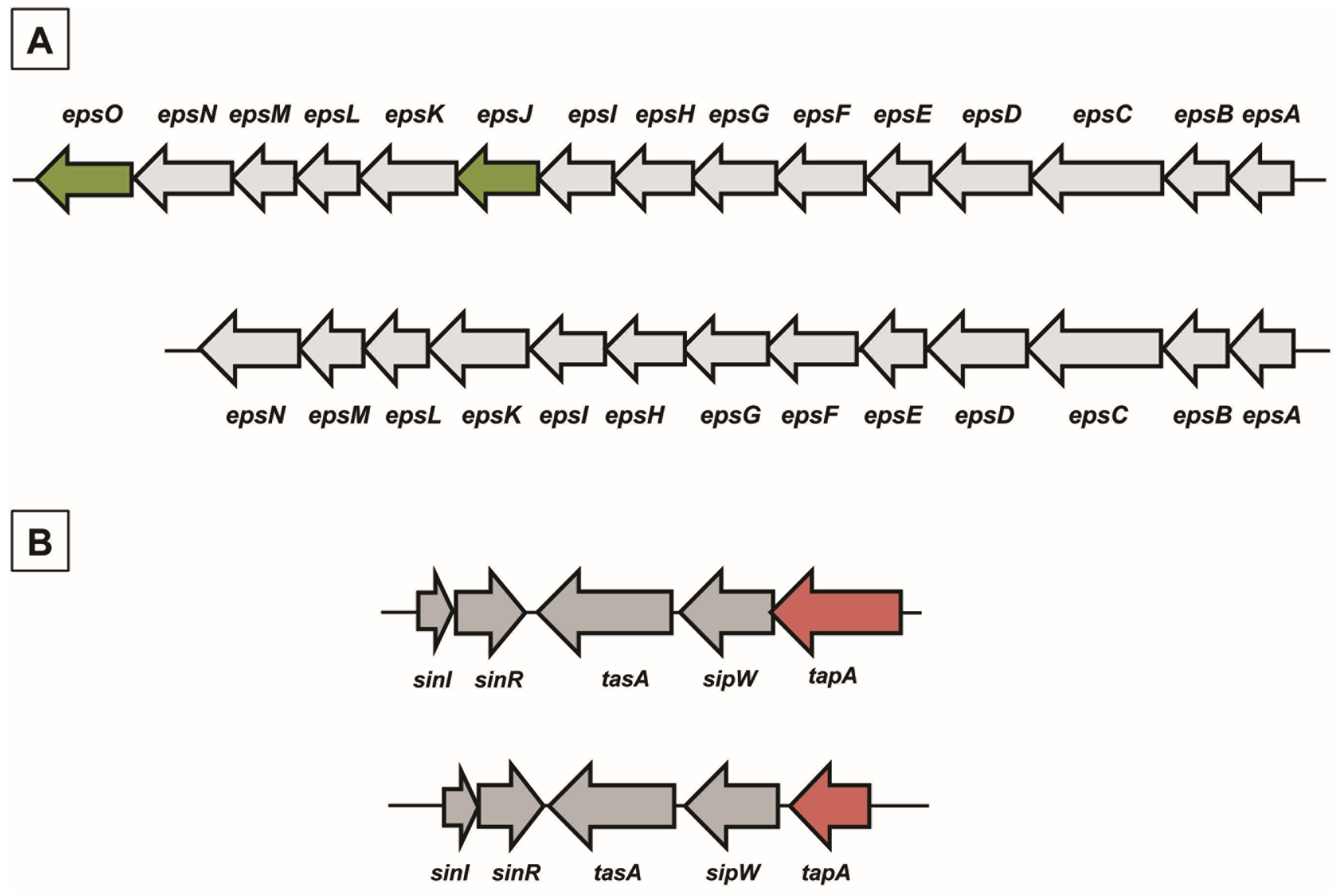
2.1. Matrix Genes in B. pumilus ATCC 7061T and SF214
In
B. subtilis the main matrix genes are clustered in the
epsA-O and
tapA-sipW-tasA operons [
10]. We analyzed the genomes of the
B. pumilus reference strain, ATCC 7061
T (PRJNA29785), and of strain SF214 (PRJNA290581) and found homologs of most biofilm genes of
B. subtilis (
Table 1). Homologs were also present in other
B. pumilus strains whose genomes are available on the NCBI databank (CCTCCM205165; SAFR032; S-1; CCMA-560; INR7; BA06). In all cases the similarities with proteins of the strain ATCC 7061
T were between 95% and 100% (data not shown). With the exception of the
epsJ and
epsO genes
, not present in either strain of
B. pumilus, all other genes of the
epsA–O (
Figure 2A) and
tapA-sipW-tasA (
Figure 2B) operons were present and organized as in
B. subtilis. The function of the products of the
epsJ and
epsO genes of
B. subtilis is not known in detail. It has only been reported that mutations in the
epsJ gene display a defect in pellicle formation and swarming [
19]. The
B. pumilus strains here considered, both lacking
epsJ, form a pellicle similar to that of the wild-type strain of
B. subtilis (not shown)
. Therefore, the function of the
epsJ product is still elusive. The gene synteny was similar between the two species, with the
tapA-sipW-tasA operon adjacent but divergently oriented with respect to the
sinI–sinR genes, also involved in matrix synthesis (
Figure 2B).
