Anti-Inflammatory Activity and Mechanism of a Lipid Extract from Hard-Shelled Mussel (Mytilus Coruscus) on Chronic Arthritis in Rats


Abstract
:1. Introduction
2. Results and Discussion
2.1. Effect of HMLE on Chronic Arthritis in AIA and CIA Rats

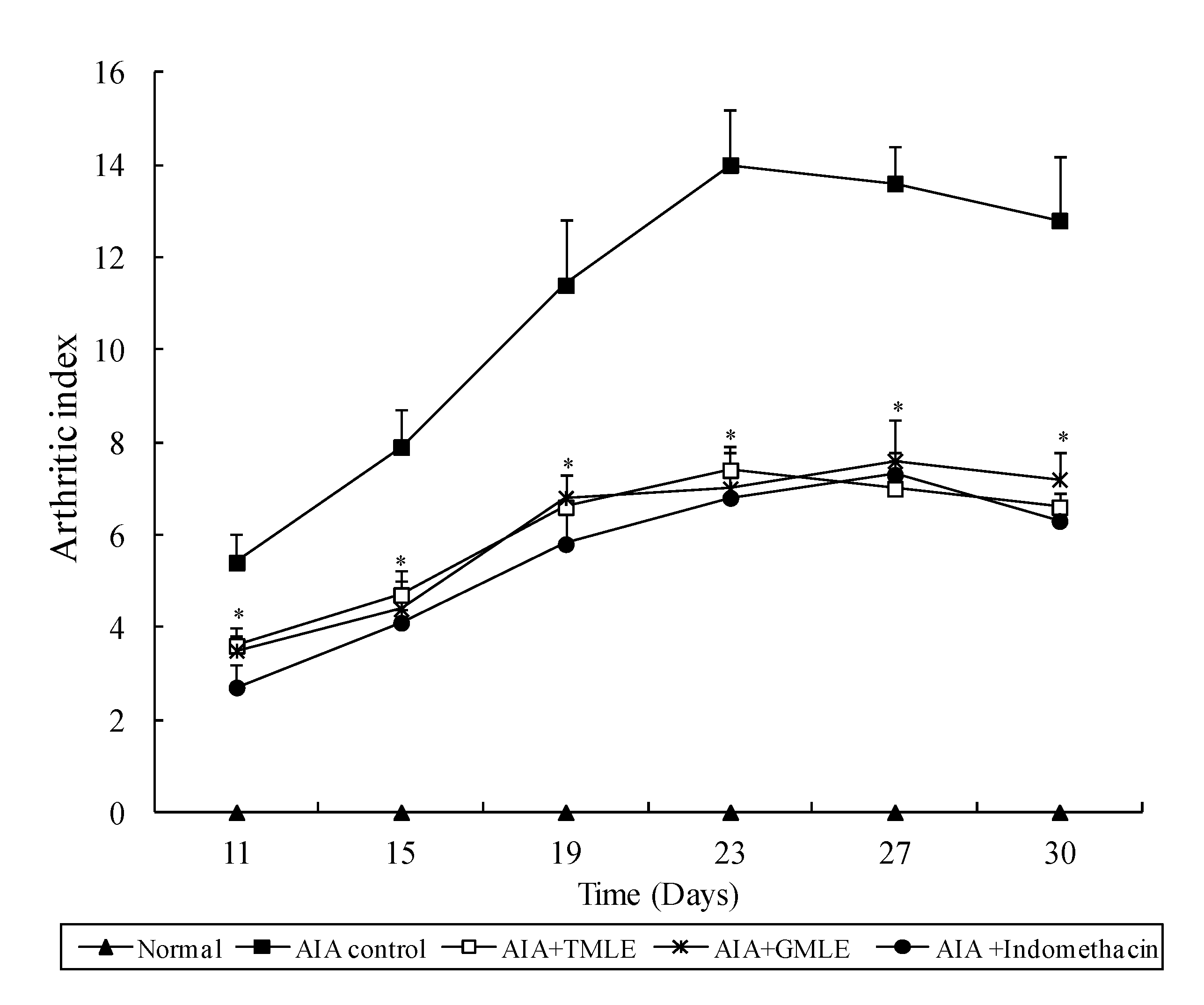{kind=link}
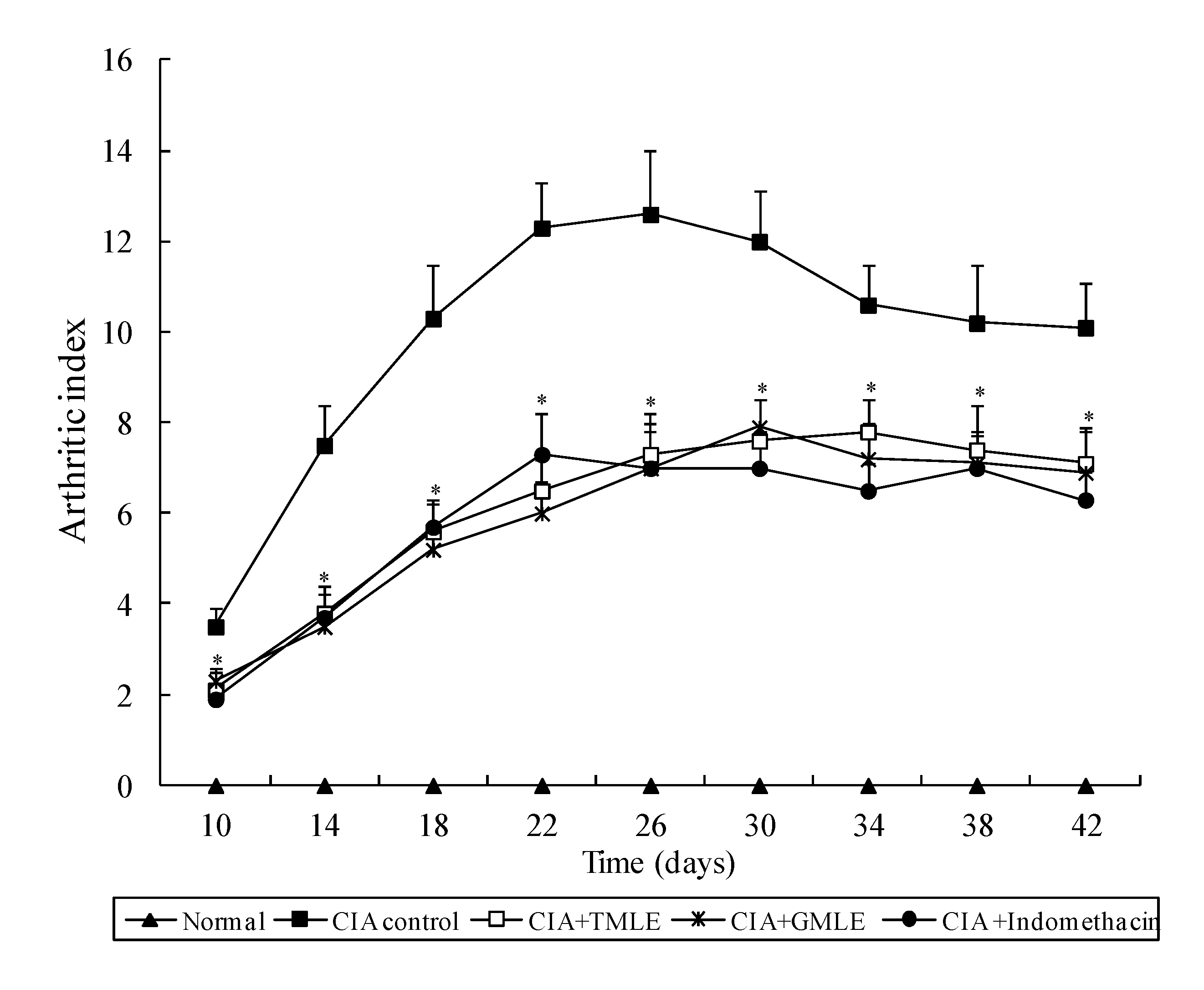{kind=link}
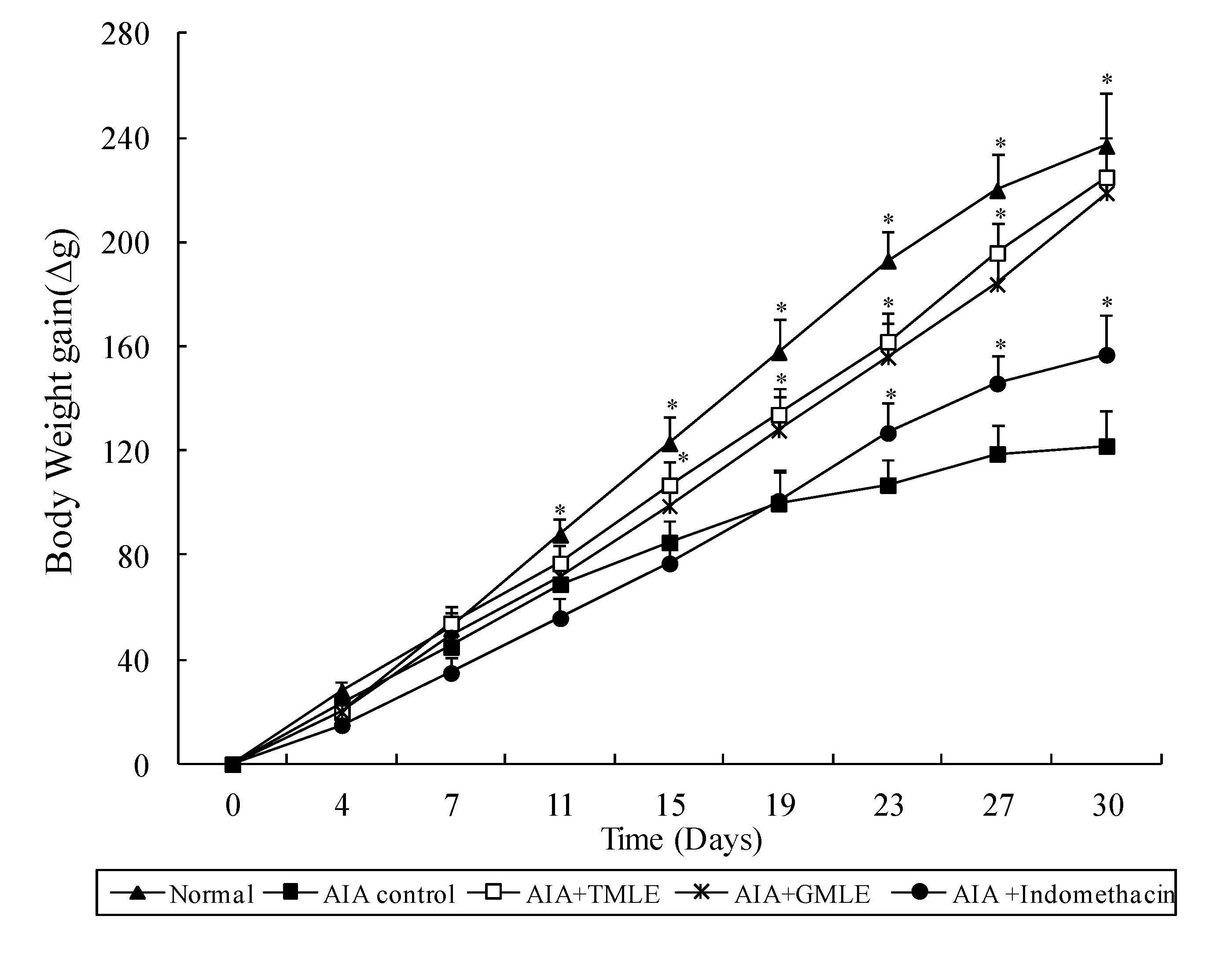{kind=link}
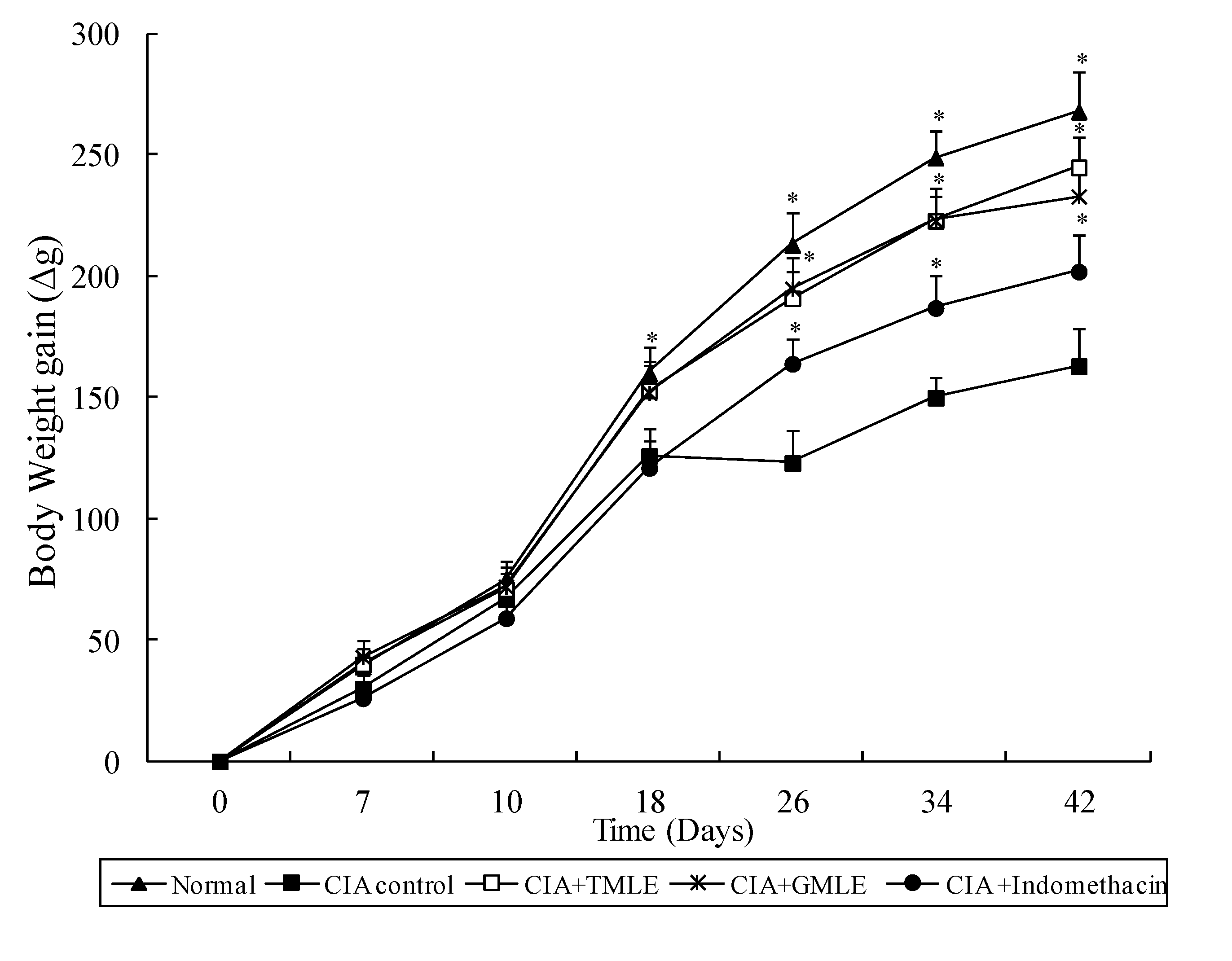{kind=link}
{kind=link}
| Groups | Number | Dose (mg/kg) | Left Hind Paw Swelling (ΔmL) | |||||
|---|---|---|---|---|---|---|---|---|
| Day 11 | Day 15 | Day 19 | Day 23 | Day 27 | Day 30 | |||
| Normal | n = 10 | - | 0.12 ± 0.01 c | 0.15 ± 0.02 c | 0.20 ± 0.02 c | 0.25 ± 0.01 c | 0.25 ± 0.00 c | 0.28 ± 0.02 c |
| AIA control | n = 10 | - | 0.79 ± 0.20 a | 1.30 ± 0.24 a | 1.93 ± 0.35a | 1.45 ± 0.42 a | 0.98 ± 0.26 a | 0.73 ± 0.17 a |
| AIA + HMLE | n = 8 | 100 | 0.43 ± 0.11 b | 0.75 ± 0.20 b | 0.98 ± 0.25 b | 0.80 ± 0.10 b | 0.56 ± 0.07 b | 0.40 ± 0.14 b |
| AIA + GMLE | n = 9 | 100 | 0.46 ± 0.14 b | 0.71 ± 0.17 b | 1.06 ± 0.22 b | 0.76 ± 0.12 b | 0.53 ± 0.14 b | 0.36 ± 0.08 b |
| AIA + Indomethacin | n = 7 | 1.5 | 0.39 ± 0.11 b | 0.65 ± 0.13 b | 0.85 ± 0.21 b | 0.75 ± 0.14 b | 0.48 ± 0.18 b | 0.39 ± 0.10 b |
| P-value | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | ||
| Groups | Number | Dose (mg/kg) | Left Hind Paw Swelling (ΔmL) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Day 10 | Day 14 | Day 18 | Day 22 | Day 26 | Day 30 | Day 34 | Day 38 | Day 42 | |||
| Normal | n = 10 | - | 0.12 ± 0.00 b | 0.13 ± 0.01 c | 0.16 ± 0.01 c | 0.22 ± 0.02 c | 0.26 ± 0.01 c | 0.29 ± 0.02 c | 0.30 ± 0.03 d | 0.30 ± 0.01 c | 0.32 ± 0.02 c |
| CIA control | n = 11 | - | 0.25 ± 0.07 a | 0.68 ± 0.13 a | 0.97 ± 0.17 a | 1.27 ± 0.25 a | 1.20 ± 0.20 a | 1.25 ± 0.27 a | 1.03 ± 0.18 a | 1.07 ± 0.24 a | 0.96 ± 0.15 a |
| CIA + HMLE | n = 12 | 100 | 0.16 ± 0.06 ab | 0.35 ± 0.07 b | 0.51 ± 0.09 b | 0.58 ± 0.10 b | 0.65 ± 0.12 b | 0.69 ± 0.10 b | 0.67 ± 0.14 bc | 0.64 ± 0.13 b | 0.60 ± 0.11 b |
| CIA + GMLE | n = 9 | 100 | 0.14 ± 0.03 b | 0.37 ± 0.06 b | 0.46 ± 0.07 b | 0.54 ± 0.05 b | 0.62 ± 0.08 b | 0.73 ± 0.14 b | 0.70 ± 0.13 b | 0.63 ± 0.10 b | 0.56 ± 0.09 b |
| CIA + Indomethacin | n = 10 | 1.5 | 0.10 ± 0.04 b | 0.36 ± 0.08 b | 0.51 ± 0.11 b | 0.67 ± 0.12 b | 0.64 ± 0.12 b | 0.60 ± 0.08 b | 0.52 ± 0.09 c | 0.49 ± 0.08 b | 0.51 ± 0.07 b |
| P-value | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | ||




2.2. Effect of HMLE on the Levels of Inflammatory Mediators in the Serum of AIA and CIA Rats
| Groups | Number | Dose (mg/kg) | LTB4 (ng/L) | PGE2 (µg/L) | TXB2 (ng/L) |
