
Marine Peptides and Their Anti-Infective Activities


Abstract
:1. Introduction
2. Marine Organisms and Microorganisms
2.1. Sponges
2.2. Algae
2.3. Microorganism
2.4. Fish
3. Anti-Infective Marine Peptides

| Activity | Name of Peptide | Source of Original Peptides | Pharmacologic Activity | Inhibition Concentrations | References |
|---|---|---|---|---|---|
| Antibacterial | Aurelin (1) | Jellyfish: Aurelia aurita | Escheichia coli inhibition | 7.7 μg/mL (MIC) | [31] |
| Antibacterial | Arenicin-1 (2) | Polychaete: Arenicola marina | Pseudomomas aeruginosa and Staphylococcus aureus inhibition | 2 μg/mL (MIC) | [32,33] |
| Antibacterial | Tauramamide (3) | Bacterium: Brevibacillus laterosporus | Enterococcus sp. inhibition | 0.1 μg/mL (MIC) | [34] |
| Antibacterial | Hepcidin (4) | Fish: Oreochromis mossambicus | Listeria monocytogenes, S. aureus, and Enterococcus faecium inhibition | 50–100 μg/mL (MIC) | [35] |
| Antibacterial | Scygonadin (5) | Mud crab: Scylla serrata | E. coli, P. aeruginosa, S. aureus, Streptococcus pyogenes inhibition | 50–100 μg/mL (MIC) | [36,37] |
| Antibacterial | Tunichromes (6) | Ascidian: Ascidia nigra | Enterococcus sp. inhibition | 0.1 μg/mL (MIC) | [38,39] |
| Antibacterial | Bacillistatins 1 (7), 2 (8) | Bacterium: Bacillus silvestris | Streptococcus. pneumonia inhibition | 0.5–2 μg/mL (GI50) | [40] |
| Antibacterial | Nacardiopsis thiopeptide TP-1161 (9) | Bacterium: Nocardiopsis sp | Vancomycin-resistant Enterococcus faecium inhibition | 1 μg/mL (MIC) | [41] |
| Antibacterial | Centrocins 1 (10), 2 (11) | Sea urchin: Strongylocentrotus droebachiensis | Corynebacterium glutamicum, S. aureus, Listonella anguillarum, E. coli inhibition | 1.3–5 μM (IC50) | [42] |
| Antibacterial | Halocyntin (12) | Ascidian: Halocynthia papillosa | Micrococcus luteus, Bacillus megaterium, Aerococcus viridans, S. aureus, Enterococcus faecalis inhibition | 0.39–50 μM (MBC) | [43] |
| Antibacterial | Papillosin (13) | Ascidian: Halocynthia papillosa | M. luteus, B. megaterium, Aerococcus viridans, S. aureus, Enterococcus faecalis inhibition | 0.05–6.25 μM (MBC) | [43] |
| Antibacterial | Hyastain (14) | Spider crab: Hyas araneus | E. coli, Corynebacterium glutamicum, S. aureus inhibition | 0.4–12.5 μM (MIC) | [44] |
| Antibacterial | Indigoidine (15) | Bacterium: Phaeobacter sp. | Vibrio fischeri inhibition | ND | [45] |
| Antibacterial | Unnarmicins A (16), C (17) | Bacterium: Photobacterium sp. | Pseudovibrio sp. inhibition | 7–18 μg/disk (disk) | [46] |
| Antibacterial | Ngercheumicins A–D (18–21) | Bacterium: Photobacterium sp. | Gram negative strain inhibition | ND. | [47] |
| Antibacterial | Solonamidine A (22), B (23) | Bacterium: Photobacterium sp | S. aureus, Methicillin-resistant S. aureus (MRSA)inhibition | ND. | [48] |
| Antibacterial | Cyclo-peptides (24) | Bacterium: Pseudomonas sp. | S. aureus, M. luteus, B. subtilis, E. coli, V. anguillarum inhibition | ND | [49] |