Figure 2.
Chromosomal organization of genes involved in matrix synthesis in B. subtilis and B. pumilus. (A) The epsA–O operon in B. subtilis (top) and B. pumilus (bottom). In dark green are indicated two genes present in B. subtilis and lacking in B. pumilus. (B) Chromosomal organization of the sinI–sinR gene pair and the tapA-sipW-tasA operon in B. subtilis (top) and B. pumilus (bottom). In red is the tapA gene, significantly smaller in B. pumilus than in B. subtilis.
Figure 2.
Chromosomal organization of genes involved in matrix synthesis in B. subtilis and B. pumilus. (A) The epsA–O operon in B. subtilis (top) and B. pumilus (bottom). In dark green are indicated two genes present in B. subtilis and lacking in B. pumilus. (B) Chromosomal organization of the sinI–sinR gene pair and the tapA-sipW-tasA operon in B. subtilis (top) and B. pumilus (bottom). In red is the tapA gene, significantly smaller in B. pumilus than in B. subtilis.
Table 1.
Genes and putative proteins involved in matrix synthesis in B. subtilis 168, B. pumilus ATCC 7061T, and B. pumilus SF214.
Table 1.
Genes and putative proteins involved in matrix synthesis in B. subtilis 168, B. pumilus ATCC 7061T, and B. pumilus SF214.
| Genes | Putative Encoded Protein | Protein Identity (%) |
|---|
| B. subtilis 168 vs. B. pumilus ATCC 7061T | B. pumilus ATCC 7061T vs. B. pumilus SF214 |
|---|
| Eps Operon |
| epsO | pyruvil transferase | - | - |
| epsN | pyridoxal phosphate-dependent aminotransferase | 68 | 97 |
| epsM | acetyl transferase | 50 | 94 |
| epsL | sugar transferase | 65 | 97 |
| epsK | membrane protein | 50 | 96 |
| epsJ | glycosyl transferase | - | - |
| epsI | pyruvil transferase | 60 | 95 |
| epsH | glycosyl transferase | 46 | 96 |
| epsG | membrane protein | 72 | 96 |
| epsF | glycosyl transferase | 54 | 95 |
| epsE | glycosyl transferase | 61 | 96 |
| epsD | glycosyl transferase | 54 | 94 |
| epsC | polysaccharide biosynthesis protein | 62 | 99 |
| epsB | tyrosine-protein kinase | 63 | 99 |
| epsA | capsular biosynthesis protein | 43 | 100 |
| TasA Operon |
| tapA | lipoprotein for formation | 42 | 93 |
| sipW | signal peptidase I | 52 | 98 |
| tasA | spore coat protein N | 62 | 97 |
The identity of the predicted products of the
epsA–O and
tapA-sipW-tasA operons varied from 42% to 68% between
B. subtilis and
B. pumilus, while proteins of the two
B. pumilus strains were almost identical (
Table 1). TapA, the product of the promoter proximal gene of the
tapA-sipW-tasA operon, was the least conserved of the biofilm proteins analyzed here. In
B. subtilis, TapA is required to anchor TasA amyloid fibers to the cell wall [
20] and its prominent features are an N-terminal signal secretion sequence, five conserved cysteine residues and an eight amino acid sequence at the N-terminal part of the protein (region 50–57), involved in TasA fiber formation
in vitro [
20]. In both
B. pumilus strains TapA is shorter than in
B. subtilis (175
vs. 253 amino acids), and the differences were mainly found in the C- and N-terminal regions of the proteins (see
Supplementary Materials Figure S1). The 60 C-terminal residues of the
B. subtilis protein, totally lacking in TapA of
B. pumilus (see
Supplementary Materials Figure S1), are homologous to the SPAM (Secreted Polymorphic Antigen-associated with Merozoites) domains of
Plasmodium falciparum [
21,
22] and are not required for TasA fiber formation
in vitro [
20]. At the N-terminal all TapA proteins have signal secretion sequences. Those of
B. pumilus are not similar to that of
B. subtilis (see
Supplementary Materials Figure S1) but were recognized as signal secretion sequences with a predicted cleavage site at position 30 (see
Supplementary Materials Figure S2). TapA of
B. pumilus has the five cysteine residues conserved in other
Bacillus species [
20], but one of the cysteine residues is within the signal secretion sequence (see
Supplementary Materials Figures S1 and S2). The eight amino acids region (50–57) essential for TasA fiber formation
in vitro in
B. subtilis [
20] is not conserved in
B. pumilus (
Supplementary Materials Figure S1), suggesting either a different mechanism of TapA-mediated anchoring of TasA fiber to the cell wall or a different role of TapA in matrix assembly. The other two products of the
tapA-sipW-tasA operon, TasA and SipW, are, however, similar in size and primary structure to the two former spore species (see
Supplementary Materials Figure S3).
