
Magnetotactic Bacteria as Potential Sources of Bioproducts

Abstract
:1. Introduction
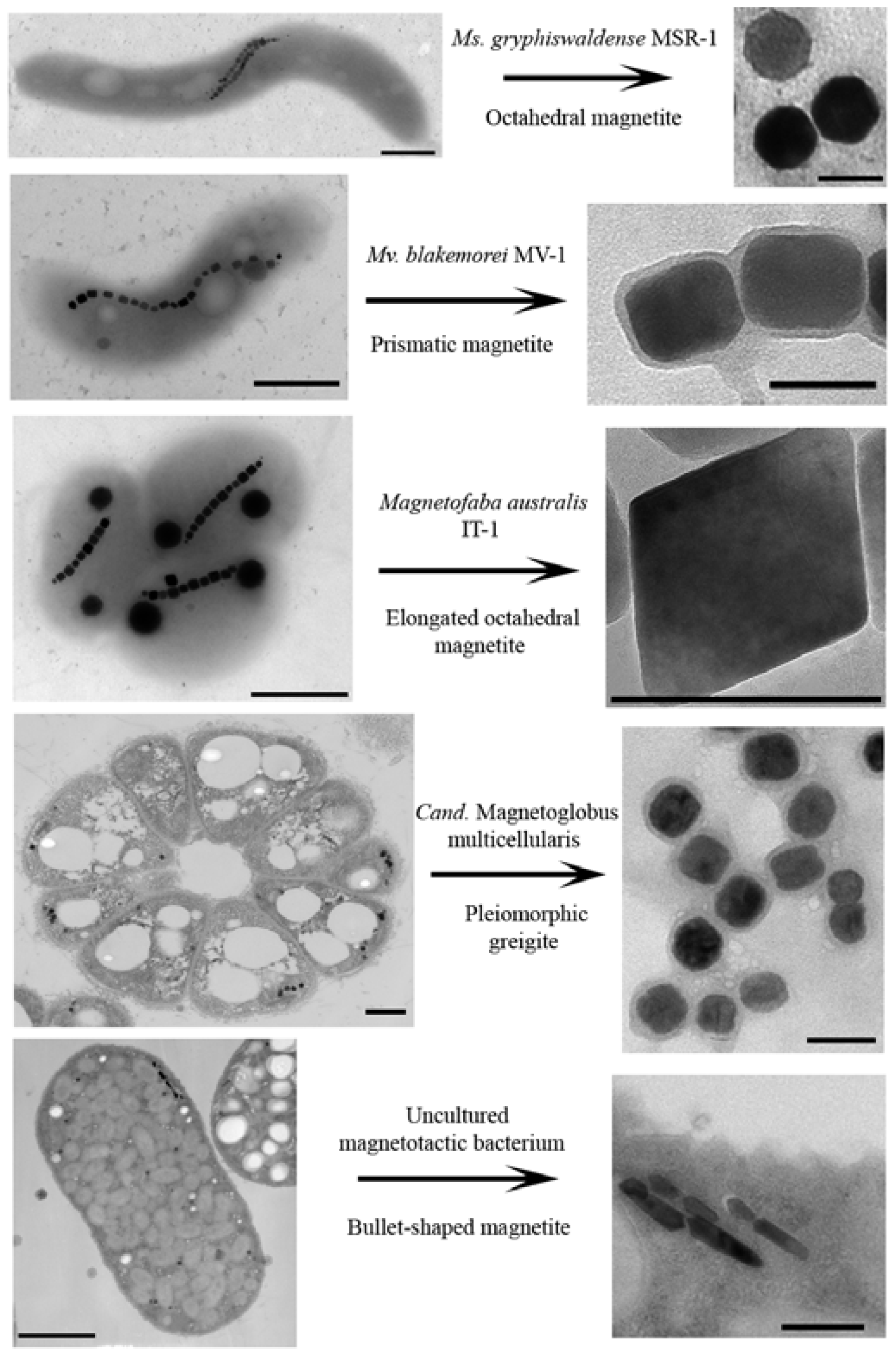
2. Biology of MTB and Their Magnetosomes
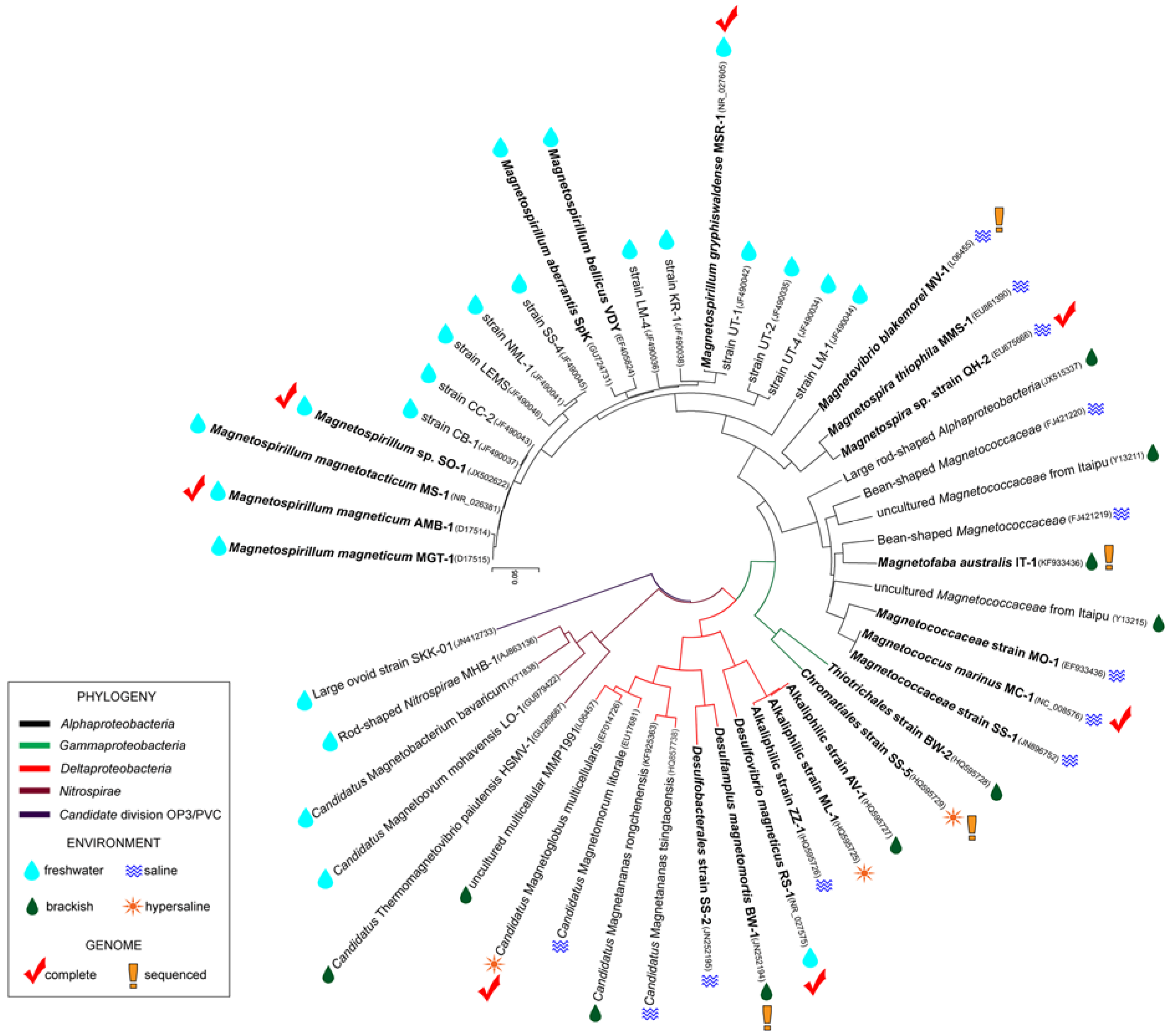
2.1. Ecology and Physiology of MTB


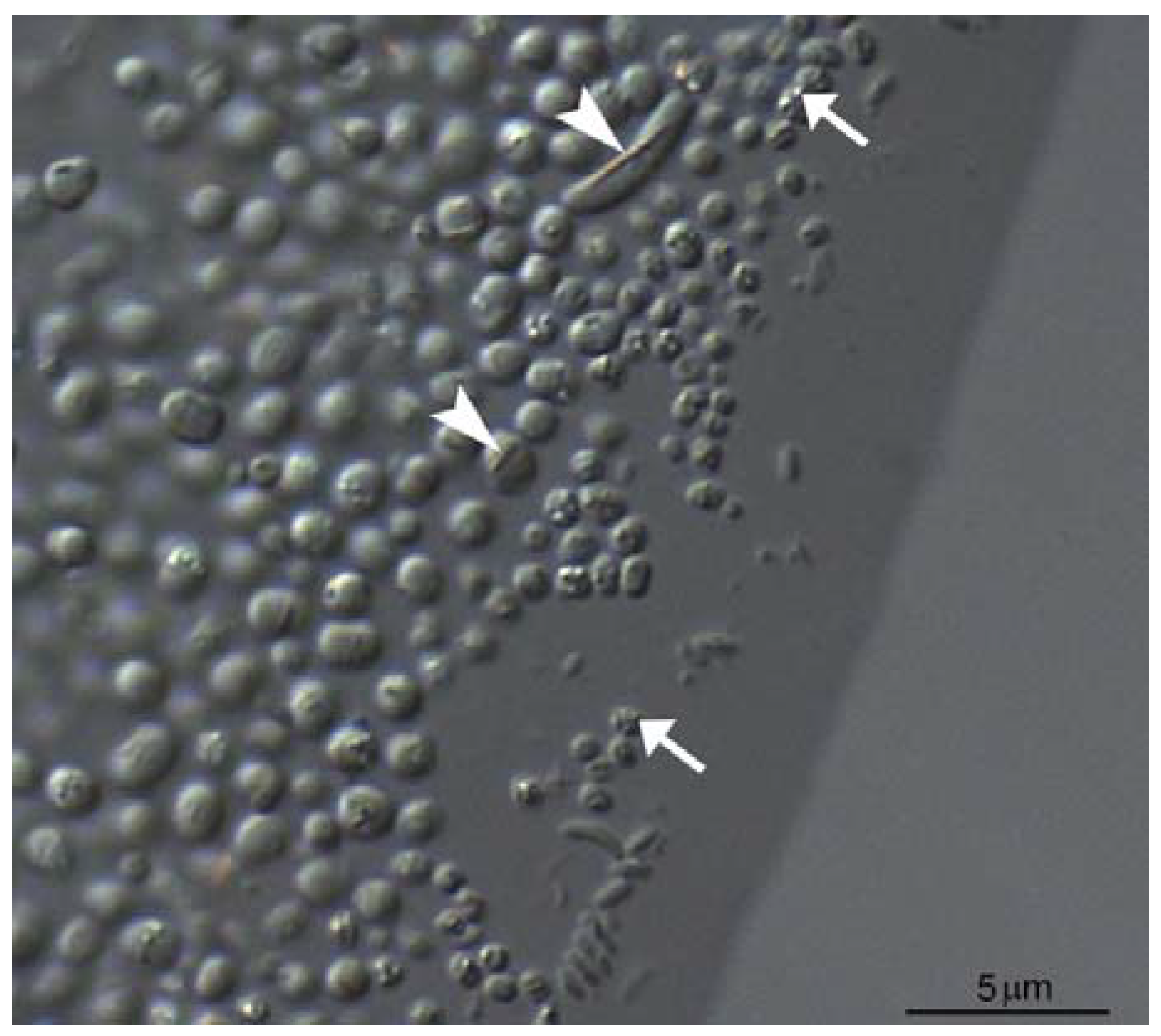
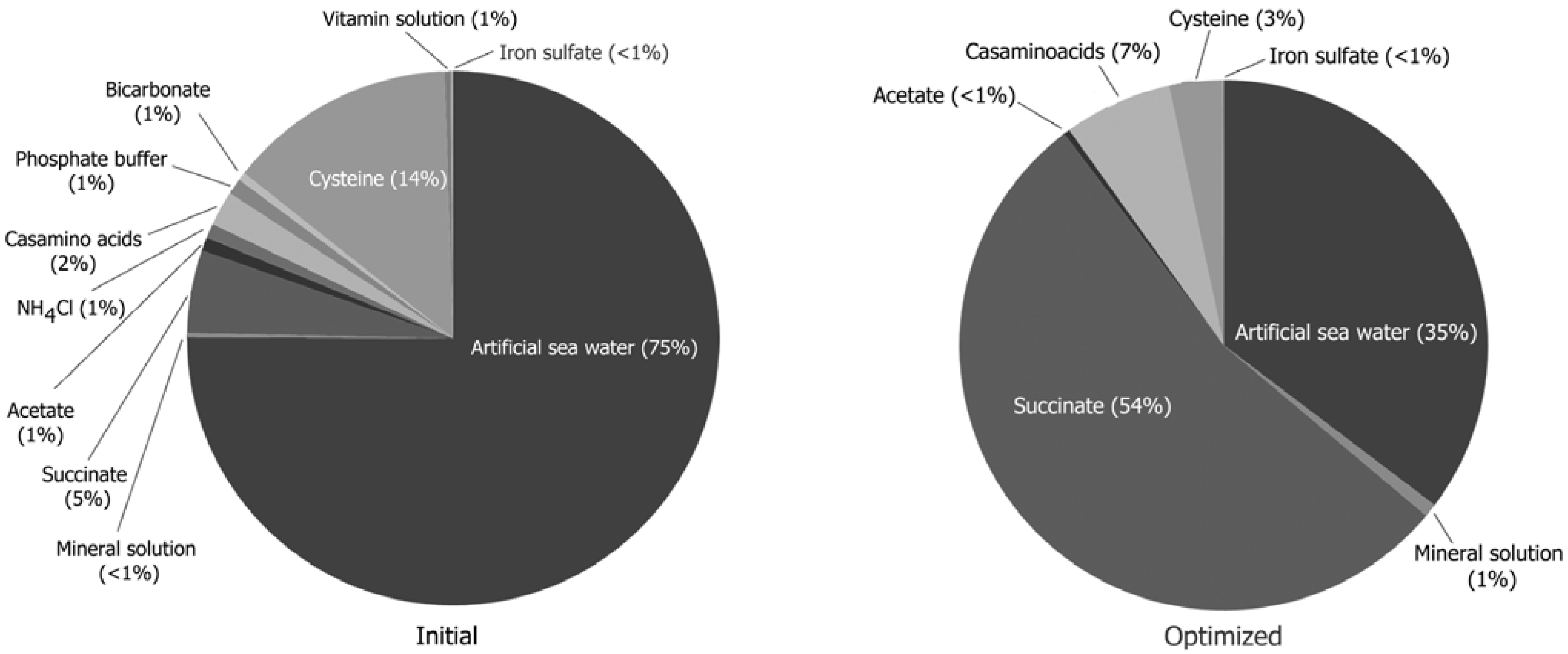
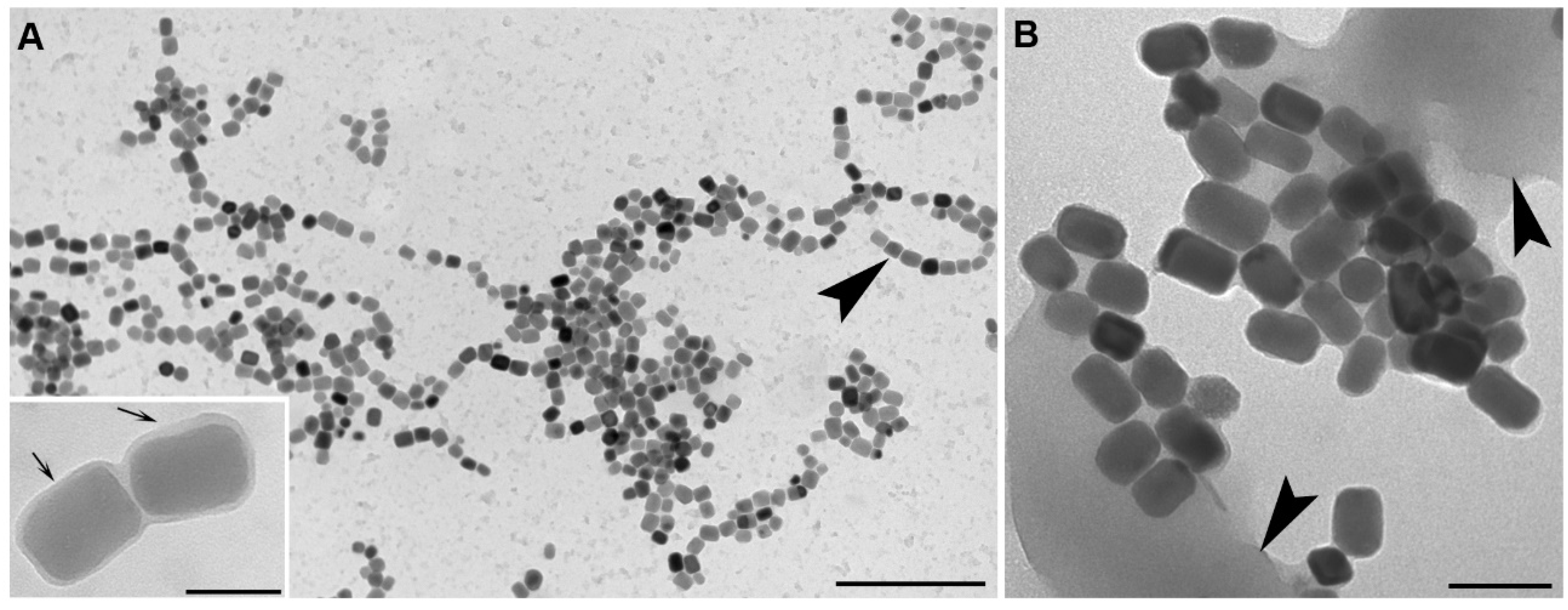
2.2. Isolation and Cultivation of MTB

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Bacterial Morphology | Magnetosome | References | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Shape | Size (Length × Width) µm | Crystal Shape and Composition | Size (Length × Width) nm | Number/Cell | Size (Length × Width) nm | Number/Cell | Magneto-Some Production | ||
| Autotrophic | Heterotrophic | ||||||||
| Magnetospirillum magneticum AMB-1 | Spirillum | 3 × 0.4–0.6 | Cuboctahedral magnetite | No growth | No growth | 41 ± 15 | 12 ± 5 (anaerobic) | 1.4 × 109 cells mL−1; 2.6 mg L−1 magnetite (=2.8% cell weight) | [12,29] |
| SF = 0.78 | |||||||||
| (anaerobic) | |||||||||
| 33 ± 8.5 | |||||||||
| SF = 0.89 | 7 ± 4 (aerobic) | ||||||||
| (aerobic) | |||||||||
| Magnetospirillum magnetotacticum MS-1 | Spirillum | 4–6 × 0.25 | Cuboctahedral magnetite | No growth | No growth | 42 (25–55) | 17.6 (5–41) | 0.2–0.6 g cell L−1 (wet weight) | [13,65] |
| SF = 0.9 | |||||||||
| Magnetospirillum Gryphiswaldense MSR-1 | Spirillum | 1–20 × 0.7 | Cuboctahedral magnetite | NI | NI | Ø 46 ± 6.8 (14–67) | 23.4 ± 0.9 | 41.7 mg L−1 (16.7 mg L−1 day−1) | [66,67] |
| SF = 0.91 | |||||||||
| Magnetovibrio blakemorei MV-1 | Vibrio | 1–3 × 0.2–0.4 | Elongated prismatic magnetite | 48 ± 5 (30–59) × 26 ± 7 (28–40) | 17 ± 4 (7–23) | 60 × 40 | 15.34 ± 4 | 15.14 mg L−1; (4.98 mg L−1 day−1) | [39,68,69] |
| AR = 1.8 ± 0.3 | SF = 0.65 | ||||||||
| Magnetospira thiophila MMS-1 | Spirillum | 1–3 × 0.2–0.5 | Elongated octahedral magnetite | NI | NI | 61 ± 12 (22–85) × 52 ± 11 (18–80) | 17 ± 5 (8–31) | NI | [18,69] |
| AR = 1.2 ± 0.1 | |||||||||
| SF = 0.85 | |||||||||
| Magnetospira thiophila QH-2 | Spirillum | 2.0 ± 0.4 (1–3) × 0.8 ± 0.2 | Elongated octahedral magnetite | NI | NI | 81 ± 23 × 58 ± 20 SF = 0.71 ± 0.11 | 16 ± 5 (7–28) | NI | [19] |
| Magnetofaba australis IT-1 | Faba-bean | 1.4 ± 0.3 × 1.1 ± 0.3 (n = 130) | Elongated octahedral magnetite | NI | 6 ± 4 (n = 100) | 83 ± 26 × 74 ± 23 | 10 ± 3 (n = 100) | NI | [16] |
| SF = 0.89 ± 0.05 | |||||||||
| Magnetococcus marinus MC-1 | Cocci | Ø = 1–2 µm | Elongated pseudo-hexagonal prismatic magnetite | 72 ± 11 (33–95) × 70 ± 13 (29–87) | 10 ± 2 (6–15) | 83 ± 14 (30–110) × 78 ± 11 (15–107) | 14 ± 3 (8–19) | NI | [15,69] |
| AR=1.2 ± 0.2 | AR = 1.2 ± 0.1 | ||||||||
| SF = 0.93 | |||||||||
| Magnetococcus MO-1 | Ovoid | 1.33 ± 0.19 × 1.85 ± 0.40 | Elongated cuboctahedral magnetite | No growth | No growth | 64 ± 20 × 57 ± 17 | 17 ± 5 | NI | [14] |
| SF = 0.89 | |||||||||
| Strain BW-2 | Rod | 4.4 ± 0.6 × 2.2 ± 0.2 (n = 62) | Cuboctahedral magnetite | 67 ± 16 × 63 ± 15 | 30 ± 9 (n = 46) | No growth | No growth | NI | [25] |
| SF= 0.94 ± 0.04 (n = 189) | |||||||||
| Strain SS-5 | Rod | 2.5 ± 0.5 × 1.2 ± 0.1 (n = 64) | Elongated prismatic magnetite | 86 ± 27 × 63 ± 19 | 20 ± 7 (n = 45) | NI | NI | NI | [25] |
| SF = 0.74 ± 0.07 (n = 171) | |||||||||
| Desulfovibrio magneticus RS-1 | Vibrio | 3–5 × 1 | Bullet-shaped magnetite | No growth | No growth | Mean length = 60 nm (32–85 nm) | 12–15 | NI | [23,70] |
| SF = 0.5 | |||||||||
| Candidatus Desulfamplus magnetomortis BW-1 | Rod | ≈ 4 × ≈1 | Bullet-shaped magnetite and/or pleomorphic greigite | No growth | No growth | Mean length = 55 nm | NI | NI | [24,71] |
| SF = 0.6 | |||||||||
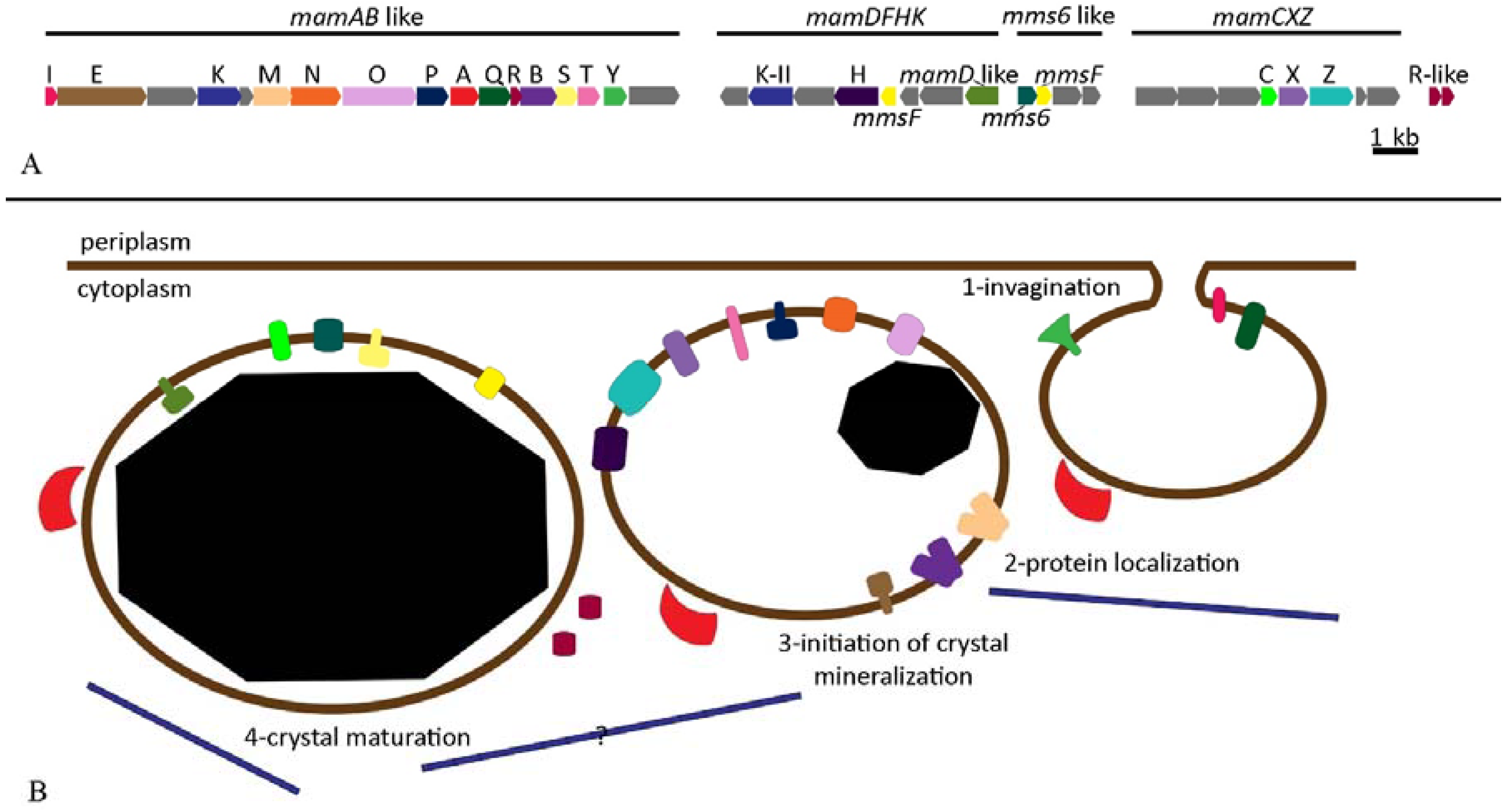
2.3. Magnetosomes


| Protein | Localization | Process | Function | Deletion Effects | References |
|---|---|---|---|---|---|
| MamA | Cytosol. Dynamic, surrounding vesicles | Invagination of cell membrane | It has multiple domains with TPR motifs (protein-protein interactions); may act as multi-protein assembly site; stabilizes magnetosome chain. | Invagination is not affected. Reduction in the number of magnetosomes and changes in iron accumulation. | [89,90,91] |
| MamB | Transmembrane in MM | Iron transport and magnetite nucleation | May be involved in iron transport since has homology to CDF (cation diffusion facilitator). Contains TPR domain (protein-protein interactions) and interacts with MamE; requires MamM for stabilization. | Loss of magnetosome vesicles and of crystal formation. | [74,79,89] |
| MamC | Transmembrane in MM | Crystal shape and size | Its loop interacts with magnetosome crystal. It is not essential to biomineralization but may control chemical conditions inside vesicles. | Changes in size and organization of chains and size of vesicles. No effects observed in crystal size or shape. | [78,89] |
| MamD | Transmembrane in MM, N-terminal in ML | Crystal shape and size | Associated with control over size of magnetosome crystal. | Changes in crystal size. | [78,89] |
| MamE | Transmembrane in MM, C-terminal towards ML | Iron transport and nucleation | Acts as a serine protease and has PDZ domain (protein-protein interaction) which interacts with MamB and I. Magnetochrome might control the magnetosome redox state and balance between Fe2+/Fe3+. | Formation of empty magnetosome vesicles, loss of magnetite synthesis, mislocation of MamI and other Mam proteins. | [74,80,89] |
| MamF | Transmembrane MM | Crystal shape and size | Associated to control of magnetosome size; interacts with crystal. | Changes in crystal size. | [78,89] |
| MamG | Transmembrane in MM | Crystal shape and size | Associated to control of magnetosome size. | Changes in crystal size. | [78,89] |
| MamH | Transmembrane in MM | Iron transport and nucleation | Contains conserved domains homologous to MFS proteins (membrane transporters) and might function as phosphate transporter during magnetite biomineralization. | Reduced number and size of magnetosomes. | [74,89,92,93] |
| MamI | Transmembrane in MM | Invagination | Involved in the formation and bending of the MM. | Absence of MM. | [28,74,89] |
| MamJ | Cytosol | Arrangement of chains | Acts as an anchor between MamK filaments and vesicle membrane to arrange magnetosomes in a chain. | Magnetosomes arranged in clusters and no longer in chains. Reduced magnetotactic response. | [78] |
| MamK | Cytosol | Arrangement of chains | Controls chain assembly and position along the cell axis; positions chain for cellular division; homologous to MreB (actin-like). | Lack of filaments near the magnetosomes. Shorter chains and wrong position of MamJ. | [94] |
| MamL | Transmembrane in MM | Invagination | Involved in the formation of MM; similar to MamI. | Absence of MM. | [74] |