| Antibacterial | Ariakemicins A (25), B (26) | Bacterium: Rapidithrix sp. | Brevibacterium sp., S. aureus, B. subtilis inhibition | 0.46–80 μg/mL (MIC) | [50] |
| Antibacterial | Damicornin (27) | Coral: Pocillopora damicorins | M. luteus, B. megaterium, S. aureus, Brevibacterium stationis, Microbacterium maritypicum, Fusarium oxysporum inhibition | 1.25–20 μM (MIC) | [51] |
| Antibacterial | Clavanis (28) | Tunicate: Styela clava | S. aureus, Klebsiella pneumonia, P. aeruginosa inhibition | ND | [52] |
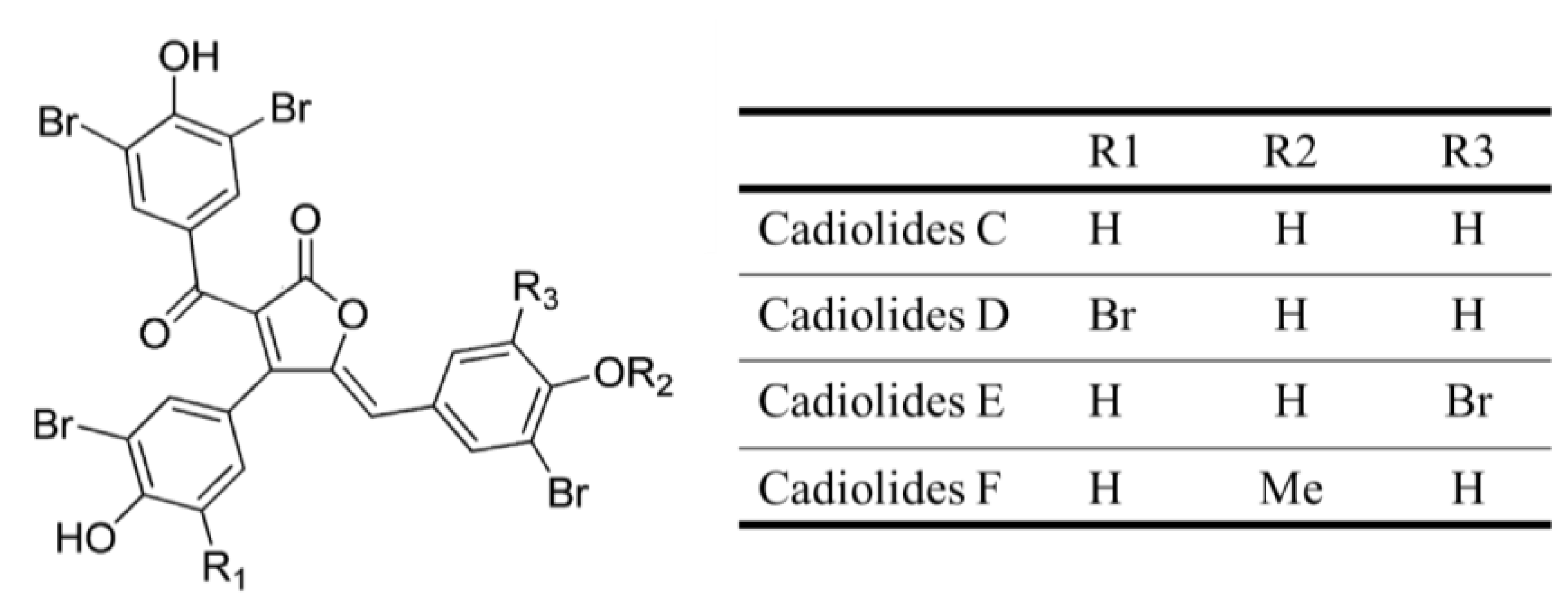
| Antibacterial | Cadiolides C–F (29–32) | Tunicate: Pseudodistoma antinboja | S. aureus, S. epidermidis, Kocuria rhizophila and B. subtilis, methicillin-sensitive S. aureus (MSSA), MRSAinhibition | 0.13–12.5 μg/mL (MIC) | [53] |
| Antibacterial | Cytosporones B (33), E (34) | Fungus: Leucostoma persoonii | S. aureus USA100, MRSA, MSSA inhibition | 72–78 μM (MIC) | [54] |
| Antibacterial | Anthracimycin (35) | Bacterium: Streptomyces sp. | B. anthracis, Enterococcus facecalis, Streptococcus pneumonia, S. aureus, MSSA, MRSA, vancomycin-resistant S. aureus inhibition | 0.03125–0.25 μg/mL (MIC) | [55] |
| Antifungal | Halocidin (36) | Ascidian: Halocynthia aurantium | Candida albicans inhibition | 1–4 μg/mL(MIC) | [56] |
| Antifungal | Callipeltine J (37), K (38) | Sponge: Latrunculia sp. | C. albicans inhibition | 1 μM (MIC) | [57] |
| Antifungal | Pedein A (39) | Bacterium: Chondromyces pediculatus | Rhizopus glutinis, Saccharomyces cerevisae, C. albicans inhibition | 0.6–1.6 μg/mL (MIC) | [58] |
| Antifungal | Theuellamide F (40), G (41) | Sponge: Theonella sp. | C. albicans inhibition | 2–4.49 μM (IC50) | [59,60] |
| Antifungal | Theopapuamide B (42), C (43) | Sponge: Siliquariaspongia mirabilis | C. albicans inhibition | 1–5 μg/disk (disk) | [61] |
| Antifungal | C(15)-surfactin (44) | Bacterium: B. amyloliquefaciens | C. albicans inhibition | 0.004 μg/mL (MIC) | [62] |
| Antifungal | Anti-CAcyclic lipopeptide (45) | Bacterium: B. amyloliquefaciens | C. tropicalis, Metschnikowia bicuspidata, Sacchromyces cerevisiae, Yarrowia lipolytica inhibition | 7.0 μg/mL (MIC) | [63] |
| Antifungal | Maribasins A (46), B (47) | Bacteriun: B. marinus | Alternaria solani, Fusarium oxysporum, Verticillium alboatrum, F. graminearum, Sclerotium sp., Penicillium sp., Rhizoctonia solani, Colletotrichum sp. inhibition | 25–200 μg/mL (MIC) | [64] |
| Antifungal | Mojavensin A (48) | Bacterium: B. mojavensis | Phytopathogenic fungi inhibition | ND | [65] |
| Antifungal | Kahalalide F (49) | Mollusk: Elysia rufescens | C. albicans, C. neoformans, Aspergillus fumigatus inhibition | 1.53–3.21 μM (IC50) | [66] |
| Antifungal | Miraenamide A (50), B (51) | Bacterium: Paraliomyxa miuraensis | A. niger, Phytophthora capsici, Rhizopus oryzae, C. rugosa, inhibition | 0.4–25 μM (MIC) | [67,68] |