|---|---|---|---|---|---|
| Normal | n = 10 | - | 10.37 ± 1.09 c | 1.03 ± 0.13 c | 38.64 ± 6.18 c |
| AIA control | n = 10 | - | 15.68 ± 1.42 a | 3.76 ± 0.37 a | 75.43 ± 10.75 a |
| AIA + HMLE | n = 8 | 100 | 11.86 ± 1.83 b | 2.38 ± 0.27 b | 41.77 ± 5.13 c |
| AIA + GMLE | n = 9 | 100 | 11.24 ± 1.25 bc | 2.03 ± 0.18 b | 48.42 ± 7.07 b |
| AIA + Indomethacin | n = 7 | 1.5 | 15.33 ± 1.72 a | 0.74 ± 0.15 d | 27.68 ± 4.10 d |
| P-value | 0.000 | 0.000 | 0.000 |
| Groups | Number | Dose (mg/kg) | LTB4 (ng/L) | PGE2 (µg/L) | TXB2 (ng/L) |
|---|---|---|---|---|---|
| Normal | n = 10 | - | 15.41 ± 1.37 c | 0.84 ± 0.13 c | 40.33 ± 4.51 d |
| CIA control | n = 11 | - | 28.26 ± 3.83 a | 3.26 ± 0.46 a | 104.15 ± 15.57 a |
| CIA + HMLE | n = 12 | 100 | 20.75 ± 3.16 b | 1.64 ± 0.23 b | 73.26 ± 9.27 b |
| CIA + GMLE | n = 9 | 100 | 18.42 ± 2.73 b | 1.82 ± 0.19 b | 58.41 ± 8.36 c |
| CIA + Indomethacin | n = 10 | 1.5 | 26.24 ± 3.02 a | 0.64 ± 0.11 d | 43.16 ± 4.26 d |
| P-value | 0.000 | 0.000 | 0.000 |
2.3. Effect of HMLE on the Levels of Inflammatory Cytokines in the Ankle Joint Synovial Fluids of AIA and CIA Rats
| Groups | Number | Dose (mg/kg) | IL-1β (ng/L) | IL-2 (ng/L) | IL-4 (ng/L) | IL-6 (ng/L) | IL-10 (ng/L) | IFN-γ (ng/L) | TNF-α (ng/L) |
|---|---|---|---|---|---|---|---|---|---|
| Normal | n = 10 | - | 29.71 ± 4.28 c | 5.23 ± 1.96 c | 20.36 ± 3.72 b | 56.16 ± 13.58 c | 11.35 ± 3.73 c | 15.83 ± 4.64 b | 49.71 ± 10.31 d |
| AIA control | n = 10 | - | 53.66 ± 10.66 a | 10.18 ± 2.74 a | 15.72 ± 3.55 b | 154.23 ± 39.63 a | 16.57 ± 5.24 c | 25.66 ± 5.83 a | 381.45 ± 89.45 a |
| AIA + HMLE | n = 8 | 100 | 40.37 ± 10.32 b | 9.45 ± 3.31 ab | 28.13 ± 6.48 a | 95.46 ± 26.73 b | 26.43 ± 7.36 b | 18.37 ± 4.36 b | 125.27 ± 28.37 b |
| AIA + GMLE | n = 9 | 100 | 38.16 ± 7.15 b | 10.61 ± 3.86 a | 27.29 ± 7.35 a | 89.37 ± 24.75 b | 24.21 ± 5.83 b | 16.21 ± 5.73 b | 100.35 ± 23.66 bc |
| AIA + Indomethacin | n = 7 | 1.5 | 38.42 ± 8.06 b | 7.48 ± 2.43 bc | 28.76 ± 6.76 a | 81.36 ± 17.26 b | 34.79 ± 7.21 a | 17.45 ± 3.94 b | 87.34 ± 15.84 c |
| P-value | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 |
| Groups | Number | Dose (mg/kg) | IL-1β (ng/L) | IL-2 (ng/L) | IL-4 (ng/L) | IL-6 (ng/L) | IL-10 (ng/L) | IFN-γ (ng/L) | TNF-α (ng/L) |
|---|---|---|---|---|---|---|---|---|---|
| Normal | n = 10 | - | 37.83 ± 9.43 d | 8.45 ± 2.68 b | 33.24 ± 5.25 a | 79.26 ± 15.46 c | 47.93 ± 10.71 c | 15.35 ± 3.68 b | 43.61 ± 7.89 c |
| CIA control | n = 11 | - | 90.61 ± 20.17 a | 17.46 ± 5.34 a | 20.67 ± 4.76 c | 168.55 ± 37.38 a | 78.35 ± 18.73 b | 27.61 ± 6.04 a | 162.29 ± 33.75 a |