A previous study has reported that the overall similarity of the
eps operons of
B. subtilis and
B. cereus is relatively low and that only the products of
epsC,
epsL, and
epsN genes are conserved [
14]. The same three genes are also among the most conserved matrix genes between
B. subtilis and
B. pumilus (
Table 1), pointing to an essential role of their products in matrix biosynthesis.
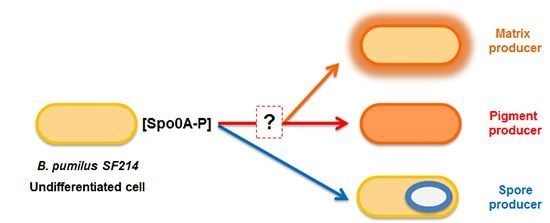
2.2. Physiological and Chemical Characterization of the Matrix of B. pumilus SF214
In liquid cultures of
B. pumilus SF214, matrix formation is an alternative to sporulation and in some cells parallel to pigment production [
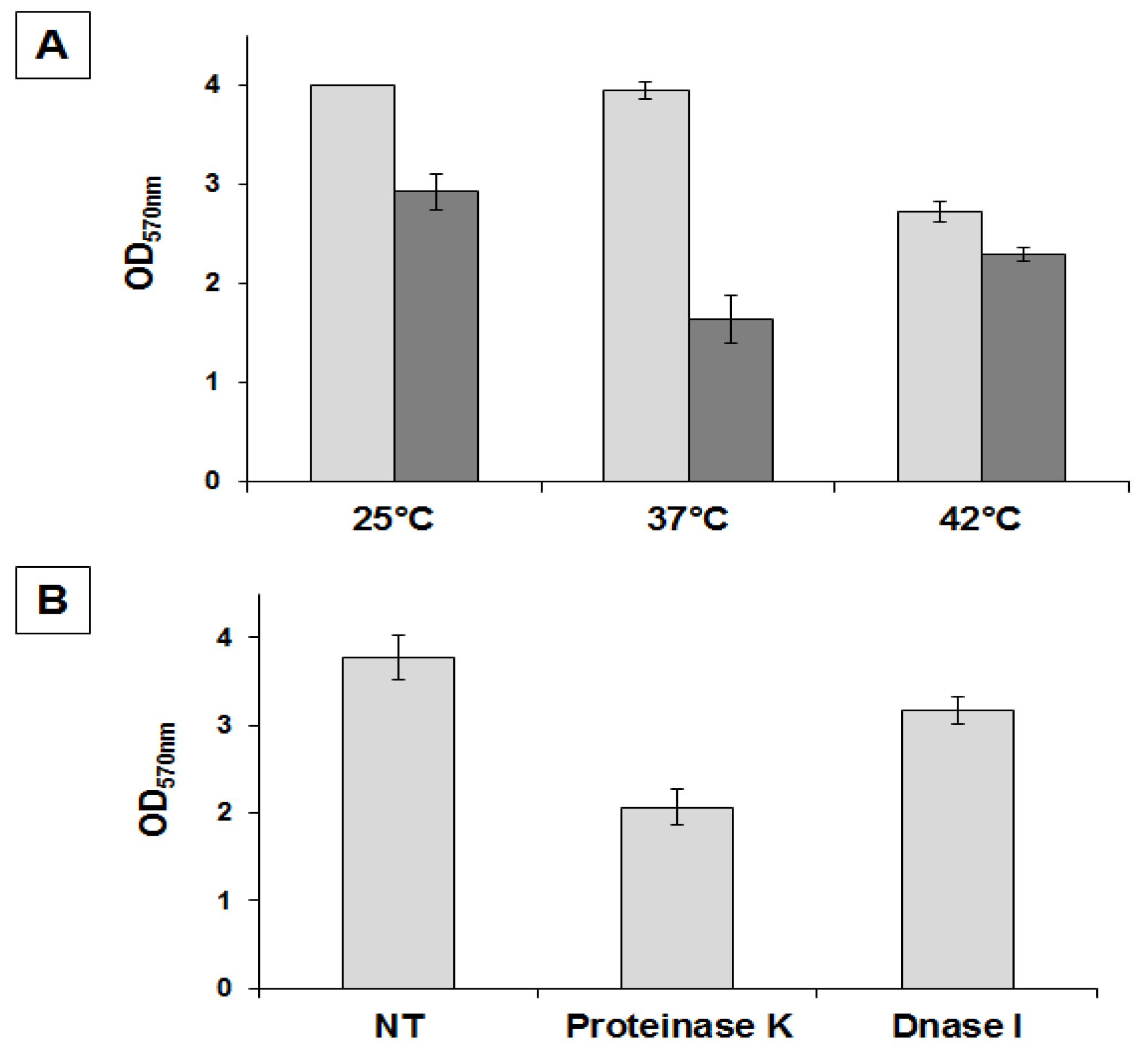
18]. In order to better characterize matrix production in strain SF214 we performed an EPS assay (see
Section 3.3) on cells grown at different temperatures and in different media. In a rich (LB) medium we observed the highest production of biofilm, similar at 25 and 37 °C and approximately 25% reduced at 42 °C (