| Antifungal | Callyaerin A (52), E (53) | Sponge: Callyspongia aerizusa | C. albicans inhibition | 5–10 μg/disk(disk) | [69] |
| Antimalarial | Dragomabin (54) | Bacterium: Lyngbya majuscula | Plasmodium falciparum W2 strain inhibition | 6.0 μM (IC50) | [70] |
| Antimalarial | Venturamid A (55), B (56) | Bacterium: Oscillatoria sp. | Plasmodium falciparum W2 strain inhibition | 5.6–8.2 μM (IC50) | [71] |
| Antimalarial | Aerucyamide A–D (57–60) | Bacteriun: Microcystis aeruginosa | Plasmodium falciparum K1 strain inhibition | 0.7 μM (IC50) | [72,73] |
| Antimalarial | Gallinamide A (61) | Bacterium: Schizothrix sp. | Plasmodium falciparum W2 strain inhibition | 8.4 μM (IC50) | [74] |
| Antimalarial | Lagunamide A (62), B (63) | Bacterium: Lyngbya majuscula | Plasmodium falciparum NF54 strain inhibition | 0.19–0.91 μM (IC50) | [75] |
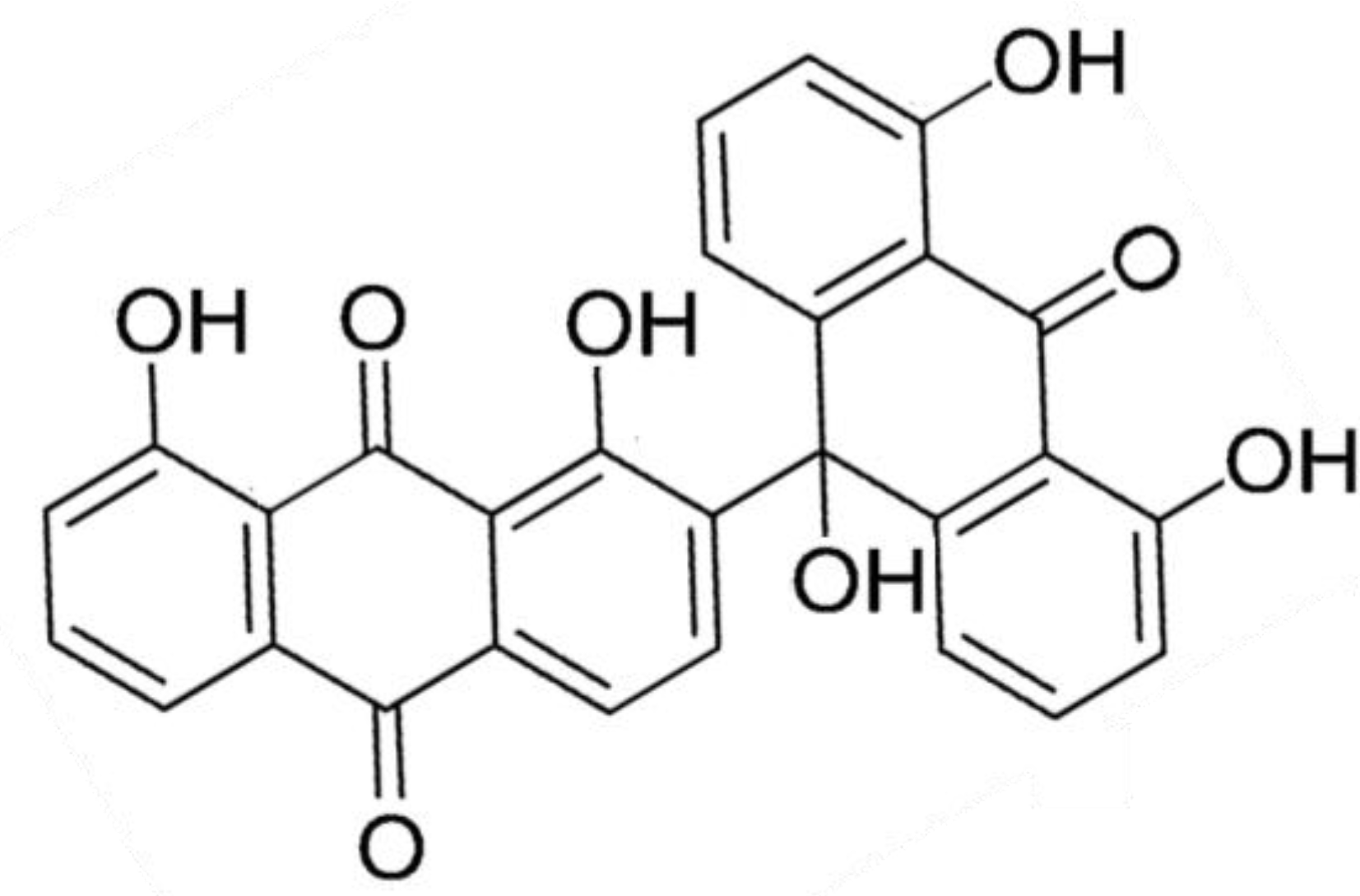
| Antimalarial | Albopunctatone (64) | Ascidian: Didemnum albopunctatum | Plasmodium falciparum Dd2, 3d7 strain inhibition | 4.4–5.3 μM (IC50) | [76] |
| Antiprotozoal | Viridamide A (65), B (66) | Bacterium: Oscillatoria nigro-viridis | Leishmania mexicana, Trypanosoma cruzi inhibition | 1.1–1.5 μM (IC50) | [77] |
| Antiprotozoal | Almiramides B (67), C (68) | Bacterium: Lyngbya majuscula | Leishmania donovani inhibition | 1.9–2.4 μM (IC50) | [78] |
| Antiprotozoal | Valinomycin (69) | Bacterium: Streptomyces sp. | Leishmania major & Trypanosoma brucei brucei inhibition | 0.0032–0.11 μM (IC50) | [79] |
| Antiprotozoal | Diketopiperazines (70–81) | Fungus: A. fumigatus, Nectria inventa | Trypanosoma brucei | 0.002–40 μM (IC50) | [80] |
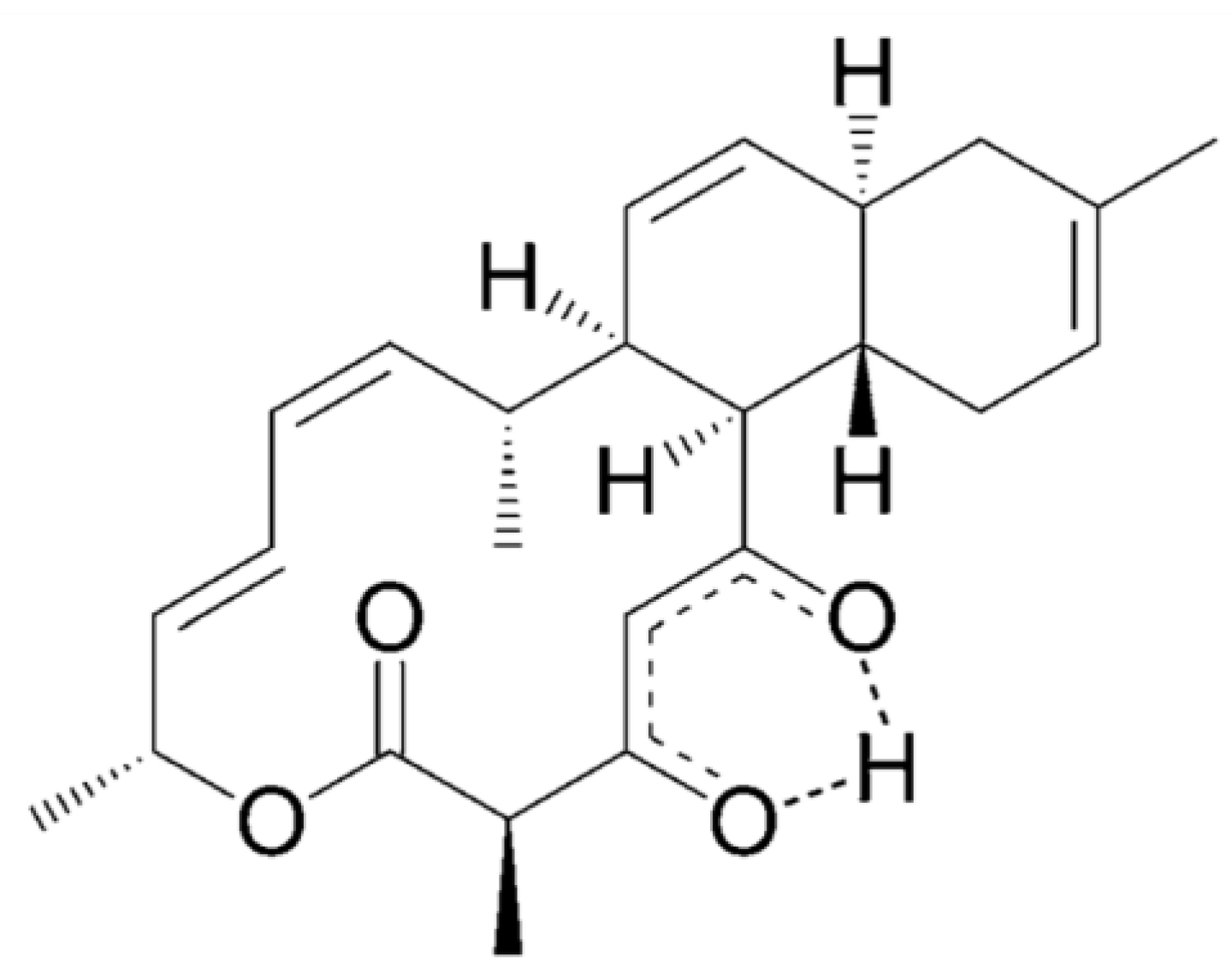
| Antituberculosis | Trichoderin A (82), A1 (83), B (84) | Fungus: Trichoderma sp. | Mycobacterium tuberculosis inhibition | 0.02–2 μg/mL (MIC) | [81] |
| Antiviral | Mirabamides A (85), C (86), D (87), E–H (88–91) | Sponge: Siliquariaspongia mirabilis | Anti-HIV-1 | 0.041–3.9 μM (IC50) | [82] |
| Antiviral | Mollamides B (92) | Tunicate: Didemmum molle | Anti-HIV | 48.7 μM (EC50) | [83] |
| Antiviral | Papuamide A (93) | Tunicate: Didemmum molle | Anti-HIV | 71 nM (EC50) | [84] |
| Antiviral | Celebesides A (94), C (95) | Sponge: Siliquariaspongia mirabilis | Anti-HIV-1 | 1.9 μg/mL (IC50) | [61] |
| Antiviral | Theopapuamide A (96), D (97) | Sponge: Theonella swinhoei | Anti-HIV-1 | 0.5 μM (IC50) | [61,85] |
| Antiviral | Asperterrestide A (98) | Fungus: Aspergillus terreus | Anti-HIN1, Anti-H3N2 | 0.41–20.2 μM (IC50) | [86] |
| Antiviral | Homophymine A–E (99–103),A1–E1 (104–108) | Sponge: Homophymia sp. | Anti-HIV-1 | 75 nM (IC50) | [87,88] |
| Antiviral | Koshikmaide B (109), F–H (110–112) | Sponge: Theonella sp. | Anti-HIV-1 | 2.3 μM (IC50) | [89,90] |
3.1. Antibacterial Activity
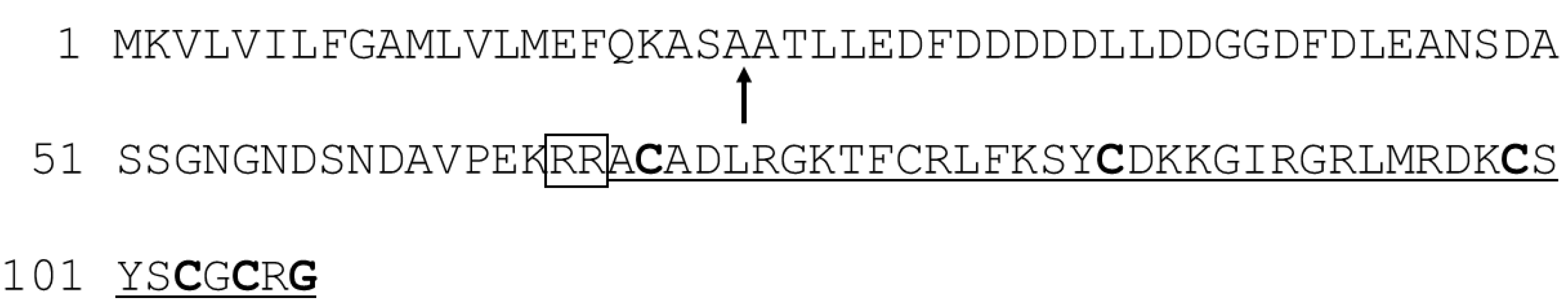
3.1.1. Aurelin