| CIA + HMLE | n = 12 | 100 | 61.33 ± 15.35 b | 18.54 ± 4.79 a | 22.82 ± 5.33 c | 111.37 ± 25.45 b | 126.68 ± 30.26 a | 18.41 ± 5.43 b | 83.77 ± 18.38 b |
| CIA + GMLE | n = 9 | 100 | 57.75 ± 13.21 bc | 15.13 ± 4.72 a | 21.57 ± 5.69 c | 119.64 ± 28.72 b | 132.47 ± 28.66 a | 18.23 ± 4.93 b | 97.64 ± 20.56 b |
| CIA + Indomethacin | n = 10 | 1.5 | 47.28 ± 8.24 cd | 10.85 ± 3.53 b | 26.83 ± 4.97 b | 111.84 ± 20.25 b | 84.38 ± 18.54 b | 16.76 ± 3.16 b | 80.63 ± 20.72 b |
| P-value | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 |
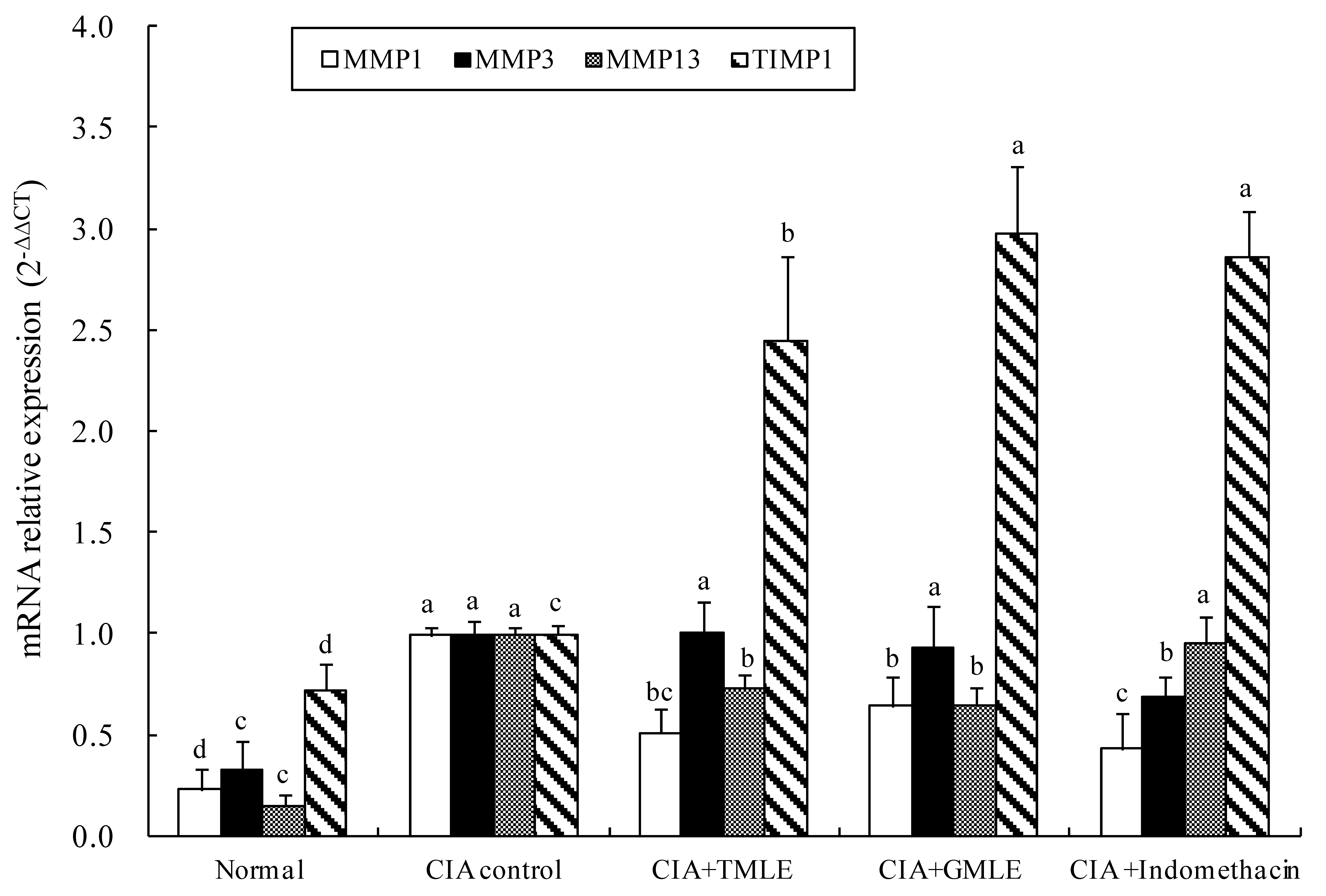
2.4. Effect of HMLE on the mRNA Expression Levels of Matrix Metalloproteinase in the Knee Joint Synovium Tissues of CIA Rats

2.5. Effect of HMLE on the Hepatic Enzyme Activities in the Serum of AIA Rats
| Groups | Number | Dose (mg/kg) | AST (U/L) | ALT (U/L) | ALP (U/L) |
|---|---|---|---|---|---|
| Normal | n = 10 | - | 48.74 ± 5.29 ab | 133.4 ± 21.2 c | 208.6 ± 31.7 c |
| AIA control | n = 10 | - | 45.68 ± 8.35 ab | 276.5 ± 35.3 b | 275.3 ± 36.5 b |
| AIA + HMLE | n = 8 | 100 | 43.86 ± 7.38 b | 161.4 ± 27.2 c | 217.7 ± 25.3 c |
| AIA + GMLE | n = 9 | 100 | 47.81 ± 7.97 ab | 148.2 ± 21.8 c | 196.4 ± 30.1 c |
| AIA + Indomethacin | n = 7 | 1.5 | 58.53 ± 10.12 a | 374.8 ± 51.6 a | 412.4 ± 60.3 a |
| P-value | 0.017 | 0.000 | 0.000 |
3. Experimental Section
3.1. Hard-Shelled Mussel Collection and HMLE Preparation
| Fatty Acid | Storage Time | P-value | |||
|---|---|---|---|---|---|
| 0 Days | 30 Days | 60 Days | 90 Days | ||
| C12:0 | 0.26 ± 0.13 b | 0.65 ± 0.09 a | 0.63 ± 0.17 a | 0.38 ± 0.11 b | 0.014 |
| C14:0 | 2.31 ± 0.19 | 2.43 ± 0.15 | 2.46 ± 0.22 | 2.37 ± 0.20 | 0.783 |
| C15:0 | 0.68 ± 0.09 | 0.75 ± 0.06 | 0.79 ± 0.13 | 0.64 ± 0.14 | 0.391 |
| C16:0 | 20.05 ± 1.35 | 20.90 ± 1.17 | 21.54 ± 1.23 | 22.03 ± 1.50 | 0.350 |
| C17:0 | 2.24 ± 0.12 b | 2.53 ± 0.30 ab | 2.28 ± 0.11 b | 2.69 ± 0.22 a | 0.049 |
| C18:0 | 6.31 ± 0.44 | 7.43 ± 0.11 | 6.79 ± 0.41 | 7.11 ± 0.67 | 0.168 |
| C23:0 | 1.41 ± 0.07 a | 1.32 ± 0.27 ab | 0.96 ± 0.15 b | 1.56 ± 0.29 a | 0.046 |
| Total SFA | 33.26 ± 2.08 | 36.01 ± 1.92 | 35.45 ± 2.20 | 36.78 ± 2.56 | 0.308 |
| C15:1 | 0.34 ± 0.14 ab | 0.41 ± 0.05 a | 0.49 ± 0.13 a | 0.17 ± 0.09 b | 0.030 |
| C16:1n-7 | 7.28 ± 0.42 | 7.17 ± 0.37 | 7.46 ± 0.32 | 7.24 ± 0.34 | 0.810 |
| C17:1 | 4.14 ± 0.37 | 3.62 ± 0.31 | 3.82 ± 0.38 | 3.93 ± 0.34 | 0.388 |
| C18:1n-9 | 1.65 ± 0.45 | 1.31 ± 0.08 | 1.39 ± 0.15 | 1.26 ± 0.13 | 0.298 |
| C18:1n-7 | 2.88 ± 0.18 | 2.77 ± 0.16 | 2.87 ± 0.14 | 2.75 ± 0.12 | 0.641 |
| C20:1 | 5.81 ± 0.35 | 6.20 ± 0.43 | 6.17 ± 0.50 | 5.87 ± 0.47 | 0.620 |
| Total MUFA | 22.10 ± 1.58 | 21.48 ± 1.28 | 22.20 ± 1.07 | 21.22 ± 1.69 | 0.802 |