| MamM | Transmembrane in MM | Iron transport and magnetite nucleation | Involved in iron transport and may use H+/cation antiporter mechanism. Involved in the begining of crystalization and localization of other Mam proteins; stabilizes MamB; homologous to CDF (cation diffusion facilitator). | Loss of magnetite crystals, formation of empty vesicles. | [74,79,89] |
| MamN | Transmembrane in MM | Iron transport and magnetite nucleation | Homologous to Na+/H+ antiporter and might be involved in the extrusion of H+ from the vesicle. | Formation of empty magnetosome vesicles. Does not affect localization of other proteins. | [28,74,89] |
| MamO | Transmembrane in MM; C-terminal in ML | Iron transport and magnetite nucleation | Composed of two domains: (1) transmembrane, homologous to proteins involved in transport of anions across cell membrane and (2) similar to a trypsin-like peptidase, but possibly with no protease function. | Formation of empty magnetosome vesicles. | [74,89] |
| MamP | Transmembrane in MM with active sites towards ML | Iron transport and magnetite nucleation | Involved in control of crystal number and size and in electron transfer necessary to magnetosome assembly and magnetite formation; similar to MamE and MamT; may contain an iron-binding site. | Defects in crystal size, fewer magnetosomes per cell. | [74,89,95] |
| MamQ | Transmembrane in MM; C-terminal in ML | Invagination | Unknown function; homologous to LemA. | Complete loss of magnetosome formation in AMB-1. | [74,89] |
| MamR | Cytosol | Crystal shape and size | Controls the number and size of crystals; predicted to have a DNA-binding domain. | Smaller magnetosome and weaker magnetotactic response. | [74,80,89] |
| MamS | Transmembrane in MM; C-terminal in ML | Crystal shape and size | Controls the number and size of crystals. | Defects in crystal size and morphology, weaker magnetotactic response. | [74] |
| MamT | Transmembrane in MM; C-terminal in ML | Iron transport and magnetite nucleation | Involved in regulation of crystal size and morphology; has a magnetochrome domain. | Defects in crystal maturation and loss of magnetotactic response. | [74,89,95] |
| MamU | Cytosol | Invagination | Unknown function. Homologous to DGK Family, that includes kinase involved in regulation of cell response. | None observed. | [74,89] |
| MamV | Transmembrane in MM | Iron transport and magnetite nucleation | Putative CDF transporter. | None observed. | [74,79] |
| MamW | MM (structure unknown) | Iron transport and magnetite nucleation | Implicated it magnetite synthesis or not associated to magnetosomes. | None observed. | [76,77] |
| MamX | Transmembrane in MM; C-terminal in ML | Iron transport and magnetite nucleation | Involved in electron transport, with Cytochrome c-like domain; weak similarity to MamS and E. | Smaller crystals and with irregular shapes. Weaker magnetotactic cell response. | [89,92] |
| MamY | Transmembrane in MM; C-terminal in cytosol | Invagination | Constricts the MM and consequently affects crystal growth; homologous to BAR proteins (involved in membrane dynamics). | Enlarged magnetosome vesicles with smaller crystals. | [96] |
| MamZ | Transmembrane in MM; C-terminal in ML | Iron transport and magnetite nucleation | Involved in redox control for magnetosome formation; creates an iron oxidoreductase and transport complex with MamX and MamH. | Smaller size of crystals and higher proportion of twinned crystals. | [89,92] |
| Mms6 | Transmembrane in MM | Crystal shape and size | Involved in the initiation of magnetite synthesis and control of crystal shape; presents in vitro activity. | Smaller magnetosomes with heterogeneous shapes. Irregular alignment of chains. | [89,97] |
| MmsF | Transmembrane in MM | Crystal shape and size | Involved in the control of size and shape of magnetite crystal during maturation. | Formation of elongated crystals and of non-magnetotactic cells. | [74,89] |
2.4. Mass Production of MTB and Magnetosomes
| MTB | Culture | Medium | Magnetite production (mg L−1) * | Magnetite productivity (mg L−1 day−1) * | References |
|---|---|---|---|---|---|
| Ms. magneticum | Fed-Batch | MSGM | 9 ± 0.7 | 3.7 ± 0.13 | [104] |
| Ms. gryphiswaldense | Batch | LSM | 7.9 | 6.3 | [38] |
| Ms. gryphiswaldense NPHB | Fed-Batch | OFM | 58.4 ± 6.4 | - | [107] |
| Ms. gryphiswaldense | Fed-Batch | OFM | 41.7 | 16.7 | [105] |
| Ms. gryphiswaldense | Fed-Batch | OFM | 83.23 ± 5.36 | 55.49 | [108] |
| Ms. gryphiswaldense | Fed-Batch | OFM | 356.52 | 178.26 | [109] |
| Ms. gryphiswaldense | Semi-continuous | OFM | 168.3 | 83.5 | [109] |
| Mv. blakemorei | Batch-flask | [17] | 15.14 | 4.98 | [39] |
| Mv. blakemorei | Batch-flask | Optimized | 64.35 | 16.09 | [39] |
| Mv. blakemorei | Batch | Optimized | 22.4 | 5.6 | [39] |
| Mv. blakemorei | Fed-Batch | Optimized | 26 | 3.2 | [39] |


2.5. Biotechnological Applications of Magnetosomes
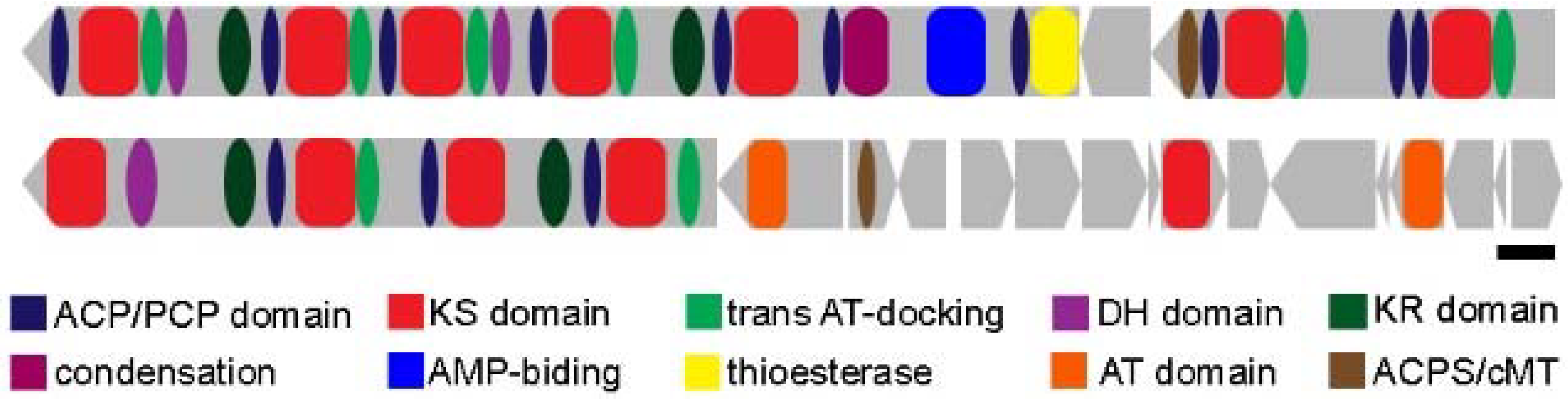
3. Genomic Studies on MTB as Potential Sources for New (Natural) Products
| Species | Strain | Class † | Source | Salinity | Genome (Mb)/MAI (Kb) | PKS | NRPS | Hybrid |
|---|---|---|---|---|---|---|---|---|
| Mc. marinus | MC-1 | α | Pettaquamscutt Estuary—USA | Brackish to marine | 4.71/55.82 | 0 | 0 | 0 |
| Mf. Itaipuensis * | IT-1 | α | Itaipu lagoon—Brazil | Brackish to marine | 4.98/64.9 | 3 | 0 | 0 |
| Mv. blakemorei | MV-1 | α | Saltmarsh pool—USA | Brackish | 3.70/66.03 | 6 | 0 | 1 |
| Magnetospira sp. | QH-2 | α | Intertidal seawater—China | Saline | 4.0/45 | 0 | 2 | 0 |
| Ms. gryphiswaldense | MSR-1 | α | Eutrophic river—Germany | Freshwater | 4.36 + 0.036/74.6 | 3 | 0 | 2 |
| Magnetospirillum sp. * | SO-1 | α | River—Russia | Freshwater | 4.87/100 | 0 | 0 | 1 |
| Ms. magneticum | AMB-1 | α | Koganei ponds—Japan | Freshwater | 4.97/73 | 0 | 0 | 1 |
| Order Chromatiales | SS-5 | γ | Salton Sea—USA | Hypersaline | 3.7/ND | 1 | 3 | 5 |
| Ca. Da. Magnetomortis * | BW-1 | δ | Badwater Basin—USA | Brackish | 6.8/ND | 8 | 4 | 3 |
| Ca. Mg. multicellularis * | MMP | δ | Araruama Lagoon—Brazil | Hypersaline | 12.8/15.7 | 9 | 11 | 4 |
| Desulfovibrio magneticus | RS-1 | δ | Kameno River—Japan | Freshwater | 5.25 + 0.058 + 0.008/71 | 0 | 0 | 0 |

4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bazylinski, D.A.; Frankel, R.B. Magnetosome formation in prokaryotes. Nat. Rev. Microbiol. 2004, 2, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Bazylinski, D.A.; Frankel, R.B.; Heywood, B.R.; Mann, S.; King, J.W.; Donaghay, L.; Hanson, A.K. Controlled biomineralization of magnetite (Fe3O4) and greigite (Fe3S4) in a magnetotactic bacterium. Appl. Environ. Microbiol. 1995, 61, 3232–3239. [Google Scholar] [PubMed]