Figure 3A). In Difco Sporulation (DS) medium we observed the highest production of biofilm at 25 °C, similar at 42 °C and reduced by approximately 30% at 37 °C (
Figure 3A). Cells grown in DS medium produced less biofilm than cells grown in LB medium at all tested temperatures (
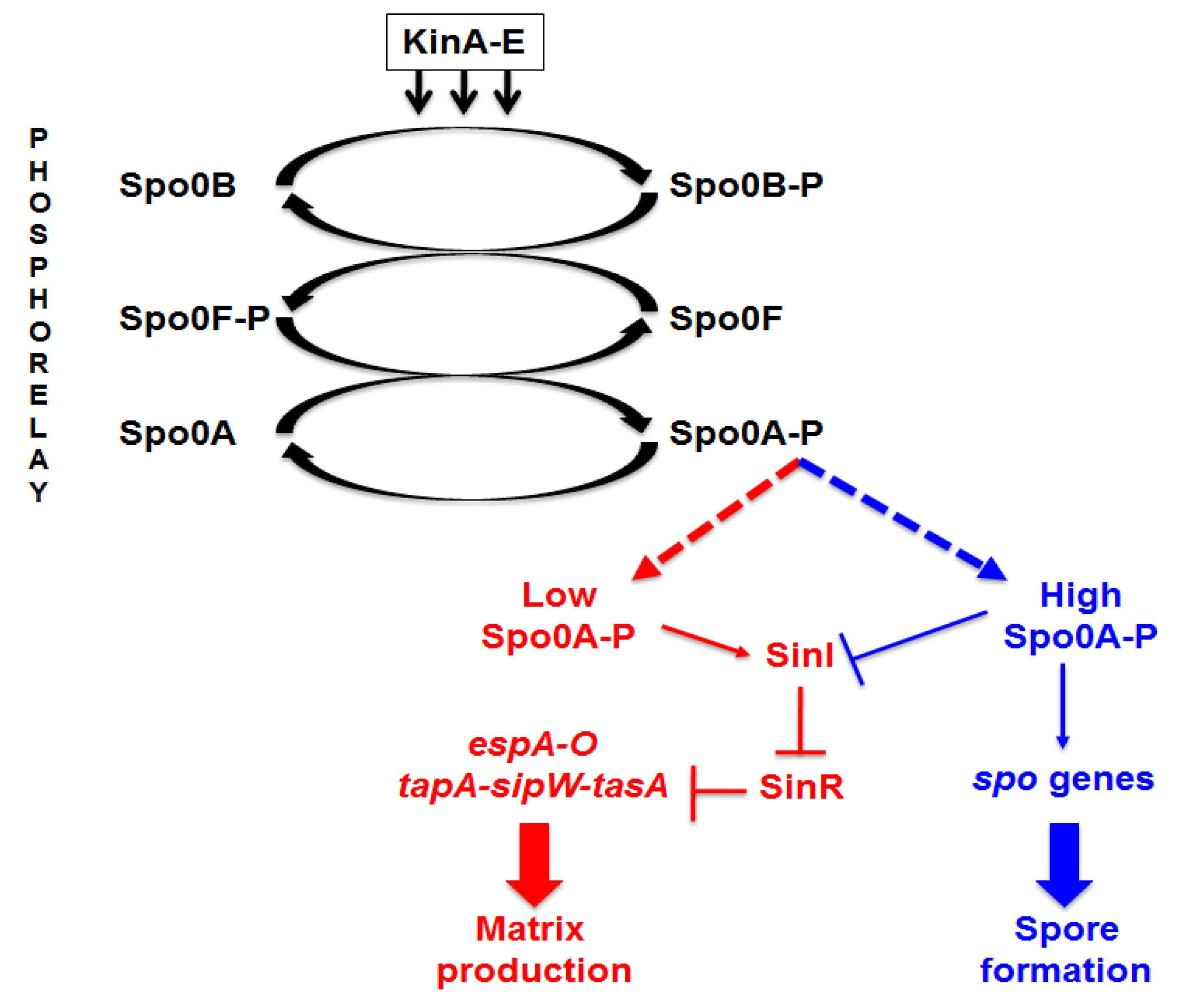
Figure 3A). A reduced production of biofilm in a medium that induces sporulation (DS), known to cause a rapid increase in the cellular concentration of Spo0A–P, is in agreement with the regulatory model previously reported for
B. subtilis [
10] and schematically summarized in
Figure 1. We next analyzed the composition of the matrix, investigating whether proteins and/or DNA are important matrix components by preventing their accumulation outside cells with protease and nuclease treatment as previously described for the matrix of
B. cereus [
14]. Proteinase K was more effective at preventing biofilm formation than DNase I, reducing the amount of biofilm by approximately 50% (