| C18:3n-3 | 1.48 ± 0.15 | 1.78 ± 0.21 | 1.63 ± 0.12 | 1.67 ± 0.09 | 0.182 |
| C18:4n-3 | 3.52 ± 0.26 | 3.84 ± 0.27 | 3.32 ± 0.32 | 3.57 ± 0.21 | 0.205 |
| C20:5n-3 | 13.06 ± 1.02 | 12.50 ± 0.57 | 12.79 ± 0.62 | 11.75 ± 0.87 | 0.280 |
| C22:5n-3 | 0.43 ± 0.14 b | 0.63 ± 0.08 b | 1.14 ± 0.18 a | 1.33 ± 0.25 a | 0.001 |
| C22:6n-3 | 18.15 ± 1.10 | 16.70 ± 0.92 | 16.34 ± 1.12 | 16.18 ± 1.52 | 0.239 |
| Total n-3 PUFA | 36.64 ± 2.51 | 35.45 ± 1.46 | 35.22 ± 1.93 | 34.50 ± 1.85 | 0.628 |
| C18:2n-6 | 2.21 ± 0.27 | 2.24 ± 0.20 | 1.91 ± 0.17 | 1.84 ± 0.37 | 0.225 |
| C20:2n-6 | 0.52 ± 0.14 a | 0.18 ± 0.05 b | 0.18 ± 0.04 b | 0.47 ± 0.23 a | 0.027 |
| C20:3n-6 | 0.45 ± 0.26 ab | 0.14 ± 0.07 b | 0.23 ± 0.09 b | 0.56 ± 0.14 a | 0.040 |
| C20:4n-6 | 2.83 ± 0.31 | 2.60 ± 0.23 | 2.55 ± 0.29 | 2.37 ± 0.21 | 0.274 |
| C22:3n-6 | 1.42 ± 0.58 | 1.17 ± 0.09 | 1.24 ± 0.12 | 1.48 ± 0.34 | 0.667 |
| C22:4n-6 | 0.18 ± 0.11 b | 0.37 ± 0.12 ab | 0.58 ± 0.19 a | 0.61 ± 0.22 a | 0.043 |
| C22:5n-6 | 0.40 ± 0.04 | 0.35 ± 0.07 | 0.38 ± 0.17 | 0.22 ± 0.14 | 0.304 |
| Total n-6 PUFA | 8.01 ± 0.74 | 7.05 ± 0.49 | 7.07 ± 0.54 | 7.55 ± 0.58 | 0.232 |
| Total PUFA | 44.65 ± 2.72 | 42.50 ± 1.86 | 42.29 ± 1.91 | 42.05 ± 2.12 | 0.479 |
3.2. Other Materials and Chemicals
3.3. Experimental Animals
3.4. Induction, Treatment and Measurement of AIA in Rats
3.5. Induction, Treatment, and Measurement of CIA in Rats
3.6. Sample Collection
3.7. Measurement of Inflammatory Mediators and Cytokines in Serum and Ankle Joint Synovial Fluids of Rats
3.8. Measurement of MMPs and TIMPs mRNA Expressions in the Knee Joint Synovium Tissues of Rats
| Gene Name | Primer Sequence (5'–3') | Size |
|---|---|---|
| MMP1 | Forward: GAGAAAGAAGACAAAGGCAA Reverse: AGCCACATCAGGCACTCC | 164 bp |
| MMP3 | Forward: CGGTGGCTTCAGTACCTT Reverse: CCTCCTCCCAGACCTTCA | 140 bp |
| MMP13 | Forward: GCCAGAACTTCCCAACCA Reverse: ACCCTCCATAATGTCATACCC | 115 bp |
| TIMP1 | Forward: CTCTGGCATCCTCTTGTTG Reverse: CGCTGGTATAAGGTGGTCT | 156 bp |
| GAPDH | Forward: GCAAGTTCAACGGCACAG Reverse: GCCAGTAGACTCCACGACAT | 140 bp |
3.9. Measurement of Hepatic Enzyme Activities in the Serum of Rats
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Scott, D.L.; Wolfe, F.; Huizinga, T.W. Rheumatoid arthritis. Lancet 2010, 376, 1094–1108. [Google Scholar] [CrossRef]
- Van Manen, M.D.; Nace, J.; Mont, M.A. Management of primary knee osteoarthritis and indications for total knee arthroplasty for general practitioners. J. Am. Osteopath. Assoc. 2012, 112, 709–715. [Google Scholar]
- Hawkins, C.; Hanks, G.W. The gastroduodenal toxicity of nonsteroidal anti-inflammatory drugs: A review of the literature. J. Pain Symptom Manag. 2000, 20, 140–151. [Google Scholar] [CrossRef]