- Lins, U.; Keim, C.N.; Evans, F.F.; Farina, M.; Buseck, P.R. Magnetite (Fe3O4) and Greigite (Fe3S4) Crystals in multicellular magnetotactic prokaryotes. Geomicrobiol. J. 2007, 24, 43–50. [Google Scholar] [CrossRef]
- Moskowitz, B.M.; Bazylinski, D.A.; Egli, R.; Frankel, R.B.; Edwards, K.J. Magnetic properties of marine magnetotactic bacteria in a seasonally stratified coastal pond (Salt Pond, MA, USA). Geophys. J. Int. 2008, 174, 75–92. [Google Scholar] [CrossRef]
- Jogler, C.; Schüler, D. Genomics, genetics, and cell biology of magnetosome formation. Annu. Rev. Microbiol. 2009, 63, 501–521. [Google Scholar] [CrossRef] [PubMed]
- Lefèvre, C.T.; Bazylinski, D.A. Ecology, diversity, and evolution of magnetotactic bacteria. Microbiol. Mol. Biol. Rev. 2013, 77, 497–526. [Google Scholar] [CrossRef] [PubMed]
- Simmons, S.L.; Sievert, S.M.; Frankel, R.B.; Bazylinski, D.A.; Edwards, K.J. Spatiotemporal Distribution of Marine Magnetotactic Bacteria in a Seasonally Stratified Coastal Salt Pond. Appl. Environ. Microbiol. 2004, 70, 6230–6239. [Google Scholar] [CrossRef] [PubMed]
- Vali, H.; Förster, O.; Amarantidis, G.; Petersen, N. Magnetotactic bacteria and their magnetofossils in sediments. Earth Planet. Sci. Lett. 1987, 86, 389–400. [Google Scholar] [CrossRef]
- Lefèvre, C.T.; Abreu, F.; Schmidt, M.M.L.; Lins, U.; Frankel, R.B.; Hedlund, B.P.; Bazylinski, D.A. Moderately thermophilic magnetotactic bacteria from hot springs in Nevada. Appl. Environ. Microbiol. 2010, 76, 3740–3743. [Google Scholar] [CrossRef] [PubMed]
- Kolinko, S.; Jogler, C.; Katzmann, E.; Wanner, G.; Peplies, J.; Schüler, D. Single-cell analysis reveals a novel uncultivated magnetotactic bacterium within the candidate division OP3. Environ. Microbiol. 2012, 14, 1709–1721. [Google Scholar] [CrossRef] [PubMed]
- Schleifer, K.H.; Schüler, D.; Spring, S.; Weizenegger, M.R.; Amann, L.W.; Köhler, M. The genus Magnetospirillum gen. nov. Description of Magnetospirillum gryphiswaldense sp. nov. and transfer of Aquaspirillum magnetotacticum to Magnetospirillum magnetotacticum comb. nov. Syst. Appl. Microbiol. 1991, 14, 379–385. [Google Scholar] [CrossRef]
- Matsunaga, T.; Sakaguchi, T.; Tadokoro, F. Magnetite formation by a magnetic bacterium capable of growing aerobically. Appl. Microbiol. Biotechnol. 1991, 35, 651–655. [Google Scholar]
- Blakemore, R.P.; Maratea, D.; Wolfe, R.S. Isolation and pure culture of a freshwater magnetic spirillum in chemically defined medium. J. Bacteriol. 1979, 140, 720–729. [Google Scholar] [PubMed]
- Lefèvre, C.T.; Bernadac, A.; Yu-Zhang, K.; Pradel, N.; Wu, L.-F. Isolation and characterization of a magnetotactic bacterial culture from the Mediterranean Sea. Environ. Microbiol. 2009, 11, 1646–1657. [Google Scholar] [CrossRef] [PubMed]
- Bazylinski, D.A.; Williams, T.J.; Lefèvre, C.T.; Berg, R.J.; Zhang, C.L.; Bowser, S.S.; Dean, A.J.; Beveridge, T.J. Magnetococcus marinus gen. nov., sp. nov., a marine, magnetotactic bacterium that represents a novel lineage (Magnetococcaceae fam. nov., Magnetococcales ord. nov.) at the base of the Alphaproteobacteria. Int. J. Syst. Evol. Microbiol. 2013, 63, 801–808. [Google Scholar] [CrossRef] [PubMed]
- Morillo, V.; Abreu, F.; Araujo, A.C.; de Almeida, L.G.P.; Enrich-Prast, A.; Farina, M.; de Vasconcelos, A.T.R.; Bazylinski, D.A.; Lins, U. Isolation, cultivation and genomic analysis of magnetosome biomineralization genes of a new genus of South-seeking magnetotactic cocci within the Alphaproteobacteria. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Bazylinski, D.A.; Williams, T.J.; Lefèvre, C.T.; Trubitsyn, D.; Fang, J.; Beveridge, T.J.; Moskowitz, B.M.; Ward, B.; Schübbe, S.; Dubbels, B.L.; et al. Magnetovibrio blakemorei gen. nov. nov., a magnetotactic bacterium (Alphaproteobacteria: Rhodospirillaceae) isolated from a salt marsh. Int. J. Syst. Evol. Microbiol. 2013, 63, 1824–1833. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.J.; Lefèvre, C.T.; Zhao, W.; Beveridge, T.J.; Bazylinski, D.A. Magnetospira thiophila gen. nov., sp. nov., a marine magnetotactic bacterium that represents a novel lineage within the Rhodospirillaceae (Alphaproteobacteria). Int. J. Syst. Evol. Microbiol. 2012, 62, 2443–2450. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Pan, H.; Li, J.; Yu-Zhang, K.; Zhang, S.-D.; Zhang, W.-Y.; Zhou, K.; Yue, H.; Pan, Y.; Xiao, T.; et al. Isolation and characterization of a marine magnetotactic spirillum axenic culture QH-2 from an intertidal zone of the China Sea. Res. Microbiol. 2010, 161, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Abreu, F.; Martins, J.L.; Silveira, T.S.; Keim, C.N.; de Barros, H.G.P.L.; Filho, F.J.G.; Lins, U. “Candidatus Magnetoglobus multicellularis”, a multicellular, magnetotactic prokaryote from a hypersaline environment. Int. J. Syst. Evol. Microbiol. 2007, 57, 1318–1322. [Google Scholar] [CrossRef] [PubMed]
- Zhou, K.; Zhang, W.-Y.; Pan, H.-M.; Li, J.-H.; Yue, H.-D.; Xiao, T.; Wu, L.-F. Adaptation of spherical multicellular magnetotactic prokaryotes to the geochemically variable habitat of an intertidal zone. Environ. Microbiol. 2013, 15, 1595–1605. [Google Scholar] [CrossRef] [PubMed]
- Wenter, R.; Wanner, G.; Schüler, D.; Overmann, J. Ultrastructure, tactic behaviour and potential for sulfate reduction of a novel multicellular magnetotactic prokaryote from North Sea sediments. Environ. Microbiol. 2009, 11, 1493–1505. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, T.; Arakaki, A.; Matsunaga, T. Desulfovibrio magneticus sp. nov., a novel sulfate-reducing bacterium that produces intracellular single-domain-sized magnetite particles. Int. J. Syst. Evol. Microbiol. 2002, 52, 215–221. [Google Scholar] [PubMed]
- Lefèvre, C.T.; Menguy, N.; Abreu, F.; Lins, U.; Pósfai, M.; Prozorov, T.; Pignol, D.; Frankel, R.B.; Bazylinski, D.A. A cultured greigite-producing magnetotactic bacterium in a novel group of sulfate-reducing bacteria. Science 2011, 334, 1720–1723. [Google Scholar] [CrossRef] [PubMed]
- Lefèvre, C.T.; Viloria, N.; Schmidt, M.L.; Pósfai, M.; Frankel, R.B.; Bazylinski, D.A. Novel magnetite-producing magnetotactic bacteria belonging to the Gammaproteobacteria. ISME J. 2012, 6, 440–450. [Google Scholar] [CrossRef] [PubMed]
- Lefèvre, C.T.; Trubitsyn, D.; Abreu, F.; Kolinko, S.; de Almeida, L.G.P.; de Vasconcelos, A.T.R.; Lins, U.; Schüler, D.; Ginet, N.; Pignol, D.; et al. Monophyletic origin of magnetotaxis and the first magnetosomes. Environ. Microbiol. 2013, 15, 2267–2274. [Google Scholar] [CrossRef] [PubMed]
- Gorby, Y.A.; Beveridge, T.J.; Blakemore, R.P. Characterization of the bacterial magnetosome membrane. J. Bacteriol. 1988, 170, 834–841. [Google Scholar] [PubMed]