Figure 3B). This suggests that proteins play an important role in biofilm formation and thus confirms the similarities with
B. subtilis observed by the bioinformatic analysis.
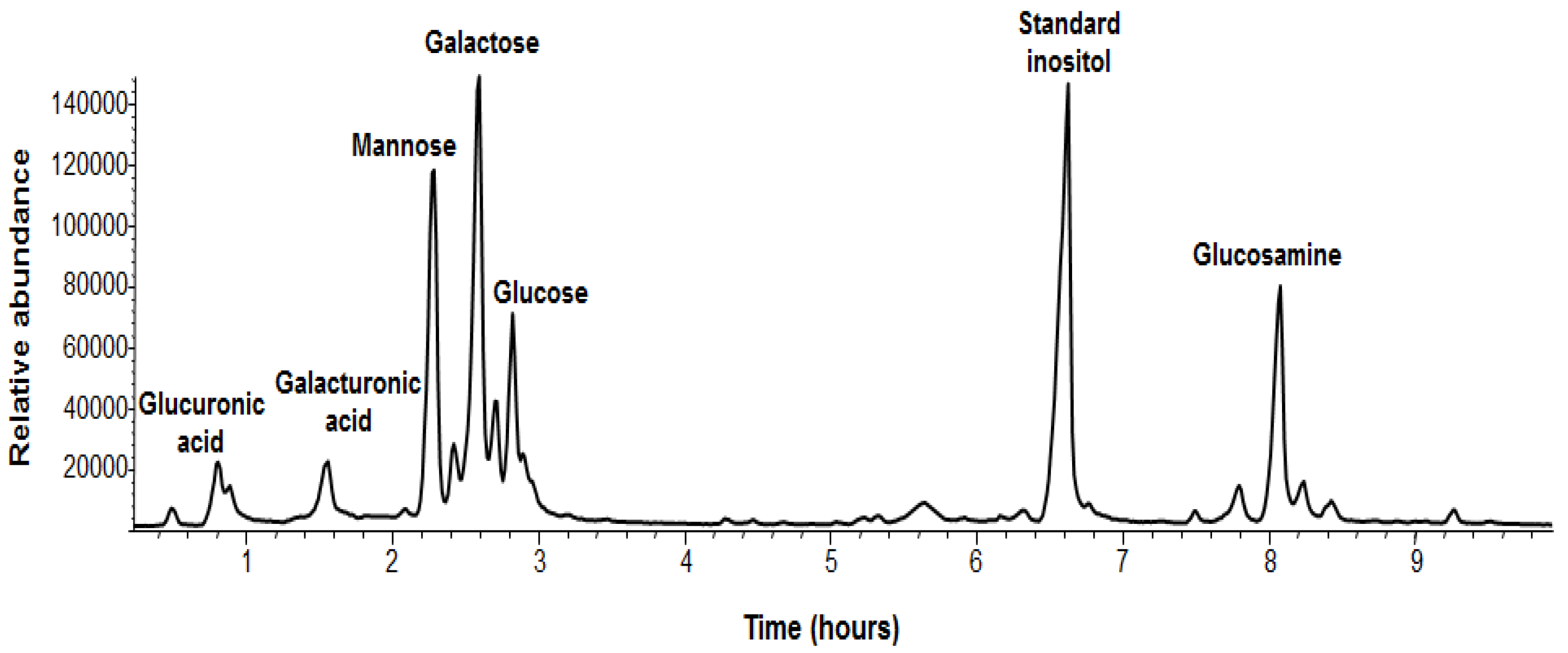
In addition, we analyzed the chemical composition of the sugars of the EPS of
B. pumilus SF214 (
Figure 4 and
Table 2). EPS monosaccharides were analyzed by GC-MS as acetylated
O-methyl glycosides. The sample was treated with 1.25 M MeOH/HCl and subsequently acetylated with acetic anhydride in pyridine and analyzed by GC-MS. The evaluation of the retention times and the fragmentation pattern from the GC-MS spectra compared with authentic standards allowed the identification of the monosaccharide residues. The sample contained galactose as its major component, together with minor amounts of mannose, 2-deoxy-2-amino glucose (glucosamine), galacturonic acid, and glucuronic acid (
Figure 4). The relative abundances of monosaccharides are shown in
Table 2.