3.1.2. Arenicin-1

3.1.3. Tauramamide
3.1.4. Hepcidins

3.1.5. Scygonadin

3.1.6. Tunichromes
3.1.7. Bacillistatins 1 and 2
3.1.8. Thiopeptide TP-1161
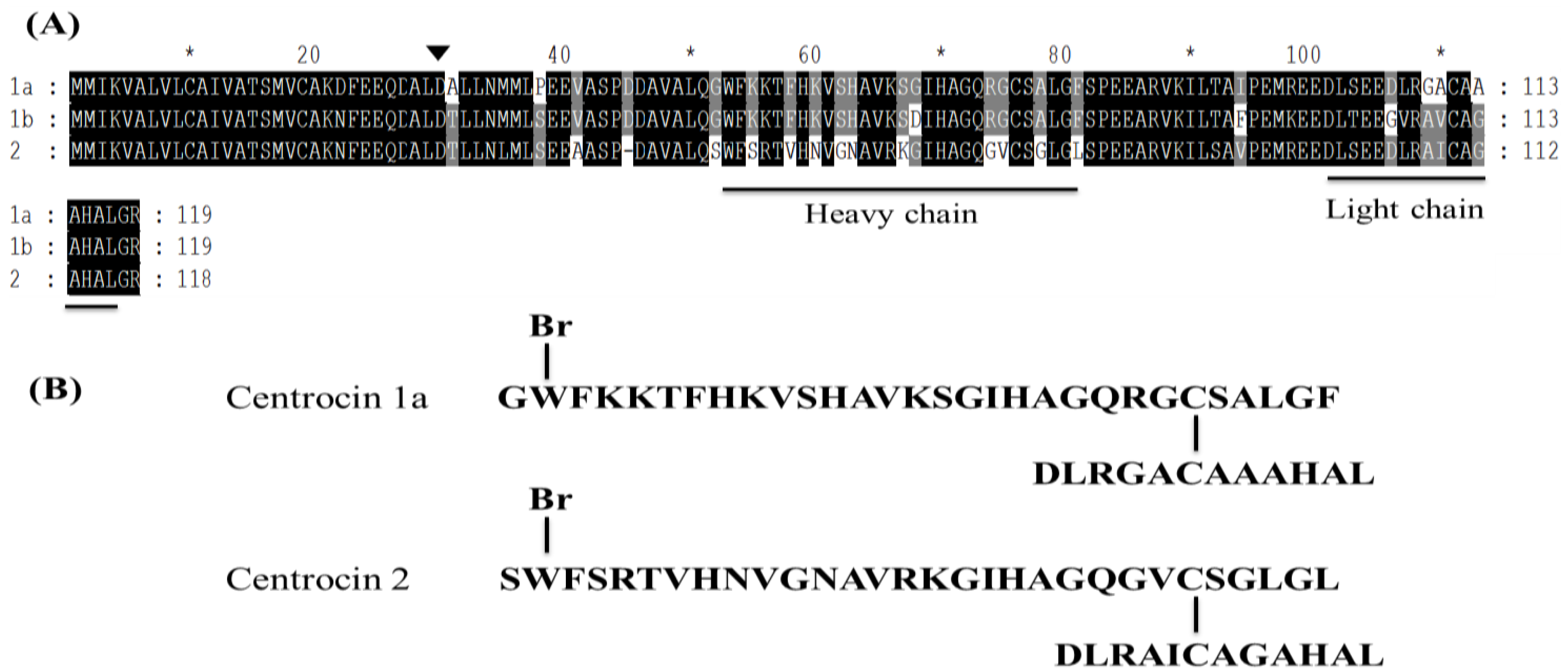
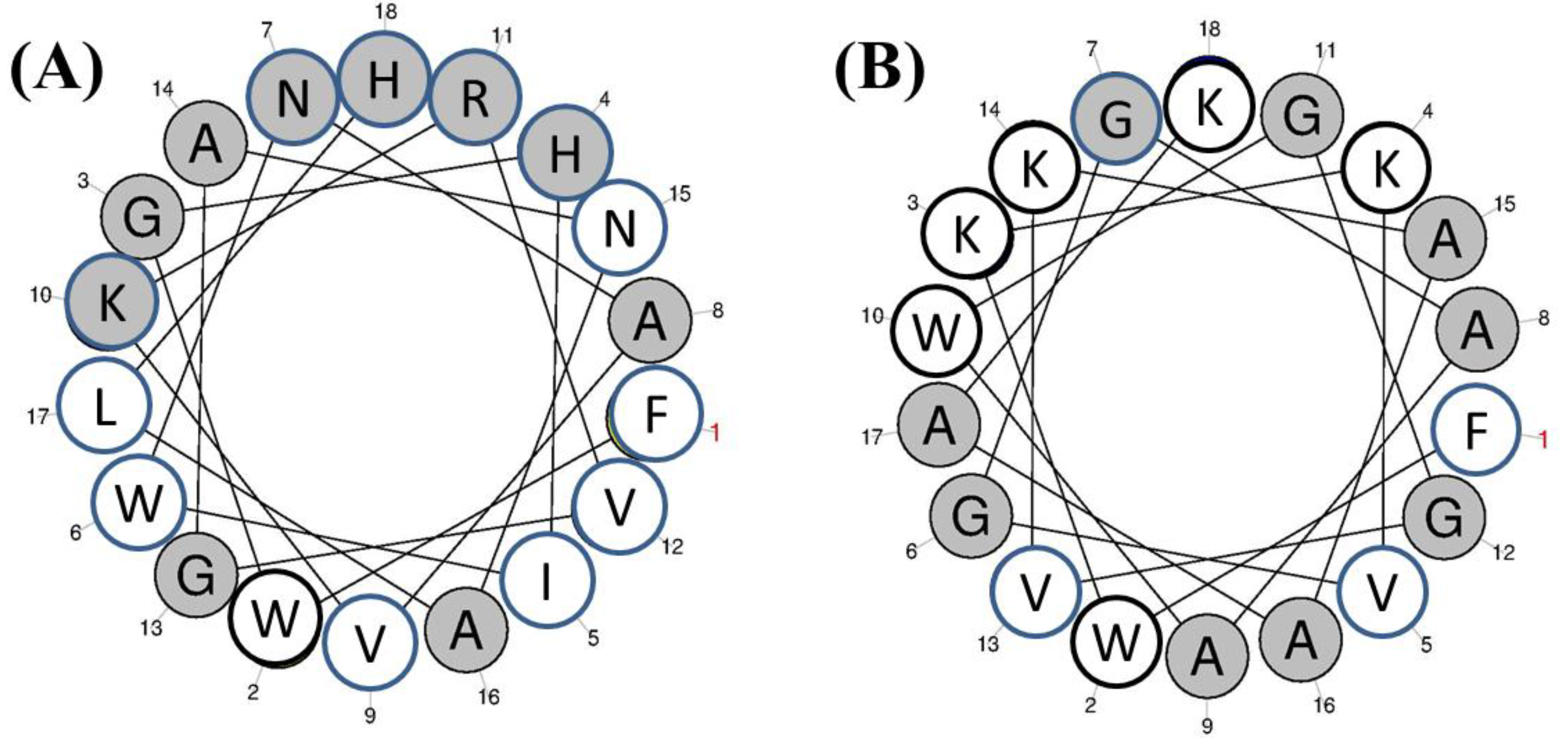
3.1.9. Centrocins
3.1.10. Halocyntin and Papillosin


3.1.11. Hyastatin

3.1.12. Indigoidine
3.1.13. Unnarmicins
3.1.14. Ngercheumicins
3.1.15. Solonamides
3.1.16. Cyclo-Peptides
3.1.17. Ariakemicins
3.1.18. Damicornin

3.1.19. Clavanins
3.1.20. Cadiolides

3.1.21. Cytosporones B and E

3.1.22. Anthracimycin

3.2. Antifungal Activity
3.2.1. Halocidin

3.2.2. Callipeltins J and K
3.2.3. Pedein A
3.2.4. Theonellamide
3.2.5. Theopapuamides
3.2.6. Surfactin

3.2.7. Anti-CA Cyclic Lipopeptide
3.2.8. Maribasins A and B
3.2.9. Mojavensin A
3.2.10. Kahalalides
3.2.11. Miuraenamides
3.2.12. Callyaerin
3.3. Anti-Malarial Activity
3.3.1. Dragomabin
3.3.2. Venturamides
3.3.3. Aerucyclamides
3.3.4. Gallinamide A
3.3.5. Lagunamide A and B
3.3.6. Albopunctatone
3.4. Antiprotozoal Activity

3.4.1. Viridamides
3.4.2. Almiramides
3.4.3. Valinomycin
3.4.4. Diketopiperazines

3.5. Antituberculosis Activity
Trichoderins
3.6. Anti-Viral Activity
3.6.1. Mirabamides A, C, and D
3.6.2. Mollamide B
3.6.3. Papuamides
3.6.4. Celebesides A and C
3.6.5. Theopapuamides
3.6.6. Asperterrestide A
3.6.7. Homophymine A
3.6.8. Koshikamide