- Komeili, A. Molecular mechanisms of compartmentalization and biomineralization in magnetotactic bacteria. FEMS Microbiol. Rev. 2012, 36, 232–255. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Pan, Y. Environmental Factors Affect Magnetite Magnetosome Synthesis in Magnetospirillum magneticum AMB-1: Implications for Biologically Controlled Mineralization. Geomicrobiol. J. 2012, 29, 362–373. [Google Scholar] [CrossRef]
- Faivre, D.; Menguy, N.; Pósfai, M.; Schüler, D. Environmental parameters affect the physical properties of fast-growing magnetosomes. Am. Mineral. 2008, 93, 463–469. [Google Scholar] [CrossRef]
- Blakemore, R.P.; Short, K.A.; Bazylinski, D.A.; Rosenblatt, C.; Frankel, R.B. Microaerobic Conditions Are Required for Magnetite Formation within Aquaspirillum magnetotacticum. Geomicrobiol. J. 1985, 4, 53–71. [Google Scholar] [CrossRef]
- Bazylinski, D.A.; Garratt-Reed, A.J.; Frankel, R.B. Electron Microscopic Studies of Magnetosomes in Magnetotactic Bacteria. Microsc. Res. Tech. 1994, 27, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Faivre, D.; Schüler, D. Magnetotactic Bacteria and Magnetosomes. Chem. Rev. 2008, 108, 4875–4898. [Google Scholar] [CrossRef] [PubMed]
- Plank, C.; Schillinger, U.; Scherer, F.; Bergemann, C.; Rémy, J.S.; Krötz, F.; Anton, M.; Lausier, J.; Rosenecker, J. The Magnetofection Method: Using Magnetic Force to Enhance Gene Delivery. Biol. Chem. 2003, 384, 737–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gleich, B.; Weizenecker, J. Tomographic imaging using the nonlinear response of magnetic particles. Nature 2005, 435, 1214–1217. [Google Scholar] [CrossRef] [PubMed]
- Miltenyi, S.; Müller, W.; Weichel, W.; Radbruch, A. High gradient magnetic cell separation with MACS. Cytometry 1990, 11, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Alphandéry, E.; Guyot, F.; Chebbi, I. Preparation of chains of magnetosomes, isolated from Magnetospirillum magneticum strain AMB-1 magnetotactic bacteria, yielding efficient treatment of tumors using magnetic hyperthermia. Int. J. Pharm. 2012, 434, 444–452. [Google Scholar] [CrossRef] [PubMed]
- Heyen, U.; Schüler, D. Growth and magnetosome formation by microaerophilic Magnetospirillum strains in an oxygen-controlled fermentor. Appl. Microbiol. Biotechnol. 2003, 61, 536–544. [Google Scholar] [CrossRef] [PubMed]
- Silva, K.T.; Leão, P.E.; Abreu, F.; López, J.A.; Gutarra, M.L.; Farina, M.; Bazylinski, D.A.; Freire, D.M.G.; Lins, U. Optimization of magnetosome production and growth by the magnetotactic vibrio Magnetovibrio blakemorei strain MV-1 through a statistics-based experimental design. Appl. Environ. Microbiol. 2013, 79, 2823–2827. [Google Scholar] [CrossRef] [PubMed]
- Kolinko, I.; Lohsse, A.; Borg, S.; Raschdorf, O.; Jogler, C.; Tu, Q.; Pósfai, M.; Tompa, E.; Plitzko, J.M.; Brachmann, A.; et al. Biosynthesis of magnetic nanostructures in a foreign organism by transfer of bacterial magnetosome gene clusters. Nat. Nanotechnol. 2014, 9, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Bazylinski, D.A.; Lefèvre, C.T.; Schüler, D. Magnetotactic Bacteria. In The Prokaryotes—Prokaryotic Physiology and Biochemistry, 4th ed.; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 453–494. [Google Scholar]
- Fassbinder, J.W.E.; Stanjek, H.; Vali, H. Occurrence of magnetic bacteria in soil. Nature 1990, 343, 161–163. [Google Scholar] [CrossRef] [PubMed]
- Petermann, H.; Bleil, U. Detection of live magnetotactic bacteria in South Atlantic deep-sea sediments. Earth Planet. Sci. Lett. 1993, 117, 223–228. [Google Scholar] [CrossRef]
- Bazylinski, D.A.; Lefèvre, C.T. Magnetotactic Bacteria from Extreme Environments. Life 2013, 3, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Martins, J.L.; Silveira, T.S.; Silva, K.T.; Lins, U. Salinity dependence of the distribution of multicellular magnetotactic prokaryotes in a hypersaline lagoon. Int. Microbiol. 2009, 12, 193–201. [Google Scholar] [PubMed]
- Lin, W.; Wang, Y.; Li, B.; Pan, Y. A biogeographic distribution of magnetotactic bacteria influenced by salinity. ISME J. 2012, 6, 475–479. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Wang, Y.; Gorby, Y.; Nealson, K.; Pan, Y. Integrating niche-based process and spatial process in biogeography of magnetotactic bacteria. Sci. Rep. 2013, 3. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Pan, Y. Temporal variation of magnetotactic bacterial communities in two freshwater sediment microcosms. FEMS Microbiol. Lett. 2010, 302, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Postec, A.; Tapia, N.; Bernadac, A.; Joseph, M.; Davidson, S.; Wu, L.-F.; Ollivier, B.; Pradel, N. Magnetotactic bacteria in microcosms originating from the French Mediterranean Coast subjected to oil industry activities. Microb. Ecol. 2012, 63, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Simmons, S.L.; Edwards, K.J.; DeLong, E. Geobiology of Magnetotactic Bacteria. In Magnetoreception and Magnetosomes in Bacteria, 1st ed.; Schüler, D., Ed.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 77–102. [Google Scholar]
- Schüler, D.; Baeuerlein, E. Iron-limited growth and kinetics of iron uptake in Magnetospirillum gryphiswaldense. Arch. Microbiol. 1996, 166, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The Neighbor-joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Bazylinski, D.A.; Dean, A.J.; Williams, T.J.; Long, L.K.; Middleton, S.L.; Dubbels, B.L. Chemolithoautotrophy in the marine, magnetotactic bacterial strains MV-1 and MV-2. Arch. Microbiol. 2004, 182, 373–387. [Google Scholar] [CrossRef] [PubMed]
- Santoro, A.E.; Buchwald, C.; McIlvin, M.R.; Casciotti, K.L. Isotopic signature of N2O produced by marine ammonia-oxidizing archaea. Science 2011, 333, 1282–1285. [Google Scholar] [CrossRef] [PubMed]
- Ji, B.; Zhang, S.-D.; Arnoux, P.; Rouy, Z.; Alberto, F.; Philippe, N.; Murat, D.; Zhang, W.-J.; Rioux, J.-B.; Ginet, N.; et al. Comparative genomic analysis provides insights into the evolution and niche adaptation of marine Magnetospira sp. QH-2 strain. Environ. Microbiol. 2014, 16, 525–544. [Google Scholar] [CrossRef] [PubMed]
- Schübbe, S.; Williams, T.J.; Xie, G.; Kiss, H.E.; Brettin, T.S.; Martinez, D.; Ross, C.A.; Schüler, D.; Cox, B.L.; Nealson, K.H.; et al. Complete genome sequence of the chemolithoautotrophic marine magnetotactic coccus strain MC-1. Appl. Environ. Microbiol. 2009, 75, 4835–4852. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.-Y.; Zhou, K.; Pan, H.-M.; Yue, H.-D.; Jiang, M.; Xiao, T.; Wu, L.-F. Two Genera of Magnetococci with Bean-like Morphology from Intertidal Sediments of the Yellow Sea, China. Appl. Environ. Microbiol. 2012, 78, 5606–5611. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.-Y.; Zhou, K.; Pan, H.-M.; Du, H.-J.; Chen, Y.-R.; Zhang, R.; Ye, W.; Lu, C.; Xiao, T.; Wu, L.-F.; et al. Novel Rod-shaped Magnetotactic Bacteria Belonging to Alphaproteobacteria. Appl. Environ. Microbiol. 2013, 79, 3137–3140. [Google Scholar] [CrossRef] [PubMed]