Figure 3.
Determination of amount of biofilm produced by
B. pumilus SF214. (
A) Cells were grown at the indicated temperatures in rich (LB; light gray bars) or sporulation-inducing (DS; dark gray bars) media for 48 h. Cells were then removed, wells were stained and washed, and the OD (570 nm) was determined. (
B) Biofilm formation at 25 °C in LB medium supplemented with proteinase K or DNase I, as previously indicated [
14].
Figure 3.
Determination of amount of biofilm produced by
B. pumilus SF214. (
A) Cells were grown at the indicated temperatures in rich (LB; light gray bars) or sporulation-inducing (DS; dark gray bars) media for 48 h. Cells were then removed, wells were stained and washed, and the OD (570 nm) was determined. (
B) Biofilm formation at 25 °C in LB medium supplemented with proteinase K or DNase I, as previously indicated [
14].
Figure 4.
GC-MS chromatogram and assignments of B. pumilus SF214 strain biofilm monosaccharides. The non-indicated peaks in the chromatogram are different forms of the same monosaccharide derivative (i.e., α,β-furanosidic and pyranosidic forms).
Figure 4.
GC-MS chromatogram and assignments of B. pumilus SF214 strain biofilm monosaccharides. The non-indicated peaks in the chromatogram are different forms of the same monosaccharide derivative (i.e., α,β-furanosidic and pyranosidic forms).
Table 2.
Relative abundance of monosaccharides found in the matrix of B. pumilus SF214.
Table 2.
Relative abundance of monosaccharides found in the matrix of B. pumilus SF214.
| Monosaccharide | Relative Abundance a |
|---|
| Galactose | 1.00 |
| Mannose | 0.75 |
| Glucosamine | 0.54 |
| Glucose | 0.52 |
| Galacturonic acid | 0.21 |
| Glucuronic acid | 0.15 |
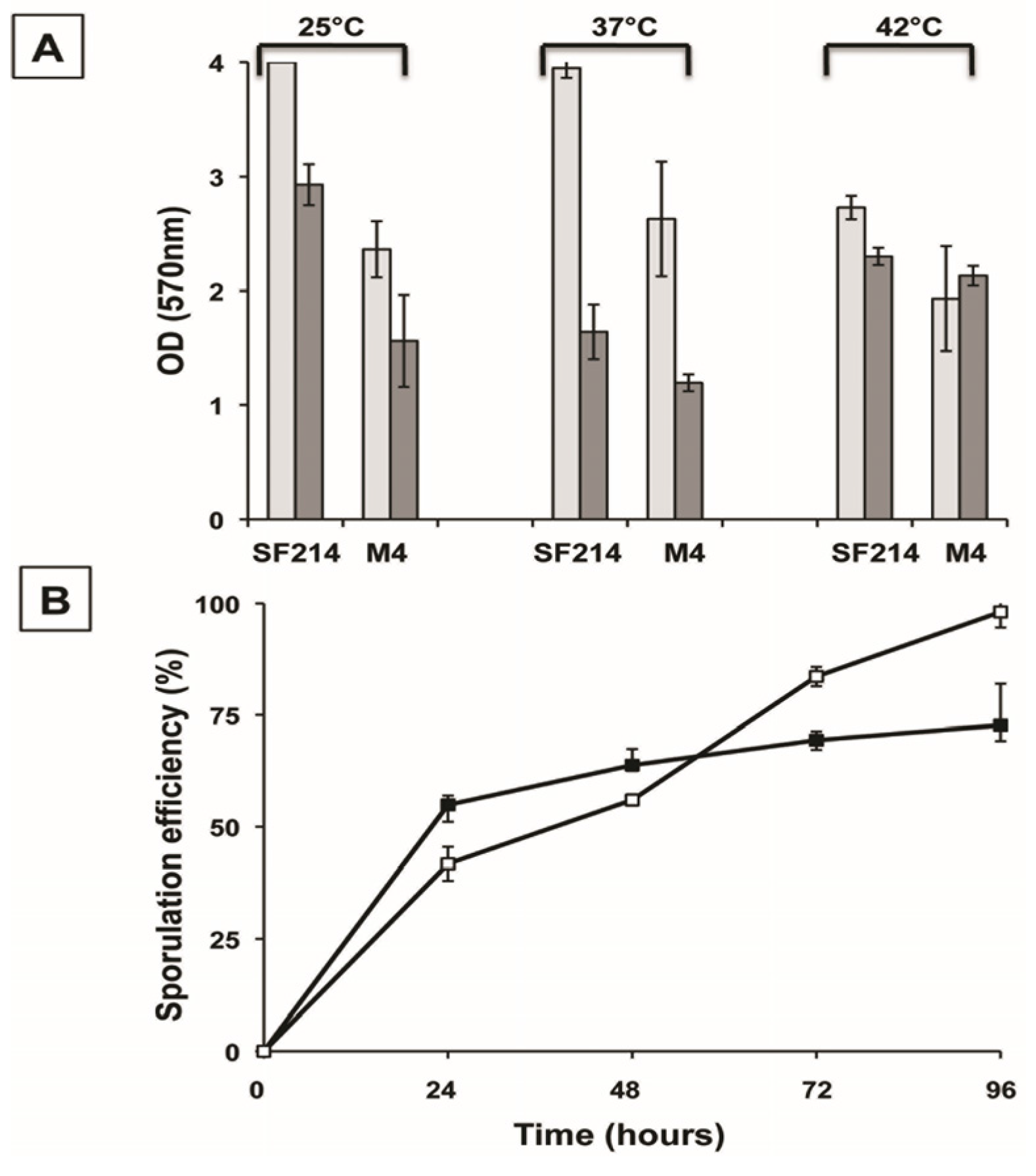
2.3. Mutants with Altered Pigmentation Are Also Altered in Matrix Formation and Sporulation