{kind=link}
{kind=link}
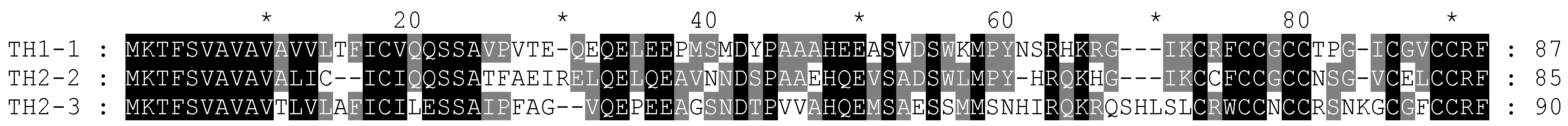{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
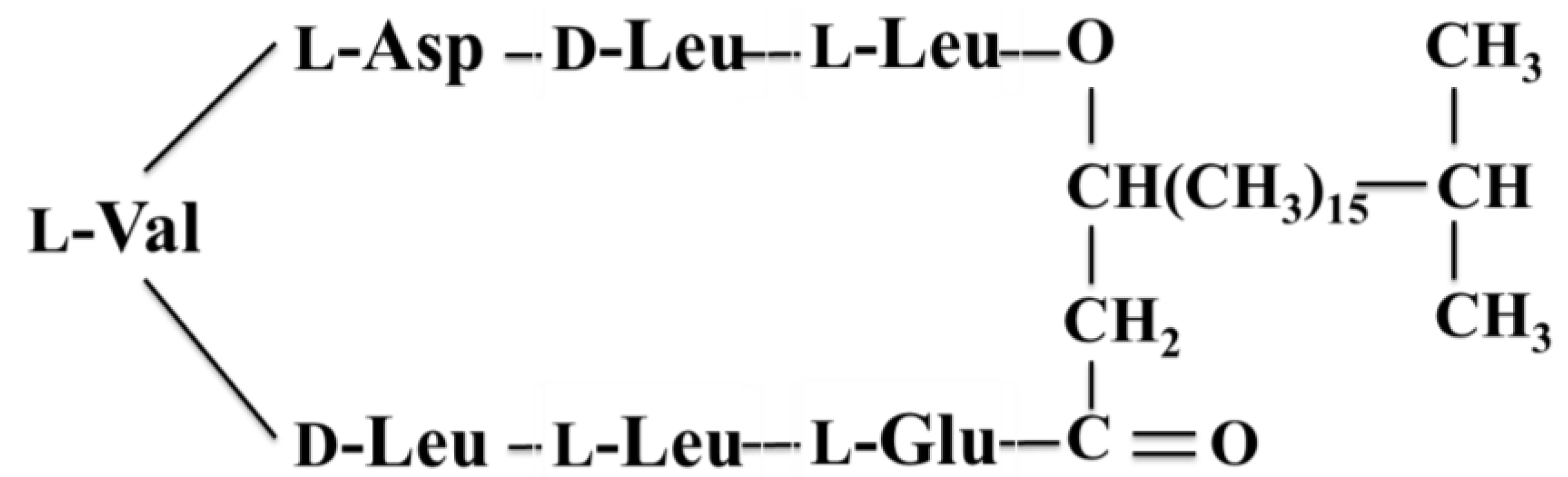{kind=link}
{kind=link}
{kind=link}
{kind=link}
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mayer, A.M.; Rodriguez, A.D.; Berlinck, R.G.; Fusetani, N. Marine pharmacology in 2009–2013: Marine compounds with antibacterial, antidiabetic, antifungal, anti-inflammatory, antiprotozoal, antituberculosis, and antiviral activities; affecting the immune and nervous systems, and other miscellaneous mechanisms of action. Mar Drugs 2013, 11, 2510–2573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masso-Silva, J.A.; Diamond, G. Antimicrobial Peptides from Fish. Pharmaceuticals 2014, 7, 265–310. [Google Scholar] [CrossRef] [PubMed]
- Winder, P.L.; Pomponi, S.A.; Wright, A.E. Natural Products from the Lithistida: A Review of the Literature since 2000. Mar. Drugs 2011, 9, 2643–2682. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M.; Rodriguez, A.D.; Berlinck, R.G.; Fusetani, N. Marine pharmacology in 2007–2008: Marine compounds with antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiprotozoal, antituberculosis and antiviral activities; affecting the immune and nervous system and other miscellaneous mechanism of action. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2011, 153, 191–222. [Google Scholar] [CrossRef] [PubMed]
- Villa, F.A.; Gernwick, L. Marine natural product drug discovery: Leads for treatment of inflammation, cancer, infections and neurological disorders. Immunopharmacol. Immunotoxicol. 2010, 32, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Munso, M.H.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2011, 28, 196–268. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Liu, J.; Yang, B.; Lin, X.; Yang, X.W.; Liu, Y. Marine natural products with anti-HIV activities in the last decade. Curr. Med. Chem. 2013, 20, 953–973. [Google Scholar] [PubMed]
- Kim, S.K.; Wijesekara, I. Development and biological activities of marine-derived bioactive peptides: A review. Curr. Protein Pept. Sci. 2010, 2, 1–9. [Google Scholar]
- Ngo, D.H.; Vo, T.S.; Ngo, D.N.; Wijesekara, I.; Kim, S.K. Biological activities and potential health benefits of bioactive peptides derived from marine organisms. Int. J. Biol. Macromol. 2012, 51, 378–383. [Google Scholar] [CrossRef] [PubMed]
- Laport, M.S.; Santos, D.C.; Muricy, G. Marine sponges: Potential sources of new antimicrobial drugs. Curr. Pharm. Biotechnol. 2009, 10, 86–105. [Google Scholar] [CrossRef] [PubMed]
- Sagar, S.; Kaur, M.; Minneman, K.P. Antiviral lead compounds from marine sponges. Mar. Drugs 2010, 8, 2619–2638. [Google Scholar] [CrossRef] [PubMed]
- Cardozo, K.H.M.; Guaratini, T.; Barros, M.P.; Falcao, V.R.; Tonon, A.P.; Lopes, N.P.; Campos, S.; Torres, M.A.; Souza, A.D.; Colepicolo, P.; et al. Metabolites from algae with economical impact. Comp. Biochem. Physiol. Part C 2007, 146, 60–78. [Google Scholar] [CrossRef]
- Bold, H.C.; Wynne, M.J. Introduction to the Algae Structure and Reproduction, 2nd ed.; Prentice-Hall Inc.: Englewood Cliffs, NJ, USA, 1985; pp. 1–33. [Google Scholar]
- Garson, J. Marine natural products. Nat. Prod. Rep. 1989, 6, 143–170. [Google Scholar] [CrossRef]
- Nair, R.; Chabbadiya, R.; Chanda, S. Marine algae: Screening for a potent antibacterial agent. Int. J. Herb. Pharmacother. 2007, 7, 73–86. [Google Scholar]
- Raja, A.; Vipin, C.; Aiyappan, A. Biological importance of Marine Algae-An overview. Int. J. Curr. Microbiol. Appl. Sci. 2013, 2, 222–227. [Google Scholar]
- Nunnery, J.K.; Mevers, E.; Gerwick, W.H. Biologically active secondary metabolites from marine cyanobacteria. Curr. Opin. Biotechnol. 2010, 21, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Cabrita, M.T.; Vale, C.; Router, A.P. Halogenated compounds from marine algae. Mar. Drugs 2010, 8, 2301–2317. [Google Scholar] [CrossRef] [PubMed]
- El Gamal, A.A. Biological importance of marine algae. Saudi. Pharm. J. 2010, 18, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Abad, M.J.; Bedoya, L.M.; Bermejo, P. Marine Compounds and their Antimicrobial Activities. In Science against Microbial Pathogens: Communicating Current Research and Technological Advances, 1st ed.; Mendez-Vilas, A., Ed.; FORMATEX: Badajoz, Spain, 2011; Volume 51, pp. 1293–1306. [Google Scholar]
- Ananya, A.K.; Iffat, Z.A. Cyanobacteria “the blue green algae” and its novel applications: A brief review. Int. J. Innov. Appl. Stud. 2014, 7, 251–261. [Google Scholar]
- Tan, L.T. Bioactive natural products from marine cyanobacteria for drug discovery. Phytochemistry 2007, 68, 954–979. [Google Scholar] [CrossRef] [PubMed]
- Jaiganesh, R.; Sampath Kumar, N.S. Marine bacterial sources of bioactive compounds. Adv. Food Nutr. Res. 2012, 65, 389–408. [Google Scholar] [PubMed]
- Debbab, A.; Aly, A.H.; Lin, W.H.; Proksch, P. Bioactive compounds from marine bacteria and fungi. Microb. Biotechnol. 2010, 3, 544–563. [Google Scholar] [CrossRef] [PubMed]
- Waters, A.L.; Hill, R.T.; Place, A.R.; Hamann, M.T. The expanding role of marine microbes in pharmaceutical development. Curr. Opin. Biotechnol. 2010, 21, 780–786. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.R.A.; Kaulekar, D.P.; Lokabarathi, P.A. Marine drugs from sponge-microbe association: A review. Mar. Drugs 2010, 8, 1417–1468. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Gao, X.; Liu, X.; Jiao, B. Marine microbes-derived anti-bacterial agents. Mini Rev. Med. Chem. 2010, 10, 1077–1090. [Google Scholar] [CrossRef] [PubMed]
- Rahman, H.; Austin, B.; Mitchell, W.J.; Morris, P.C.; Jamieson, A.J.; Adams, D.R.; Spragg, A.M.; Schweizer, M. Novel anti-infective compounds from marine bacteria. Mar. Drugs 2010, 8, 498–518. [Google Scholar] [CrossRef] [PubMed]
- Suarez-Jimenez, G.M.; Burgos-Hernandez, A.; Ezquerra-Brauer, J.M. Bioactive peptides and depsipeptides with anticancer potential: Sources from marine animals. Mar. Drugs 2012, 10, 963–986. [Google Scholar] [CrossRef] [PubMed]
- Brinkman, D.L.; Aziz, A.; Loukas, A.; Potriquet, J.; Seymour, J.; Mulvenna, J. Venom proteome of the box jellyfish Chironex fleckeri. PLoS One 2012, 7, e47866. [Google Scholar] [CrossRef] [PubMed]
- Ovchinnikova, T.V.; Balandin, S.V.; Aleshina, G.M.; Tagaev, A.A.; Leonova, Y.F.; Krasnodembsky, E.D.; Men’shenin, A.V.; Kokryakov, V.N. Aurelin, a novel antimicrobial peptide from jellyfish Aurelia aurita with structural features of defensins and channel-blocking toxins. Biochem. Biophys. Res. Commun. 2006, 348, 514–523. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.U.; Kang, D.I.; Zhu, W.L.; Shin, S.Y.; Hahm, K.S.; Kim, Y. Solution structures and biological functions of the antimicrobial peptide, arenicin-1, and its linear derivative. Biopolymers 2007, 88, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Balandin, S.V.; Zhmak, M.N.; Kudelina, I.A.; Finkina, E.I.; Kokryakov, V.N.; Arseniev, A.S. Recombinant expression, synthesis, purification, and solution structure of arenicin. Biochem. Biophys. Res. Commun. 2007, 360, 1561–1562. [Google Scholar]