- Lins, U.; Freitas, F.; Keim, C.N.; de Barros, H.L.; Esquivel, D.M.S.; Farina, M. Simple Homemade Apparatus for Harvesting Uncultured Magnetotactic Microorganisms. Braz. J. Microbiol. 2003, 34, 111–116. [Google Scholar] [CrossRef]
- Wolfe, R.; Thauer, R.K.; Pfennig, N. A “capillary racetrack” method for isolation of magnetotactic bacteria. FEMS Microbiol. Lett. 1987, 45, 31–35. [Google Scholar]
- Frankel, R.B.; Bazylinski, D.A. Structure and function of magnetosomes in magnetotactic bacteria. In Design and Processing of Materials by Biomimetics; Sarikaya, M., Aksay, I., Eds.; American Institute of Physics: New York, NY, USA, 1994; pp. 199–216. [Google Scholar]
- Yan, L.; Zhang, S.; Chen, P.; Liu, H.; Yin, H.; Li, H. Magnetotactic bacteria, magnetosomes and their application. Microbiol. Res. 2012, 167, 507–519. [Google Scholar] [CrossRef] [PubMed]
- Moisescu, C.; Ardelean, I.I.; Benning, L.G. The effect and role of environmental conditions on magnetosome synthesis. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Balkwill, D.L.; Maratea, D.; Blakemore, R.P. Ultrastructure of a magnetotactic spirillum. J. Bacteriol. 1980, 141, 1399–1408. [Google Scholar] [PubMed]
- Moisescu, C.; Bonneville, S.; Tobler, D.J.; Ardelean, I.; Benning, L.G. Controlled biomineralization of magnetite (Fe3O4) by Magnetospirillum gryphiswaldense. Mineral. Mag. 2008, 72, 333–336. [Google Scholar] [CrossRef]
- Geelhoed, J.S.; Kleerebezem, R.; Sorokin, D.Y.; Stams, A.J.M.; van Loosdrecht, M.C.M. Reduced inorganic sulfur oxidation supports autotrophic and mixotrophic growth of Magnetospirillum strain J10 and Magnetospirillum gryphiswaldense. Environ. Microbiol. 2010, 12, 1031–1040. [Google Scholar] [CrossRef] [PubMed]
- Bazylinski, D.A.; Frankel, R.B.; Jannasch, H.W. Anaerobic magnetite production by a marine magnetotatic bacterium. Nature 1988, 334, 518–519. [Google Scholar] [CrossRef]
- Meldrum, F.C.; Mann, S.; Heywood, B.R.; Frankel, R.B.; Bazylinski, D.A. Electron microscopy study of magnetosomes in two cultured vibrioid magnetotactic bacteria. Proc. R. Soc. Lond. B Biol. Sci. 1993, 251, 231–236. [Google Scholar] [CrossRef]
- Byrne, M.E.; Ball, D.A.; Guerquin-Kern, J.-L.; Rouillere, I.; Wu, T.-D.; Downing, K.H.; Valie, H.; Komeili, A. Desulfovibrio magneticus RS-1 contains an iron- and phosphorus-rich organelle distinct from its bullet-shaped magnetosomes. Proc. Natl. Acad. Sci. USA 2010, 107, 12263–12268. [Google Scholar] [CrossRef] [PubMed]
- Pósfai, M.; Lefèvre, C.T.; Trubitsyn, D.; Bazylinski, D.A.; Frankel, R.B. Phylogenetic significance of composition and crystal morphology of magnetosome minerals. Front. Microbiol. 2014, 4. [Google Scholar] [CrossRef]
- Pósfai, M.; Buseck, P.R.; Bazylinski, D.A.; Frankel, R.B. Iron sulfides from magnetotactic bacteria: Structure, composition, and phase transitions. Am. Mineral. 1998, 83, 1469–1481. [Google Scholar]
- Komeili, A.; Li, Z.; Newman, D.K.; Jensen, G.J. Magnetosomes are cell membrane invaginations organized by the actin-like protein MamK. Science 2006, 311, 242–245. [Google Scholar] [CrossRef] [PubMed]
- Murat, D.; Quinlan, A.; Vali, H.; Komeili, A. Comprehensive genetic dissection of the magnetosome gene island reveals the step-wise assembly of a prokaryotic organelle. Proc. Natl. Acad. Sci. USA 2010, 107, 5593–5598. [Google Scholar] [CrossRef] [PubMed]
- Faivre, D.; Böttger, L.H.; Matzanke, B.F.; Schüler, D. Intracellular magnetite biomineralization in bacteria proceeds by a distinct pathway involving membrane-bound ferritin and an iron(II) species. Angew. Chem. Int. Ed. Engl. 2007, 46, 8495–8499. [Google Scholar] [CrossRef] [PubMed]
- Richter, M.; Kube, M.; Bazylinski, D.A.; Lombardot, T.; Glöckner, F.O.; Reinhardt, R.; Schüler, D. Comparative genome analysis of four magnetotactic bacteria reveals a complex set of group-specific genes implicated in magnetosome biomineralization and function. J. Bacteriol. 2007, 189, 4899–4910. [Google Scholar] [CrossRef] [PubMed]
- Lohße, A.; Ullrich, S.; Katzmann, E.; Borg, S.; Wanner, G.; Richter, M.; Voigt, B.; Schweder, T.; Schüler, D. Functional analysis of the magnetosome island in Magnetospirillum gryphiswaldense: The mamAB operon is sufficient for magnetite biomineralization. PLoS One 2011, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheffel, A.; Gruska, M.; Faivre, D.; Linaroudis, A.; Plitzko, J.M.; Schüler, D. An acidic protein aligns magnetosomes along a filamentous structure in magnetotactic bacteria. Nature 2006, 7080, 110–114. [Google Scholar] [CrossRef]
- Uebe, R.; Junge, K.; Henn, V.; Poxleitner, G.; Katzmann, E.; Plitzko, J.M.; Zarivach, R.; Kasama, T.; Wanner, G.; Pósfai, M.; et al. The cation diffusion facilitator proteins MamB and MamM of Magnetospirillum gryphiswaldense have distinct and complex functions, and are involved in magnetite biomineralization and magnetosome membrane assembly. Mol. Microbiol. 2011, 82, 818–835. [Google Scholar] [CrossRef] [PubMed]
- Quinlan, A.; Murat, D.; Vali, H.; Komeili, A. The HtrA/DegP family protease MamE is a bifunctional protein with roles in magnetosome protein localization and magnetite biomineralization. Mol. Microbiol. 2011, 80, 1075–1087. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Li, R.; Peng, T.; Zhang, Y.; Jiang, W.; Li, Y.; Li, J. mamO and mamE genes are essential for magnetosome crystal biomineralization in Magnetospirillum gryphiswaldense MSR-1. Res. Microbiol. 2010, 161, 701–705. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Li, J.; Liu, J.; Yang, J.; Jiang, W.; Tian, J.; Li, Y.; Pan, Y.; Li, J. Deletion of the ftsZ-like gene results in the production of superparamagnetic magnetite magnetosomes in Magnetospirillum gryphiswaldense. J. Bacteriol. 2010, 192, 1097–1105. [Google Scholar] [CrossRef] [PubMed]
- Scheffel, A.; Gärdes, A.; Grünberg, K.; Wanner, G.; Schüler, D. The major magnetosome proteins MamGFDC are not essential for magnetite biomineralization in Magnetospirillum gryphiswaldense but regulate the size of magnetosome crystals. J. Bacteriol. 2008, 190, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Arakaki, A.; Masuda, F.; Amemiya, Y.; Tanaka, T.; Matsunaga, T. Control of the morphology and size of magnetite particles with peptides mimicking the Mms6 protein from magnetotactic bacteria. J. Colloid Interface Sci. 2010, 343, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Ginet, N.; Pardoux, R.; Adryanczyk, G.; Garcia, D.; Brutesco, C.; Pignol, D. Single-step production of a recyclable nanobiocatalyst for organophosphate pesticides biodegradation using functionalized bacterial magnetosomes. PLoS One 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Alphandéry, E.; Faure, S.; Seksek, O.; Guyot, F.; Chebbi, I. Chains of magnetosomes extracted from AMB-1 magnetotactic bacteria for application in alternative magnetic field cancer therapy. ACS Nano 2011, 5, 6279–6296. [Google Scholar] [CrossRef] [PubMed]