We have previously reported the isolation of an unpigmented mutant of strain SF214 after nitrosoguanidine (NTG) mutagenesis [
18]. One mutant (M4), carrying a single mutation, was used to demonstrate the anti-oxidative activity of the pigment [
18]. We used the unpigmented M4 mutant to measure biofilm formation and sporulation. Compared to its wild type (SF214), in the mutant strain (M4) biofilm production at 25 and 37 °C was strongly reduced in a rich (LB) medium, while a lesser reduction was observed at 42 °C (
Figure 5A). In a sporulation-inducing (DS) medium the wild type produced more biofilm than the mutant only at 25 °C, while at 37 and 42 °C similar amounts of biofilm were produced by the two strains (
Figure 5A). The efficiency of sporulation of SF214 and M4 strains is shown in
Figure 5B. Strain M4 showed an efficiency of sporulation similar to its wild-type strain for the first 48 h of growth and a slightly higher sporulation efficiency after 72 and 96 h. The efficiency of sporulation was also measured by counting the free spores under the light microscope at the various time points. Five microscope fields containing at least 200 cells/spores were randomly selected for each strain and cells and phase bright spores counted. Results are shown in
Supplementary Materials Figure S4 and were almost identical to those reported in
Figure 5B. Those results indicate that the mutation carried by strain M4, in addition to block pigment synthesis ([
18];
Figure 6A), also affects sporulation and biofilm formation. Based on this, we hypothesize that the mutation carried by the M4 strain is most likely in a gene coding for a regulatory factor rather than in one of the genes coding for an enzyme of the pigment biosynthetic pathway.