- Desjardine, K.; Pereira, A.; Wright, H.; Matainaho, T.; Kelly, M.; Andersen, R.J. Tauramamide, a lipopeptide antibiotic produced in culture by Brevibacillus laterosporus isolated from a marine habitat: Structure elucidation and synthesis. J. Nat. Prod. 2007, 70, 1850–1853. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.H.; Chen, J.Y.; Kuo, C.M. Three different hepcidins from tilapia, Oreochromis mossambicus: Analysis of their expressions and biological functions. Mol. Immunol. 2007, 44, 1922–1934. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.S.; Wang, K.J.; Yang, M.; Cai, J.J.; Li, S.J.; Wang, G.Z. Purification and part characterization of a novel antibacterial protein scygonadin, isolated from the seminal plasma of mud crab, Scylla serrata (Forskal). J. Exp. Mar. Bio. Ecol. 2006, 339, 37–42. [Google Scholar] [CrossRef]
- Wang, K.J.; Huang, W.S.; Yang, M.; Chen, H.Y.; Bo, J.; Li, S.J.; Wang, G.Z. A malespecific expression gene, encodes a novel anionic antimicrobial peptide, scygonadin, in Scylla serrata. Mol. Immunol. 2007, 44, 1961–1968. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Sugumaran, M.; Robinson, W.E. The crosslinking and antimicrobial properties of tunichrome. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2008, 151, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Sugumaran, M.; Robinson, W.E. Structure, biosynthesis and possible function of tunichromes and related compounds. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2012, 163, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Pettit, G.R.; Knight, J.C.; Herald, D.L.; Pettit, R.K.; Hogan, F.; Mukku, V.J.; Hamblin, J.S.; Dodson, M.J.; Chapuis, J.C. Antineoplastic agents. Isolation and structure elucidation of bacillistatins 1 and 2 from a marine Bacillus silvestris. J. Nat. Prod. 2009, 72, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Engelhardt, K.; Degnes, K.F.; Kemmler, M.; Bredholt, H.; Fjaervik, E.; Klinkenberg, G.; Sletta, H.; Ellingsen, T.E.; Zotchev, S.B. Production of a new thiopeptide antibiotic, TP-1161, by a marine Nocardiopsis species. Appl. Environ. Microbiol. 2010, 76, 4969–4976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Haug, T.; Moe, M.K.; Styrvold, O.B.; Stensvag, K. Centrocins: Isolation and characterization of novel dimeric antimicrobial peptides from the green sea urchin, Strongylocentrotus droebachiensis. Dev. Comp. Immunol. 2010, 34, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Galinier, R.; Roger, E.; Sautiere, P.E.; Aumelas, A.; Banaigs, B.; Mitta, G. Halocyntin and papillosin, two new antimicrobial peptides isolated from hemocytes of the solitary tunicate, Halocynthia papillosa. J. Pept. Sci. 2009, 15, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Sperstad, S.V.; Haug, T.; Vasskog, T.; Stensvag, K. Hyastatin, a glycine-rich multi-domain antimicrobial peptide isolated from the spider crab (Hyas araneus) hemocytes. Mol. Immunol. 2009, 46, 2604–2612. [Google Scholar] [CrossRef] [PubMed]
- Slightom, R.N.; Buchan, A. Surface colonization by marine roseobacters: Integrating genotype and phenotype. Appl. Environ. Microbiol. 2009, 75, 6027–6037. [Google Scholar] [CrossRef] [PubMed]
- Oku, N.; Kawabata, K.; Adachi, K.; Katsuta, A.; Shizuri, Y. Unnarmicins A and C, new antibacterial depsipeptides produced by marine bacterium Photobacterium sp. MBIC06485. J. Antibiot. 2008, 61, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Adachi, K.; Kawabata, Y.; Kasai, H.; Katsuta, M.; Shizuri, Y. Novel Ngercheumicin or its salt useful for treating infection caused by Pseudovibrio denitrificans. Japanese Patent JP2007230911-A, 13 September 2007. [Google Scholar]
- Mansson, M.; Gram, L.; Larsen, T.O. Production of bioactive secondary metabolites by marine Vibrionaceae. Mar. Drugs 2011, 9, 1440–1468. [Google Scholar] [CrossRef] [PubMed]
- Rungprom, W.; Siwu, E.R.O.; Lambert, L.K.; Dechsakulwatana, C.; Barden, M.C.; Kokpol, U.; Blanchfield, J.T.; Kita, M.; Garson, M.J. Cyclic tetrapeptides from marine bacteria associated with the seaweed Diginea sp. and the sponge Halisarca ectofibrosa. Tetrahedron 2008, 64, 3147–3152. [Google Scholar] [CrossRef]
- Oku, N.; Adachi, K.; Matsuda, S.; Kasai, H.; Takatsaki, A.; Shizuri, Y. Ariakemicins A and B, novel polyketide-peptide antibiotics from a marine gliding bacterium of the genus Rapidithrix. Org. Lett. 2008, 10, 2481–2484. [Google Scholar] [CrossRef] [PubMed]
- Vidal-Dupiol, J.; Ladriere, O.; Destoumieux-Garzon, D.; Sautiere, P.E.; Meistertzheim, A.L.; Tambutte, E.; Tambutte, S.; Duval, D.; Foure, L.; Adjeroud, M.; et al. Innate immune responses of a scleractinian coral to vibriosis. J. Biol. Chem. 2011, 286, 22688–22698. [Google Scholar] [CrossRef] [PubMed]
- Saude, A.C.; Ombredane, A.S.; Silva, O.N.; Barbosa, J.A.; Moreno, S.E.; Guerra Araujo, A.C.; Falcão, R.; Silva, L.P.; Dias, S.C.; Franco, O.L. Clavanin bacterial sepsis control using a novel methacrylate nanocarrier. Int. J. Nanomed. 2014, 9, 5055–5069. [Google Scholar]
- Wang, W.; Kim, H.; Nam, S.J.; Rho, B.J.; Kang, H. Antibacterial butenolides from the Korean tunicate Pseudodistoma antinboja. J. Nat. Prod. 2012, 75, 2049–2054. [Google Scholar] [CrossRef] [PubMed]
- Beau, J.; Mahid, N.; Burda, W.N.; Harrington, L.; Shaw, L.N.; Mutka, T.; Kyle, D.E.; Barisic, B.; van Olphen, A.; Baker, B.J. Epigenetic tailoring for the production of anti-infective cytosporones from the marine fungus Leucostoma persoonii. Mar. Drugs 2012, 10, 762–774. [Google Scholar] [CrossRef] [PubMed]
- Jang, K.H.; Nam, S.J.; Locke, J.B.; Kauffman, C.A.; Beatty, D.S.; Paul, L.A.; Fenical, W. Anthracimycin, a potent anthrax antibiotic from a marine-derived actinomycete. Angew Chem. Int. Ed. Engl. 2013, 52, 7822–7824. [Google Scholar] [CrossRef] [PubMed]
- Jang, W.S.; Kim, H.K.; Lee, K.Y.; Kim, S.A.; Han, Y.S.; Lee, I.H. Antifungal activity of synthetic peptide derived from halocidin, antimicrobial peptide from the tunicate, Halocynthia aurantium. FEBS Lett. 2006, 580, 1490–1496. [Google Scholar] [CrossRef] [PubMed]
- D’Auria, M.V.; Sepe, V.; D’Orsi, R.; Bellotta, F.; Debitus, C.; Zampella, A. Isolation and structural elucidation of callipeltins J–M: Antifungal peptides from the marine sponge Latrunculia sp. Tetrahedron 2007, 63, 131–140. [Google Scholar] [CrossRef]
- Kunze, B.; Böhlendorf, B.; Reichenbach, H.; Höfle, G. Pedein A and B: Production, isolation, structure elucidation and biological properties of new antifungal cyclopeptides from Chondromyces pediculatus (Myxobacteria). J. Antibiot. 2008, 61, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, S.; Arita, Y.; Honda, M.; Iwamoto, K.; Matsuyama, A.; Shirai, A.; Kawasaki, H.; Kakeya, H.; Kobayashi, T.; Matsunaga, S.; et al. Marine antifungal theonellamides target 3β-hydroxysterol to activate rho1 signaling. Nat. Chem. Biol. 2010, 6, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Youssef, D.T.; Shaala, L.A.; Mohamed, G.A.; Badr, J.M.; Bamanie, F.H.; Ibrahim, S.R. Theonellamide G, a potent antifungal and cytotoxic bicyclic glycopeptide from the red sea marine sponge Theonella swinhoei. Mar. Drugs 2014, 12, 1911–1923. [Google Scholar] [CrossRef] [PubMed]