- Walker-Bone, K. ‘Natural remedies’ in the treatment of osteoarthritis. Drugs Aging 2003, 20, 517–526. [Google Scholar] [CrossRef]
- Macrides, T.A.; Kalafatis, N. Super-Critical Lipid Extract from Mussels Having Anti-Inflammatory Activity. United States Patent 6,083,536, 4 July 2000. [Google Scholar]
- Whitehouse, M.W.; Macrides, T.A.; Kalafatis, N.; Betts, W.H.; Haynes, D.R.; Broadbent, J. Anti-inflammatory activity of a lipid fraction (Lyprinol) from the NZ green-lipped mussel. Inflammopharmacology 1997, 5, 237–246. [Google Scholar] [CrossRef]
- Pollard, B.; Guilford, W.G.; Ankenbauer-Perkins, K.L.; Hedderley, D. Clinical efficacy and tolerance of an extract of green-lipped mussel (Perna canaliculus) in dogs presumptively diagnosed with degenerative joint disease. N. Z. Vet. J. 2006, 54, 114–118. [Google Scholar] [CrossRef]
- Gibson, S.L.M.; Gibson, R.G. The treatment of arthritis with a lipid extract of Perna canaliculus: A randomized trial. Comp. Ther. Med. 1998, 6, 122–126. [Google Scholar] [CrossRef]
- Lau, C.S.; Chiu, P.K.Y.; Chu, E.M.Y.; Cheng, I.Y.W.; Tang, W.M.; Man, R.Y.K.; Halpern, G.M. Treatment of knee osteoarthritis with Lyprinol®, lipid extract of the green-lipped mussel-a double-blind placebo-controlled study. Prog. Nutr. 2004, 6, 17–31. [Google Scholar]
- Cho, S.H.; Jung, Y.B.; Seong, S.C.; Park, H.B.; Byun, K.Y.; Lee, D.C.; Song, E.K.; Son, J.H. Clinical efficacy and safety of Lyprinol, a patented extract from New Zealand green-lipped mussel (Perna Canaliculus) in patients with osteoarthritis of the hip and knee: A multicenter 2-month clinical trial. Eur. Ann. Allergy Clin. Immunol. 2003, 35, 212–216. [Google Scholar]
- Brien, S.; Prescott, P.; Coghlan, B.; Bashir, N.; Lewith, G. Systematic review of the nutritional supplement Perna Canaliculus (green-lipped mussel) in the treatment of osteoarthritis. Q. J. Med. 2008, 101, 167–179. [Google Scholar] [CrossRef]
- Singh, M.; Hodges, L.D.; Wright, P.F.A.; Cheah, D.M.Y.; Wynne, P.M.; Kalafatis, N.; Macrides, T.A. The CO2-SFE crude lipid extract and the free fatty acid extract from Perna canaliculus have anti-inflammatory effects on adjuvant-induced arthritis in rats. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2008, 149, 251–258. [Google Scholar] [CrossRef]
- McPhee, S.; Hodges, L.D.; Wright, P.F.A.; Wynne, P.M.; Kalafatis, N.; Harney, D.W.; Macrides, T.A. Anti-cyclooxygenase effects of lipid extracts from the New Zealand green-lipped mussel, Perna canaliculus. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2007, 146, 346–356. [Google Scholar] [CrossRef]