- Prozorov, T.; Bazylinski, D.A.; Mallapragada, S.K.; Prozorov, R. Novel magnetic nanomaterials inspired by magnetotactic bacteria: Topical review. Mater. Sci. Eng. R Rep. 2013, 74, 133–172. [Google Scholar] [CrossRef]
- Benoit, M.R.; Mayer, D.; Barak, Y.; Chen, I.Y.; Hu, W.; Cheng, Z.; Wang, S.X.; Spielman, D.M.; Gambhir, S.S.; Matin, A.; et al. Visualizing implanted tumors in mice with magnetic resonance imaging using magnetotactic bacteria. Clin. Cancer Res. 2009, 15, 5170–5177. [Google Scholar] [CrossRef] [PubMed]
- Nudelman, H.; Zarivach, R. Structure prediction of magnetosome-associated proteins. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Komeili, A.; Vali, H.; Beveridge, T.J.; Newman, D.K. Magnetosome vesicles are present before magnetite formation, and MamA is required for their activation. Proc. Natl. Acad. Sci. USA 2004, 101, 3839–3844. [Google Scholar] [CrossRef] [PubMed]
- Zeytuni, N.; Ozyamak, E.; Ben-Harush, K.; Davidov, G.; Levin, M.; Gat, Y.; Moyal, T.; Brik, A.; Komeili, A.; Zarivach, R.; et al. Self-recognition mechanism of MamA, a magnetosome-associated TPR-containing protein, promotes complex assembly. Proc. Natl. Acad. Sci. USA 2011, 108, E480–E487. [Google Scholar] [CrossRef] [PubMed]
- Raschdorf, O.; Müller, F.D.; Pósfai, M.; Plitzko, J.M.; Schüler, D. The magnetosome proteins MamX, MamZ and MamH are involved in redox control of magnetite biomineralization in Magnetospirillum gryphiswaldense. Mol. Microbiol. 2013, 89, 872–886. [Google Scholar] [CrossRef] [PubMed]
- Fdez-Gubieda, M.L.; Muela, A.; Alonso, J.; Garcıa-Prieto, A.; Olivi, L.; Fernández-Pacheco, R.; Barandiarán, J.M. Magnetite Biomineralization in Magnetospirillum gryphiswaldense: Time-resolved magnetic and structural studies. ACS Nano 2013, 7, 3297–3305. [Google Scholar] [CrossRef] [PubMed]
- Katzmann, E.; Scheffel, A.; Gruska, M.; Plitzko, J.M.; Schüler, D. Loss of the actin-like protein MamK has pleiotropic effects on magnetosome formation and chain assembly in Magnetospirillum gryphiswaldense. Mol. Microbiol. 2010, 77, 208–224. [Google Scholar] [CrossRef] [PubMed]
- Siponen, M.I.; Adryanczyk, G.; Ginet, N.; Arnoux, P.; Pignol, D. Magnetochrome: A c-type cytochrome domain specific to magnetotatic bacteria. Biochem. Soc. Trans. 2012, 40, 1319–1323. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Arakaki, A.; Matsunaga, T. Identification and functional characterization of liposome tubulation protein from magnetotactic bacteria. Mol. Microbiol. 2010, 76, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Mazuyama, E.; Arakaki, A.; Matsunaga, T. MMS6 protein regulates crystal morphology during nano-sized magnetite biomineralization in vivo. J. Biol. Chem. 2011, 286, 6386–6392. [Google Scholar] [CrossRef] [PubMed]
- Schüler, D. Genetics and cell biology of magnetosome formation in magnetotactic bacteria. FEMS Microbiol. Rev. 2008, 32, 654–672. [Google Scholar] [CrossRef] [PubMed]
- Bazylinski, D.A.; Williams, T.J. Ecophysiology of Magnetotactic Bacteria. In Magnetoreception and Magnetosomes in Bacteria, 1st ed.; Schüler, D., Ed.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 37–75. [Google Scholar]
- Mandernack, K.W.; Bazylinski, D.A.; Shanks, W.C., III; Bullen, T.D. Oxygen and iron isotope studies of magnetite produced by magnetotactic bacteria. Science 1999, 285, 1892–1896. [Google Scholar] [CrossRef] [PubMed]
- Schüler, D.; Baeuerlein, E. Dynamics of iron uptake and Fe3O4 biomineralization during aerobic and microaerobic growth of Magnetospirillum gryphiswaldense. J. Bacteriol. 1998, 180, 159–162. [Google Scholar] [PubMed]
- Schultheiss, D.; Schüler, D. Development of a genetic system for Magnetospirillum gryphiswaldense. Arch. Microbiol. 2003, 179, 89–94. [Google Scholar] [PubMed]
- Abreu, F.; Sousa, A.A.; Aronova, M.A.; Kim, Y.; Cox, D.; Leapman, R.D.; Andrade, L.R.; Kachar, B.; Bazylinski, D.A.; Lins, U.; et al. Cryo-electron tomography of the magnetotactic vibrio Magnetovibrio blakemorei: Insights into the biomineralization of prismatic magnetosomes. J. Struct. Biol. 2013, 181, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-D.; Takeyama, H.; Matsunaga, T. Iron feeding optimization and plasmid stability in production of recombinant bacterial magnetic particles by Magnetospirillum magneticum AMB-1 in fed-batch culture. J. Biosci. Bioeng. 2001, 91, 213–216. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.-B.; Zhao, F.; Tang, T.; Jiang, W.; Tian, J.; Li, Y.; Li, J.-L. High-yield growth and magnetosome formation by Magnetospirillum gryphiswaldense MSR-1 in an oxygen-controlled fermentor supplied solely with air. Appl. Microbiol. Biotechnol. 2008, 79, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Alphandéry, E.; Amor, M.; Guyot, F.; Chebbi, I. The effect of iron-chelating agents on Magnetospirillum magneticum strain AMB-1: Stimulated growth and magnetosome production and improved magnetosome heating properties. Appl. Microbiol. Biotechnol. 2012, 96, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ding, Y.; Jiang, W.; Tian, J.; Li, Y.; Li, J. A mutation upstream of an ATPase gene significantly increases magnetosome production in Magnetospirillum gryphiswaldense. Appl. Microbiol. Biotechnol. 2008, 81, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, G.R.; Guo, F.F.; Jiang, W.; Li, Y.; Li, L.J. Large-scale production of magnetosomes by chemostat culture of Magnetospirillum gryphiswaldense at high cell density. Microb. Cell Factoriels 2010, 9, 99–106. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.; Jiang, W.; Li, Y.; Li, J. Semicontinuous culture of Magnetospirillum gryphiswaldense MSR-1 cells in an autofermentor by nutrient-balanced and isosmotic feeding strategies. Appl. Environ. Microbiol. 2011, 77, 5851–5856. [Google Scholar] [CrossRef] [PubMed]
- Alphandéry, E.; Ding, Y.; Ngo, A.T.; Wang, Z.L.; Wu, L.F.; Pileni, M.P. Assemblies of aligned magnetotactic bacteria and extracted magnetosomes: What is the main factor responsible for the magnetic anisotropy? ACS Nano 2009, 3, 1539–1547. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Liu, Y.; Chen, Y.; Tang, T.; Jiang, W.; Li, Y.; Li, J. A novel rapid and continuous procedure for large-scale purification of magnetosomes from Magnetospirillum gryphiswaldense. Appl. Microbiol. Biotechnol. 2011, 90, 1277–1283. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.-B.; Duan, J.-H.; Dai, S.-L.; Ren, J.; Zhang, Y.-D.; Tian, J.-S.; Li, Y. In vitro and in vivo antitumor effects of doxorubicin loaded with bacterial magnetosomes (DBMs) on H22 cells: The magnetic bio-nanoparticles as drug carriers. Cancer Lett. 2007, 258, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-D.; Takeyama, H.; Tanaka, T.; Matsunaga, T. Effects of growth medium composition, iron sources and atmospheric oxygen concentrations on production of luciferase-bacterial magnetic particle complex by a recombinant Magnetospirillum magneticum AMB-1. Enzym. Microb. Technol. 2001, 29, 13–19. [Google Scholar] [CrossRef]