- Plaza, A.; Bifulco, G.; Keffer, J.L.; Lloyd, J.R.; Baker, H.L.; Bewley, C.A. Celebesides A–C and theopapuamides B–D, depsipeptides from an indonesian sponge that inhibit HIV-1 entry. J. Org. Chem. 2009, 74, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Berrue, F.; Ibrahim, A.; Boland, P.; Kerr, R.G. Newly isolated marine Bacillus pumilus (SP21): A source of novel lipoamides and other antimicrobial agents. Pure Appl. Chem. 2009, 81, 1027–1031. [Google Scholar] [CrossRef]
- Song, B.; Rong, Y.J.; Zhao, M.X.; Chi, Z.M. Antifungal activity of the lipopeptides produced by Bacillus amyloliquefaciens anti-CA against Candida albicans isolated from clinic. Appl. Microbiol. Biotechnol. 2013, 97, 7141–7150. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.J.; Liu, R.F.; Li, Y.G.; Tao, L.M.; Tian, L. Two new antifungal cyclic lipopeptides from Bacillus marinus B-9987. Chem. Pharm. Bull. (Tokyo) 2010, 58, 1630–1634. [Google Scholar] [CrossRef]
- Ma, Z.; Wang, N.; Hu, J.; Wang, S. Isolation and characterization of a new iturinic lipopeptide, mojavensin A produced by a marinederived bacterium Bacillus mojavensis B0621A. J. Antibiot. (Tokyo) 2012, 65, 317–322. [Google Scholar] [CrossRef]
- Gao, J.; Hamann, M.T. Chemistry and biology of kahalalides. Chem. Rev. 2011, 111, 3208–3235. [Google Scholar] [CrossRef] [PubMed]
- Iizuka, T.; Fudou, R.; Jojima, Y.; Ogawa, S.; Yamanaka, S.; Inukai, Y.; Ojika, M. Miuraenamides A and B, novel antimicrobial cyclic depsipeptides from a new slightly halophilic myxobacterium: Taxonomy, production, and biological properties. J. Antibiot. 2006, 59, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Ojika, M.; Inukai, Y.; Kito, Y.; Hirata, M.; Iizuka, T.; Fudou, R. Miuraenamides: Antimicrobial cyclic depsipeptides isolated from a rare and slightly halophilic myxobacterium. Chem. Asian J. 2008, 3, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.; Min, C.C.; Teuscher, F.; Ebel, R.; Kakoschke, C.; Lin, W.; Wray, V.; Edrada-Ebel, R.; Proksch, P. Callyaerins A–F and H, new cytotoxic cyclic peptides from the Indionesian marine sponge Callyspongia aerizusa. Bioorg. Med. Chem. 2010, 18, 4947–4956. [Google Scholar] [CrossRef] [PubMed]
- McPhail, K.L.; Correa, J.; Linington, R.G.; Gonzalez, J.; Ortega-Barria, E.; Capson, T.L.; Gerwick, W.H. Antimalarial linear lipopeptides from a Panamanian strain of the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2007, 70, 984–988. [Google Scholar] [CrossRef] [PubMed]
- Linington, R.G.; Gonzalez, J.; Urena, L.D.; Romero, L.I.; Ortega-Barria, E.; Gerwick, W.H. Venturamides A and B: Antimalarial constituents of the Panamanian marine cyanobacterium Oscillatoria sp. J. Nat. Prod. 2007, 70, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Portmann, C.; Blom, J.F.; Gademann, K.; Juttner, F. Aerucyclamides A and B: Isolation and synthesis of toxic ribosomal heterocyclic Peptides from the Cyanobacterium Microcystis aeruginosa PCC 7806. J. Nat. Prod. 2008, 71, 1193–1196. [Google Scholar] [CrossRef] [PubMed]
- Ziemert, N.; Ishida, K.; Quillardet, P.; Bouchier, C.; Hertweck, C.; de Marsac, N.T.; Dittmann, E. Microcyclamide biosynthesis in two strains of Microcystis aeruginosa: From structure to genes and Vice Versa. Appl. Environ. Microbiol. 2008, 74, 1791–1797. [Google Scholar] [CrossRef] [PubMed]
- Linington, R.G.; Clark, B.R.; Trimble, E.E.; Almanza, A.; Urena, L.D.; Kyle, D.E.; Gerwick, W.H. Antimalarial peptides from marine cyanobacteria: Isolation and structural elucidation of gallinamide A. J. Nat. Prod. 2009, 72, 14–17. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.; Puddick, J.; Prinsep, M.R.; Rottmann, M.; Tan, L.T. Lagunamides A and B: Cytotoxic and antimalarial cyclodepsipeptides from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2010, 73, 1810–1814. [Google Scholar] [CrossRef] [PubMed]
- Carroll, A.R.; Nash, B.D.; Duffy, S.; Avery, V.M. Albopunctatone, an antiplasmodial anthrone-anthraquinone from the Australian ascidian Didemnum albopunctatum. J. Nat. Prod. 2012, 75, 1206–1209. [Google Scholar] [CrossRef] [PubMed]
- Simmons, T.L.; Engene, N.; Urena, L.D.; Romero, L.I.; Ortega-Barria, E.; Gerwick, L.; Gerwick, W.H. Viridamides A and B, lipodepsipeptides with antiprotozoal activity from the marine cyanobacterium Oscillatoria nigro-viridis. J. Nat. Prod. 2008, 71, 1544–1550. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, L.M.; Lopez, D.; Vesely, B.A.; Della Togna, G.; Gerwick, W.H.; Kyle, D.E.; Linington, R.G. Almiramides A–C: Discovery and Development of a New Class of Leishmaniasis Lead Compounds. J. Med. Chem. 2010, 53, 4187–4197. [Google Scholar] [CrossRef] [PubMed]
- Pimentel-Elardo, S.M.; Kozytska, S.; Bugni, T.S.; Ireland, C.M.; Moll, H.; Hentschel, U. Anti-parasitic compounds from Streptomyces sp. strains isolated from mediterranean sponges. Mar. Drugs 2010, 8, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Watts, K.R.; Ratnam, J.; Ang, K.H.; Tenney, K.; Compton, J.E.; McKerrow, J.; Crews, P. Assessing the trypanocidal potential of natural and semi-synthetic diketopiperazines from two deep water marine-derived fungi. Bioorg. Med. Chem. 2010, 18, 2566–2574. [Google Scholar] [CrossRef] [PubMed]
- Pruksakorn, P.; Arai, M.; Kotoku, N.; Vilcheze, C.; Baughn, A.D.; Moodley, P.; Jacobs, W.R., Jr.; Kobayashi, M. Trichoderins, novel aminolipopeptides from a marine sponge-derived Trichoderma sp., are active against dormant mycobacteria. Bioorg. Med. Chem. Lett. 2010, 20, 3658–3663. [Google Scholar] [CrossRef] [PubMed]
- Plaza, A.; Gustchina, E.; Baker, H.L.; Kelly, M.; Bewley, C.A. Mirabamides A–D, depsipeptides from the sponge Siliquariaspongia mirabilis that inhibit HIV-1 fusion. J. Nat. Prod. 2007, 70, 1753–1760. [Google Scholar] [CrossRef] [PubMed]
- Donia, M.S.; Wang, B.; Dunbar, D.C.; Desai, P.V.; Patny, A.; Avery, M.; Hamann, M.T. Mollamides B and C, cyclic hexapeptides from the Indonesian tunicate Didemnum molle. J. Nat. Prod. 2008, 71, 941–945. [Google Scholar] [CrossRef] [PubMed]
- Andjelic, C.; Planelles, V.; Barrows, L. Characterizing the anti-HIV activity of papuamide A. Mar. Drugs 2008, 6, 528–549. [Google Scholar] [CrossRef] [PubMed]
- Ratnayake, A.S.; Bugni, T.S.; Feng, X.; Harper, M.K.; Skalicky, J.J.; Mohammed, K.A.; Andjelic, C.D.; Barrows, L.R.; Ireland, C.M. Theopapuamide, a cyclic depsipeptide from a Papua New Guinea Lithistid sponge Theonella swinhoei. J. Nat. Prod. 2006, 69, 1582–1586. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Bao, J.; Zhang, X.Y.; Tu, Z.C.; Shi, Y.M.; Qi, S.H. Asperterrestide A, a cytotoxic cyclic tetrapeptide from the marine-derived fungus Aspergillus terreus SCSGAF0162. J. Nat. Prod. 2013, 76, 1182–1186. [Google Scholar] [CrossRef] [PubMed]
- Zampella, A.; Sepe, V.; Bellotta, F.; Luciano, P.; D’Auria, M.V.; Cresteil, T.; Debitus, C.; Petek, S.; Poupat, C.; Ahond, A. Homophymines B–E and A1–E1, a family of bioactive cyclodepsipeptides from the sponge Homophymia sp. Org. Biomol. Chem. 2009, 7, 4037–4044. [Google Scholar] [CrossRef] [PubMed]
- Zampella, A.; Sepe, V.; Luciano, P.; Bellotta, F.; Monti, M.C.; D’Auria, M.V.; Jepsen, T.; Petek, S.; Adeline, M.-T.; Laprevote, O.; et al. Homophymine A, an anti-HIV cyclodepsipeptide from the sponge Homophymia sp. J. Org. Chem. 2008, 73, 5319–5327. [Google Scholar] [CrossRef] [PubMed]
