–666. [Google Scholar] [CrossRef]
- Pedemonte, H.C.; Kaplan, J.H. Inhibition and derivatization of the renal Na,K-ATPase by Dihydro-4,4′-diisothiocyanatostilbene-2,2′-disulfonate. Biochemistry 1988, 27, 7966–7973. [Google Scholar] [CrossRef]
- Jacob, C. A scent of therapy: pharmacological implications of natural products containing redox-active sulfur atoms. Nat. Prod. Rep. 2006, 23, 851–863. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-H.; Won, T.H.; Kim, H.; Ahn, C.-H.; Shin, J.; Oh, K.-B. Suvanine Sesterterpenes from a Tropical Sponge Coscinoderma sp. Inhibit Isocitrate Lyase in the Glyoxylate Cycle. Mar. Drugs 2014, 12, 5148-5159. https://doi.org/10.3390/md12105148
Lee S-H, Won TH, Kim H, Ahn C-H, Shin J, Oh K-B. Suvanine Sesterterpenes from a Tropical Sponge Coscinoderma sp. Inhibit Isocitrate Lyase in the Glyoxylate Cycle. Marine Drugs. 2014; 12(10):5148-5159. https://doi.org/10.3390/md12105148
Chicago/Turabian StyleLee, So-Hyoung, Tae Hyung Won, Heegyu Kim, Chan-Hong Ahn, Jongheon Shin, and Ki-Bong Oh. 2014. "Suvanine Sesterterpenes from a Tropical Sponge Coscinoderma sp. Inhibit Isocitrate Lyase in the Glyoxylate Cycle" Marine Drugs 12, no. 10: 5148-5159. https://doi.org/10.3390/md12105148