- Lang, C.; Schüler, D. Expression of green fluorescent protein fused to magnetosome proteins in microaerophilic magnetotactic bacteria. Appl. Environ. Microbiol. 2008, 74, 4944–4953. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Yoo, C.-H.; Lee, J.; Cha, M. Cell response induced by internalized bacterial magnetic nanoparticles under an external static magnetic field. Biomaterials 2012, 33, 5650–5657. [Google Scholar] [CrossRef] [PubMed]
- Salata, O.V. Applications of nanoparticles in biology and medicine. J. Nanobiotechnol. 2004, 2, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Pankhurst, Q.A.; Connolly, J.; Jones, S.; Dobson, J. Applications of magnetic nanoparticles in biomedicine. J. Phys. D Appl. Phys. 2003, 36, R167–R181. [Google Scholar] [CrossRef]
- Konishi, Y.; Nomura, T.; Mizoe, K. A new synthesis route from spent sulfuric acid pickling solution to ferrite nanoparticles. Hydrometallurgy 2004, 74, 57–65. [Google Scholar] [CrossRef]
- Liu, Z.L.; Wang, X.; Yao, K.L.; Du, G.H.; Lu, Q.H.; Ding, Z.H.; Tao, J.; Ning, Q.; Luo, X.P.; Tian, D.Y.; et al. Synthesis of magnetite nanoparticles in W/O microemulsion. J. Mater. Sci. 2004, 39, 2633–2636. [Google Scholar] [CrossRef]
- Franger, S.; Berthet, P.; Berthon, J. Electrochemical synthesis of Fe3O4 nanoparticles in alkaline aqueous solutions containing complexing agents. J. Solid State Electrochem. 2004, 8, 218–223. [Google Scholar] [CrossRef]
- Wu, M.; Xiong, Y.; Jia, Y.; Niu, H.; Qi, H.; Ye, J.; Chen, Q. Magnetic field-assisted hydrothermal growth of chain-like nanostructure of magnetite. Chem. Phys. Lett. 2005, 401, 374–379. [Google Scholar] [CrossRef]
- Thapa, D.; Palkar, V.; Krupp, M.; Malik, S. Properties of magnetite nanoparticles synthesized through a novel chemical route. Mater. Lett. 2004, 58, 2692–2694. [Google Scholar] [CrossRef]
- Alphandéry, E.; Chebbi, I.; Guyot, F.; Durand-Dubief, M. Use of bacterial magnetosomes in the magnetic hyperthermia treatment of tumours: A review. Int. J. Hyperth. 2013, 29, 801–809. [Google Scholar] [CrossRef]
- Arakaki, A.; Nakazawa, H.; Nemoto, M.; Mori, T.; Matsunaga, T. Formation of magnetite by bacteria and its application. J. R. Soc. Interface 2008, 5, 977–999. [Google Scholar] [PubMed]
- Tanaka, T.; Matsunaga, T. Detection of HbA(1c) by boronate affinity immunoassay using bacterial magnetic particles. Biosens. Bioelectron. 2001, 16, 1089–1094. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, N.; Hashimoto, K.; Matsunaga, T. Immunoassay method for the determination of immunoglobulin G using bacterial magnetic particles. Anal. Chem. 1991, 63, 268–272. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Kokuryu, Y.; Matsunaga, T. Novel method for selection of antimicrobial peptides from a phage display library by use of bacterial magnetic particles. Appl. Environ. Microbiol. 2008, 74, 7600–7606. [Google Scholar] [CrossRef] [PubMed]
- Yoza, B.; Arakaki, A.; Maruyama, K.; Takeyama, H.; Matsunaga, T. Fully automated DNA extraction from blood using magnetic particles modified with a hyperbranched polyamidoamine dendrimer. J. Biosci. Bioeng. 2003, 95, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Ohuchi, S.; Schüler, D. In vivo display of a multisubunit enzyme complex on biogenic magnetic nanoparticles. Appl. Environ. Microbiol. 2009, 75, 7734–7738. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, T.; Togo, H.; Kikuchi, T.; Tanaka, T. Production of luciferase-magnetic particle complex by recombinant Magnetospirillum sp. AMB-1. Biotechnol. Bioeng. 2000, 70, 704–709. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, T.; Kato, F.; Takeyama, H.; Nakai, M.; Yakabe, Y.; Matsunaga, T. Development of a novel method for screening of estrogenic compounds using nano-sized bacterial magnetic particles displaying estrogen receptor. Anal. Chim. Acta 2005, 532, 105–111. [Google Scholar] [CrossRef]
- Schüler, D.; Frankel, R.B. Bacterial magnetosomes: Microbiology, biomineralization and biotechnological applications. Appl. Microbiol. Biotechnol. 1999, 52, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Bahaj, A.S.; James, P.A.B.; Moeschler, F.D. Wastewater treatment by bio-magnetic separation: A comparison of iron oxide and iron sulphide biomass recovery. Water Sci. Technol. 1998, 38, 311–317. [Google Scholar] [CrossRef]
- Bahaj, A.S.; Croudace, I.W.; James, P.A.B.; Moeschler, F.D.; Warwick, P.E. Continuous radionuclide recovery from wastewater using magnetotactic bacteria. J. Magn. Magn. Mater. 1998, 184, 241–244. [Google Scholar] [CrossRef]
- Arakaki, A.; Takeyama, H.; Tanaka, T.; Matsunaga, T. Cadmium recovery by a sulfate-reducing magnetotactic bacterium, Desulfovibrio magneticus RS-1, using magnetic separation. Appl. Biochem. Biotechnol. 2002, 98–100, 833–840. [Google Scholar] [CrossRef] [PubMed]
- Cai, F.; Li, J.; Sun, J.; Ji, Y. Biosynthesis of gold nanoparticles by biosorption using Magnetospirillum gryphiswaldense MSR-1. Chem. Eng. J. 2011, 175, 70–75. [Google Scholar] [CrossRef]
- Matsunaga, T. Applications of bacterial magnets. Trends Biotechnol. 1991, 9, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Martel, S. Preliminary investigation of bio-carriers using magnetotactic bacteria. Conf. Proc. IEEE Eng. Med. Biol. Soc. 2006, 1, 3415–3418. [Google Scholar] [PubMed]
- Felfoul, O.; Mohammadi, M.; Martel, S. Magnetic resonance imaging of Fe3O4 nanoparticles embedded in living magnetotactic bacteria for potential use as carriers for in vivo applications. Conf. Proc. IEEE Eng. Med. Biol. Soc. 2007, 2007, 1463–1466. [Google Scholar] [PubMed]
- Yoshino, T.; Nishimura, T.; Mori, T.; Suzuki, S.; Kambara, H.; Takeyama, H.; Matsunaga, T. Nano-sized bacterial magnetic particles displaying pyruvate phosphate dikinase for pyrosequencing. Biotechnol. Bioeng. 2009, 103, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Wacker, R.; Ceyhan, B.; Alhorn, P.; Schueler, D.; Lang, C.; Niemeyer, C.M. Magneto immuno-PCR: A novel immunoassay based on biogenic magnetosome nanoparticles. Biochem. Biophys. Res. Commun. 2007, 357, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, T.; Maeda, Y.; Yoshino, T.; Takeyama, H.; Takahashi, M.; Ginya, H.; Aasahina, J.; Tajima, H. Fully automated immunoassay for detection of prostate-specific antigen using nano-magnetic beads and micro-polystyrene bead composites, “Beads on Beads”. Anal. Chim. Acta 2007, 597, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, T.; Takahashi, M.; Yoshino, T.; Kuhara, M.; Takeyama, H. Magnetic separation of CD14+ cells using antibody binding with protein A expressed on bacterial magnetic particles for generating dendritic cells. Biochem. Biophys. Res. Commun. 2006, 350, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- Amemiya, Y.; Tanaka, T.; Yoza, B.; Matsunaga, T. Novel detection system for biomolecules using nano-sized bacterial magnetic particles and magnetic force microscopy. J. Biotechnol. 2005, 120, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Saiyed, Z.; Telang, S.; Ramchand, C. Application of magnetic techniques in the field of drug discovery and biomedicine. Biomagn. Res. Technol. 2003, 1. [Google Scholar] [CrossRef] [Green Version]