- Araki, T.; Matsunaga, S.; Nakao, Y.; Furihata, K.; West, L.; Faulkner, D.J.; Fusetani, N. Koshikamide B, a cytotoxic peptide lactone from a marine sponge Theonella sp. J. Org. Chem. 2008, 73, 7889–7894. [Google Scholar] [CrossRef] [PubMed]
- Plaza, A.; Bifulco, G.; Masullo, M.; Lloyd, J.R.; Keffer, J.L.; Colin, P.L.; Hooper, J.N.A.; Bell, L.J.; Bewley, C.A. Mutremdamide A and koshikamides C–H, peptide inhibitors of HIV-1 entry from different Thennella species. J. Org. Chem. 2010, 75, 4344–4355. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, J. Cationic antimicrobial peptides: Issues for potential clinical use. BioDrugs 2003, 17, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Ovchinnikova, T.V.; Shenkarev, Z.O.; Balandin, S.V.; Nadezhdin, K.D.; Paramonov, A.S.; Kokryakov, V.N.; Arseniev, A.S. Molecular insight into mechanism of antimicrobial action of the beta-hairpin peptide arenicin: Specific oligomerization in detergent micelles. Biopolymers 2008, 89, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Yederi, R.; Reddy, K. Purification and characterization of antibacterial proteins from granular hemocytes of Indian mud crab, Scylla serrata. Acta Biochim. Pol. 2009, 56, 71–82. [Google Scholar] [PubMed]
- Cude, W.N.; Mooney, J.; Tavanaei, A.A.; Hadden, M.K.; Frank, A.M.; Gulvik, C.A.; May, A.L.; Buchan, A. Production of the antimicrobial secondary metabolite indigoidine contributes to competitive surface colonization by the marine roseobacter Phaeobacter sp. strain Y4I. Appl. Environ. Microbiol. 2012, 78, 4771–4780. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, R.; Gautam, H. Toward the synthesis and biological screening of a cyclotetrapeptide from marine bacteria. Mar. Drugs 2011, 9, 71–81. [Google Scholar] [CrossRef]
- Binz, T.M.; Maffioli, S.I.; Sosio, M.; Donadio, S.; Muller, R. Insights into an unusual nonribosomal peptide synthetase biosynthesis. J. Biol. Chem. 2010, 285, 32710–32719. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, R. Cyclopolypeptides with antifungal interest. Coll. Pharm. Commun. 2013, 1, 1–15. [Google Scholar]
- Diehl, K.B. Topical Antifungal Agents: An Update. Am. Fam. Phys. 1996, 54, 1687–1692. [Google Scholar]
- Onishi, J.; Meinz, M.; Thompson, J.; Curotto, J.; Dreikorn, S.; Rosenbach, M.; Douglas, C.; Abruzzo, G.; Tsipouras, A.; Wilson, K.; et al. Discovery of novel antifungal (1,3)-beta-d-glucan synthase inhibitors. Antimicrob. Agents Chemother. 2000, 44, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Jang, W.S.; Kim, C.H.; Kang, M.S.; Chae, H.J.; Son, S.M.; Seo, S.J.; Lee, I.H. cDNA cloning of halocidin and a new antimicrobial peptide derived from the N-terminus of Ci-META4. Peptides 2005, 26, 2360–2367. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Ren, B.; Gao, H.; Liu, M.; Dai, H.; Song, F.; Yu, Z.; Wang, S.; Hu, J.; Kokare, C.R.; Zhang, L. Optimization for the production of surfactin with a new synergistic antifungal activity. PLoS One 2012, 7, e34430. [Google Scholar] [CrossRef] [PubMed]
- Shilabin, A.G.; Hamann, M.T. In vitro and in vivo evaluation of select kahalalide F analogs with antitumor and antifungal activities. Bioorg. Med. Chem. 2011, 19, 6628–6632. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Available online: http://www.who.int/neglected_diseases/en/ (accessed on 14 October 2014).
- Penet, M.F.; Kober, F.; Confort-Gouny, S.; Le Fur, Y.; Dalmasso, C.; Coltel, N.; Liprandi, A.; Gulian, J.M.; Grau, G.E.; Cozzone, P.J.; et al. Magnetic resonance spectroscopy reveals an impaired brain metabolic profile in mice resistant to cerebral malaria infected with Plasmodium berghei ANKA. J. Biol. Chem. 2007, 282, 14505–14514. [Google Scholar]
- Snow, R.W.; Guerra, C.A.; Noor, A.M.; Myint, H.Y.; Hay, S.I. The global distribution of clinical episodes of Plasmodium falciparum malaria. Nature 2005, 434, 214–217. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.K.; Satpathy, S.K.; Mohanty, S. Survey of malaria treatment and deaths. Bull. World Health Organ. 1999, 77, 1020. [Google Scholar] [PubMed]
- Plebanski, M.; Locke, E.; Kazura, J.W.; Coppel, R.L. Malaria vaccines: Into a mirror, darkly? Trends Parasitol. 2008, 24, 532–536. [Google Scholar] [CrossRef] [PubMed]
- Schlitzer, M. Malaria chemotherapeutics part I: History of antimalarial drug development, currently used therapeutics, and drugs in clinical development. ChemMedChem 2007, 2, 944–986. [Google Scholar] [CrossRef] [PubMed]
- Schlitzer, M. Antimalarial drugs—what is in use and what is in the pipeline. Archiv. Der Pharmazie 2008, 341, 149–163. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Montero, A.; Gastaminza, P.; Whitten-Bauer, C.; Wieland, S.F.; Isogawa, M.; Fredericksen, B.; Selvarajah, S.; Gallay, P.A.; Ghadiri, M.R.; et al. A virocidal amphipathic alpha-helical peptide that inhibits hepatitis C virus infection in vitro. Proc. Natl. Acad. Sci. USA 2008, 105, 3088–3093. [Google Scholar] [CrossRef] [PubMed]
- Bai, F.; Town, T.; Pradhan, D.; Cox, J.; Ashish; Ledizet, M.; Anderson, J.F.; Flavell, R.A.; Krueger, J.K.; Koski, R.A.; et al. Antiviral peptides targeting the West Nile virus envelope protein. J. Virol. 2007, 81, 2047–2055. [Google Scholar] [CrossRef] [PubMed]
- Budge, P.J.; Graham, B.S. Inhibition of respiratory syncytial virus by RhoA-derived peptides: Implications for the development of improved antiviral agents targeting heparin-binding viruses. J. Antimicrob. Chemother. 2004, 54, 299–302. [Google Scholar] [CrossRef] [PubMed]
- Cooper, D.A.; Lange, J.M. Peptide inhibitors of virus-cell fusion: Enfuvirtide as a case study in clinical discovery and development. Lancet Infect. Dis. 2004, 4, 426–436. [Google Scholar] [CrossRef] [PubMed]
- Altmann, S.E.; Jones, J.C.; Schultz-Cherry, S.; Brandt, C.R. Inhibition of vaccinia virus entry by a broad spectrum antiviral peptide. Virology 2009, 388, 248–259. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; van Wagoner, R.M.; Harper, M.K.; Baker, H.L.; Hooper, J.N.A.; Bewley, C.A.; Ireland, C.M. Mirabamides E–H, HIV-inhibitory depsipeptides from the sponge Stelletta clavosa. J. Nat. Prod. 2011, 74, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Elyakova, L.A.; Vaskovsky, B.V.; Khoroshilova, N.I.; Vantseva, S.I.; Agapkina, Y.Y. Isolation and structure of a novel peptide inhibitor of HIV-1 integrase from marine polychaetes. Russ. J. Bioorg. Chem. 2011, 37, 207–216. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, H.K.; Seo, C.H.; Park, Y. Marine Peptides and Their Anti-Infective Activities. Mar. Drugs 2015, 13, 618-654. https://doi.org/10.3390/md13010618
Kang HK, Seo CH, Park Y. Marine Peptides and Their Anti-Infective Activities. Marine Drugs. 2015; 13(1):618-654. https://doi.org/10.3390/md13010618
Chicago/Turabian StyleKang, Hee Kyoung, Chang Ho Seo, and Yoonkyung Park. 2015. "Marine Peptides and Their Anti-Infective Activities" Marine Drugs 13, no. 1: 618-654. https://doi.org/10.3390/md13010618
APA StyleKang, H. K., Seo, C. H., & Park, Y. (2015). Marine Peptides and Their Anti-Infective Activities. Marine Drugs, 13(1), 618-654. https://doi.org/10.3390/md13010618