- Dugas, B. Lyprinol® inhibits LTB4 production by human monocytes. Allerg. Immunol. 2000, 32, 284–289. [Google Scholar]
- Li, G.; Li, J.; Li, D. Seasonal variation in nutrient composition of Mytilus coruscus from China. J. Agric. Food Chem. 2010, 58, 7831–7837. [Google Scholar] [CrossRef]
- Murphy, K.J.; Mooney, B.D.; Mann, N.J.; Nichols, P.D.; Sinclair, A.J. Lipid, FA, and sterol composition of New Zealand green lipped mussel (Perna canaliculus) and Tasmanian blue mussel (Mytilus edulis). Lipids 2002, 37, 587–595. [Google Scholar] [CrossRef]
- Yuan, G.-F.; Yao, T.; Li, J.; Zhang, Z.-G.; Li, D. Anti-inflammatory activity of the lipid extract of Mytilus crassitesta. J. Zhejiang Univ. (Agric. Life Sci.) 2007, 33, 169–173. (in Chinese). [Google Scholar]
- Bendele, A.M.; McComb, J.; Gould, T.; Mcabee, T.; Sennello, G.; Chlipala, E.; Guy, M. Animal models of arthritis: Relevance to human disease. Toxicol. Pathol. 1999, 27, 134–142. [Google Scholar] [CrossRef]
- Bendele, A.M. Animal models of rheumatoid arthritis. J. Musculoskel. Neuron. Interact. 2001, 1, 377–385. [Google Scholar]
- Joe, B.; Griffiths, M.M.; Remmers, E.F.; Wilder, R.L. Animal models of rheumatoid arthritis and related inflammation. Curr. Rheumatol. Rep. 1999, 1, 139–148. [Google Scholar]
- Bevaart, L.; Vervoordeldonk, M.J.; Tak, P.P. Evaluation of therapeutic targets in animal models of arthritis: How does it relate to rheumatoid arthritis? Arthritis Rheum. 2010, 62, 2192–2205. [Google Scholar] [CrossRef]
- Durie, F.H.; Fava, R.A.; Noelle, R.J. Collagen-induced arthritis as a model of rheumatoid arthritis. Clin. Immunol. Immunopathol. 1994, 73, 11–18. [Google Scholar] [CrossRef]
- Bui, L.M.; Bierer, T.L. Influence of green lipped mussels (Perna canaliculus) in alleviating signs of arthritis in dogs. Vet. Ther. 2003, 4, 397–407. [Google Scholar]
- Cobb, C.S.; Ernst, E. Systematic review of a marine nutriceutical supplement in clinical trials for arthritis: The effectiveness of the New Zealand green-lipped mussel Perna canaliculus. Clin. Rheumatol. 2006, 25, 275–284. [Google Scholar] [CrossRef]
- Khanapure, S.P.; Garvey, D.S.; Janero, D.R.; Letts, L.G. Eicosanoids in inflammation: Biosynthesis, pharmacology, and therapeutic frontiers. Curr. Top. Med. Chem. 2007, 7, 311–340. [Google Scholar] [CrossRef]
- Lewis, R.A.; Austen, K.F.; Soberman, R.J. Leukotrienes and other products of the 5-lipoxygenase pathway—Biochemistry and relation to pathobiology in human diseases. N. Engl. J. Med. 1990, 323, 645–655. [Google Scholar] [CrossRef]