- Tang, T.; Zhang, L.; Gao, R.; Dai, Y.; Meng, F.; Li, Y. Fluorescence imaging and targeted distribution of bacterial magnetic particles in nude mice. Appl. Microbiol. Biotechnol. 2012, 94, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.-S.; Wang, D.; Zhou, C.; Ma, W.; Zhang, Y.-Q.; Liu, B.; Zhang, S. Bacterial magnetic particles as a novel and efficient gene vaccine delivery system. Gene Ther. 2012, 19, 1187–1195. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Li, S.-Y.; Yang, Y.; Zhao, F.-M.; Huang, J.; Chang, J. Research on the structure and performance of bacterial magnetic nanoparticles. J. Biomater. Appl. 2008, 22, 433–448. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.-B.; Wang, Z.-L.; Duan, J.-H.; Ren, J.; Yang, X.-D.; Dai, S.-L.; Li, Y. Targeted distribution of bacterial magnetosomes isolated from Magnetospirillum gryphiswaldense MSR-1 in healthy Sprague-Dawley rats. J. Nanosci. Nanotechnol. 2009, 9, 1881–1885. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.-T.; Liu, J.; Tong, J.-Q.; Tang, T.; Kong, W.-C.; Wang, X.-W.; Li, Y.; Tang, J.-T. Heating effect and biocompatibility of bacterial magnetosomes as potential materials used in magnetic fluid hyperthermia. Prog. Nat. Sci. Mater. Int. 2012, 22, 31–39. [Google Scholar] [CrossRef]
- Food and Drug Administration. Drugs Development and Approval Process. Available online: http://www.fda.gov/Drugs/DevelopmentApprovalProcess/HowDrugsareDevelopedandApproved/ApprovalApplications/default.htm (accessed on 4 December 2014).
- Food and Drug Administration. Information on Gadolinium-Based Contrast Agents. Available online: http://www.fda.gov/drugs/drugsafety/postmarketdrugsafetyinformationforpatientsandproviders/ucm142882.htm (accessed on 4 December 2014).
- Abreu, F.; Morillo, V.; Nascimento, F.F.; Werneck, C.; Cantão, M.E.; Ciapina, L.P.; de Almeida, L.G.P.; Lefèvre, C.T.; Bazylinski, D.A.; de Vasconcelos, A.T.R.; et al. Deciphering unusual uncultured magnetotactic multicellular prokaryotes through genomics. ISME J. 2014, 8, 1055–1068. [Google Scholar] [CrossRef] [PubMed]
- Carini, P.; Steindler, L.; Beszteri, S.; Giovannoni, S.J. Nutrient requirements for growth of the extreme oligotroph “Candidatus Pelagibacter ubique” HTCC1062 on a defined medium. ISME J. 2013, 7, 592–602. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Medema, M.H.; Kazempour, D.; Fischbach, M.A.; Breitling, R.; Takano, E.; Weber, T. antiSMASH 2.0—A versatile platform for genome mining of secondary metabolite producers. Nucleic Acids Res. 2013, 41, W204–W212. [Google Scholar] [CrossRef] [PubMed]
- Letzel, A.-C.; Pidot, S.J.; Hertweck, C. A genomic approach to the cryptic secondary metabolome of the anaerobic world. Nat. Prod. Rep. 2013, 30, 392–428. [Google Scholar] [CrossRef] [PubMed]
- Desriac, F.; Jégou, C.; Balnois, E.; Brillet, B.; le Chevalier, P.; Fleury, Y. Antimicrobial peptides from marine proteobacteria. Mar. Drugs 2013, 11, 3632–3660. [Google Scholar] [CrossRef] [PubMed]
- Donadio, S.; Monciardini, P.; Sosio, M. Polyketide synthases and nonribosomal peptide synthetases: The emerging view from bacterial genomics. Nat. Prod. Rep. 2007, 24, 1073–1109. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, H.; Arakaki, A.; Narita-Yamada, S.; Yashiro, I.; Jinno, K.; Aoki, N.; Tsuruyama, A.; Okamura, Y.; Tanikawa, S.; Fujita, N.; et al. Whole genome sequence of Desulfovibrio magneticus strain RS-1 revealed common gene clusters in magnetotactic bacteria. Genome Res. 2009, 19, 1801–1808. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, T.; Okamura, Y.; Fukuda, Y.; Wahyudi, A.T.; Murase, Y.; Takeyama, H. Complete genome sequence of the facultative anaerobic magnetotactic bacterium Magnetospirillum sp. strain AMB-1. DNA Res. 2005, 12, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, Q.; Zhang, W.; Wang, Y.; Li, L.; Wen, T.; Zhang, T.; Zhang, Y.; Xu, J.; Hu, J.; et al. Complete Genome Sequence of Magnetospirillum gryphiswaldense. Genome Announc. 2014, 2, e00171-14. [Google Scholar] [PubMed]
- Grouzdev, D.S.; Dziuba, M.V.; Sukhacheva, M.S.; Mardanov, A.V.; Beletskiy, A.V.; Kuznetsov, B.B.; Skryabin, K.G. Draft Genome Sequence of Magnetospirillum sp. Strain SO-1, a Freshwater Magnetotactic Bacterium Isolated from the Ol’khovka River, Russia. Genome Announc. 2014, 2, e00235-14. [Google Scholar] [CrossRef] [PubMed]
- Juhas, M.J.; van der Meer, R.; Gaillard, M.; Harding, R.M.; Hood, D.W.; Crook, D.W. Genomic islands: Tools of bacterial horizontal gene transfer and evolution. FEMS Microbiol. Rev. 2009, 33, 376–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, A.C.; Xu, Y.; Lu, L.; Kersten, R.D.; Shao, Z.; Al-Suwailem, A.M.; Dorrestein, P.C.; Qian, P.-Y.; Moore, B.S. Biosynthetic multitasking facilitates thalassospiramide structural diversity in marine bacteria. J. Am. Chem. Soc. 2013, 135, 1155–1162. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, D.; Hevel, J.; Moore, R.; Moore, B. Sequence analysis and biochemical characterization of the nostopeptolide A biosynthetic gene cluster from Nostoc sp. GSV224. Gene 2003, 311, 171–180. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Araujo, A.C.V.; Abreu, F.; Silva, K.T.; Bazylinski, D.A.; Lins, U. Magnetotactic Bacteria as Potential Sources of Bioproducts. Mar. Drugs 2015, 13, 389-430. https://doi.org/10.3390/md13010389
Araujo ACV, Abreu F, Silva KT, Bazylinski DA, Lins U. Magnetotactic Bacteria as Potential Sources of Bioproducts. Marine Drugs. 2015; 13(1):389-430. https://doi.org/10.3390/md13010389
Chicago/Turabian StyleAraujo, Ana Carolina V., Fernanda Abreu, Karen Tavares Silva, Dennis A. Bazylinski, and Ulysses Lins. 2015. "Magnetotactic Bacteria as Potential Sources of Bioproducts" Marine Drugs 13, no. 1: 389-430. https://doi.org/10.3390/md13010389
APA StyleAraujo, A. C. V., Abreu, F., Silva, K. T., Bazylinski, D. A., & Lins, U. (2015). Magnetotactic Bacteria as Potential Sources of Bioproducts. Marine Drugs, 13(1), 389-430. https://doi.org/10.3390/md13010389