- Tilley, S.L.; Coffman, T.M.; Koller, B.H. Mixed messages: Modulation of inflammation and immune responses by prostaglandins and thromboxanes. J. Clin. Invest. 2001, 108, 15–23. [Google Scholar]
- James, M.J.; Gibson, R.A.; Cleland, L.G. Dietary polyunsaturated fatty acids and inflammatory mediator production. Am. J. Clin. Nutr. 2000, 71 (Suppl.), 343–348. [Google Scholar]
- Hawkes, J.S.; James, M.J.; Cleland, L.G. Biological activity of prostaglandin E3 with regard to oedema formation in mice. Agents Actions 1992, 35, 85–87. [Google Scholar] [CrossRef]
- Goldman, D.W.; Pickett, W.C.; Goetzl, E.J. Human neutrophil chemotactic and degranulating activities of leukotriene B5 (LTB5) derived from eicosapentaenoic acid. Biochem. Biophys. Res. Commun. 1983, 117, 282–288. [Google Scholar] [CrossRef]
- Mclnnes, I.B.; Schett, G. Cytokines in the pathogenesis of rheumatoid arthritis. Nat. Rev. Immunol. 2007, 7, 429–442. [Google Scholar] [CrossRef]
- Brennan, F.M.; Mclnnes, I.B. Evidence that cytokines play a role in rheumatoid arthritis. J. Clin. Invest. 2008, 118, 3537–3545. [Google Scholar] [CrossRef]
- Woods, J.M.; Katschke, K.J.; Volin, M.V.; Ruth, J.H.; Wooddruff, D.C.; Amin, M.A.; Connors, M.A.; Kurata, H.; Arai, K.; Haines, G.K.; et al. IL-4 adenoviral gene therapy reduces inflammation, proinflammatory cytokines, vascularization, and bony destruction in rat adjuvant-induced arthritis. J. Immunol. 2001, 166, 1214–1222. [Google Scholar]
- Brennan, F.M. IL-10 and arthritis. Rheumtology 1999, 38, 293–297. [Google Scholar] [CrossRef]
- Jackson, C.; Nguyen, M.; Arkell, J.; Sambrook, P. Selective matrix metalloproteinase (MMP) inhibition in rheumatoid arthritis-targetting gelatinase A activation. Inflamm. Res. 2001, 50, 183–186. [Google Scholar] [CrossRef]
- Burrage, P.S.; Mix, K.S.; Brinckerhoff, C.E. Matrix metalloproteinases: Role in arthritis. Front. Biosci. 2006, 11, 529–543. [Google Scholar] [CrossRef]
- Yoshihara, Y.; Nakamura, H.; Obata, K.; Yamada, H.; Hayakawa, T.; Fujikawa, K.; Okada, Y. Matrix metalloproteinases and tissue inhibitors of metalloproteinases in synovial fluids from patients with rheumatoid arthritis or osteoarthritis. Ann. Rheum. Dis. 2000, 59, 455–461. [Google Scholar] [CrossRef]
- Brew, K.; Dinakarpandian, D.; Nagase, H. Tissue inhibitors of metalloproteinases: Evolution, structure and function. Biochim. Biophys. Acta 2000, 1477, 267–283. [Google Scholar]
- Visse, R.; Nagase, H. Matrix metalloproteinases and tissue inhibitors of metalloproteinases: Structure, function, and biochemistry. Circ. Res. 2003, 92, 827–839. [Google Scholar] [CrossRef]
- Fries, J.R.; Gurkirpal, S.; Leneert, L.; Furst, D.E. Aspirin, hydroxichloroquine and hepatic enzyme abnormalities with methotrexate in rheumatoid arthritis. Arthritis Rheum. 1990, 33, 1611–1619. [Google Scholar] [CrossRef]
- Suzuki, Y.; Uehara, R.; Tagima, C.; Noguchi, A.; Ide, M.; Ichikawa, Y.; Mizushima, Y. Elevation of serum hepatic aminotransferases during treatment of rheumatoid arthritis with low-dose methotrexate: Risk factors and response to folic acid. Scand. J. Rheumatol. 1999, 28, 273–281. [Google Scholar] [CrossRef]
- Anderson, A.J. Lysosomal enzyme activity in rats with adjuvant induced arthritis. Ann. Rheum. Dis. 1976, 29, 307–313. [Google Scholar] [CrossRef]
- Silva, M.A.; Ishii-Iwamoto, E.L.; Bracht, A.; Caparroz-Assef, S.M.; Kimura, E.; Cuman, R.K.N.; Bersani-Amado, C.A. Efficiency of combined methotrexate/chloroquine therapy in adjuvant-induced arthritis. Fund. Clin. Pharmacol. 2005, 19, 479–489. [Google Scholar] [CrossRef]
- Wang, T.Y.; Li, J.; Ge, J.F.; Li, C.Y.; Jin, Y.; Lü, X.W.; Cheng, W.M.; Tang, J.H. Preliminary study of total flavonoids from Litsea coreana levl. on experimental adjuvant-induced arthritis in rats. Am. J. Chin. Med. 2008, 36, 899–912. [Google Scholar] [CrossRef]
- Yang, Y.; Hutchinson, P.; Morand, E.F. Inhibitory effect of annexin I on synovial inflammation in rat adjuvant arthritis. Arthritis Rheum. 1999, 42, 1538–1544. [Google Scholar] [CrossRef]
- Pine, P.R.; Chang, B.; Schoettler, N.; Banquerigo, M.L.; Wang, S.; Lau, A.; Zhao, F.; Grossbard, E.B.; Payan, D.G.; Brahn, E. Inflammation and bone erosion are suppressed in models of rheumatoid arthritis following treatment with a novel Syk inhibitor. Clin. Immunol. 2007, 124, 244–257. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, G.; Fu, Y.; Zheng, J.; Li, D. Anti-Inflammatory Activity and Mechanism of a Lipid Extract from Hard-Shelled Mussel (Mytilus Coruscus) on Chronic Arthritis in Rats. Mar. Drugs 2014, 12, 568-588. https://doi.org/10.3390/md12020568
Li G, Fu Y, Zheng J, Li D. Anti-Inflammatory Activity and Mechanism of a Lipid Extract from Hard-Shelled Mussel (Mytilus Coruscus) on Chronic Arthritis in Rats. Marine Drugs. 2014; 12(2):568-588. https://doi.org/10.3390/md12020568
Chicago/Turabian StyleLi, Guipu, Yuanqing Fu, Jusheng Zheng, and Duo Li. 2014. "Anti-Inflammatory Activity and Mechanism of a Lipid Extract from Hard-Shelled Mussel (Mytilus Coruscus) on Chronic Arthritis in Rats" Marine Drugs 12, no. 2: 568-588. https://doi.org/10.3390/md12020568
APA StyleLi, G., Fu, Y., Zheng, J., & Li, D. (2014). Anti-Inflammatory Activity and Mechanism of a Lipid Extract from Hard-Shelled Mussel (Mytilus Coruscus) on Chronic Arthritis in Rats. Marine Drugs, 12(2), 568-588. https://doi.org/10.3390/md12020568