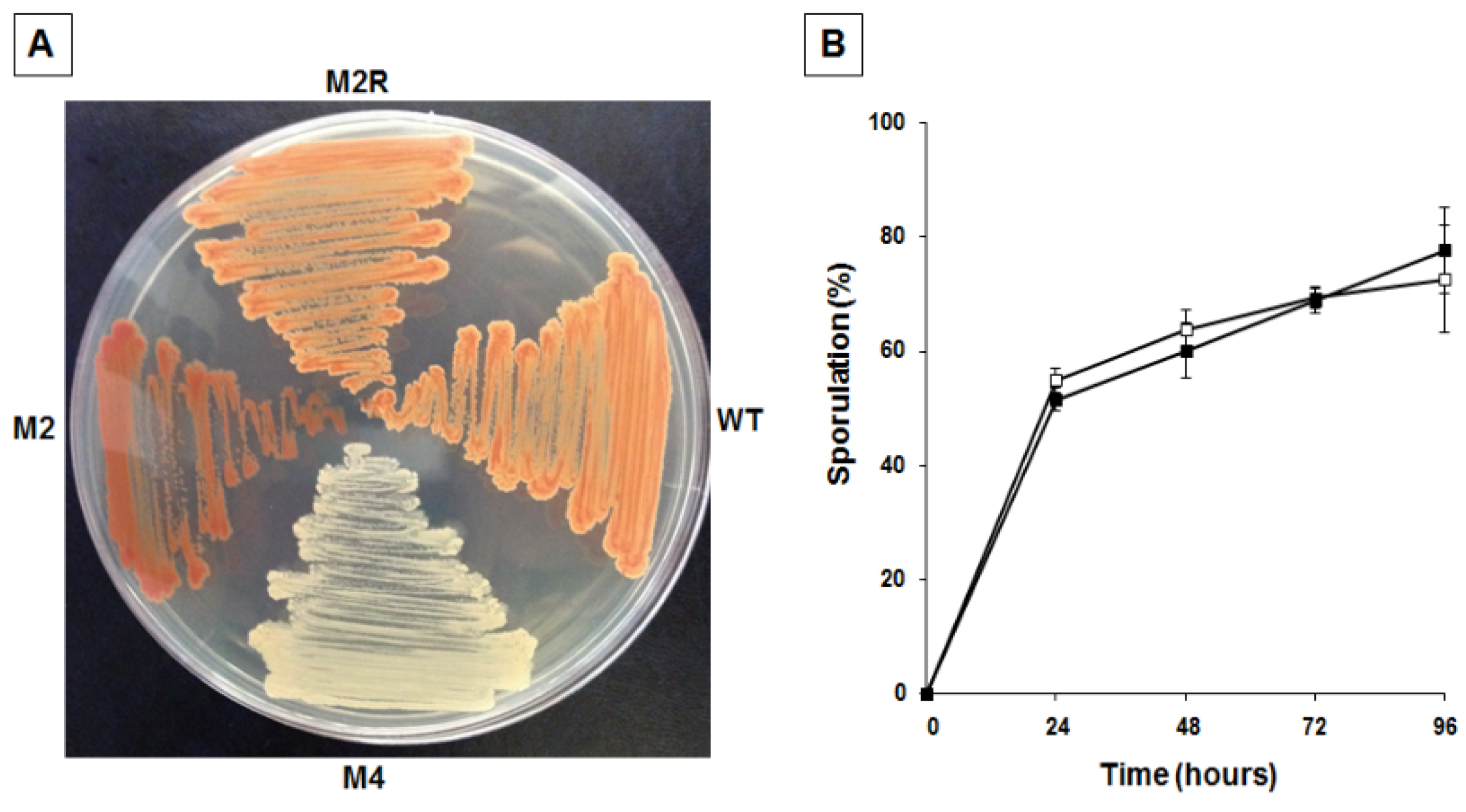
To confirm the link between pigmentation, matrix synthesis, and sporulation, we decided to identify and characterize other mutants with an altered pigmentation. Mid-exponential phase cells were incubated for different times with 10 mg of NTG and the percentage of survival assessed by colony forming unit (CFU) determination, as previously reported [
18]. To minimize the possibility of having mutants carrying multiple mutations, only cells exposed to NTG for the shortest time were considered. NTG-treated cells were then diluted, plated, and checked for pigmentation after 36 h of incubation at 25 °C. We focused on a single strongly over-pigmented mutant, M2. As previously reported [
18],
B. pumilus SF214 is refractory to genetic transformation, making it extremely difficult to identify the mutation carried by M2. A first phenotypic characterization of the mutant revealed that it was totally unable to produce spores. This surprising observation induced us to consider the possibility that we had either isolated a contaminant of some other bacterial species or that more than one mutation was carried by the M2 strain. To address these points we first analyzed and compared the DNA sequences of genes coding for the 16S RNA (not shown) and several other genes (see
Section 2.4) of SF214 and M2 strains. All M2 genes analyzed were identical to their counterparts in strain SF214 (see
Section 2.4), allowing us to exclude that M2 was a contaminant of a different bacterial species. Then, to evaluate whether more than one mutation was carried by the M2 strain we followed a classical genetic approach. We observed that the over-pigmented phenotype reverted spontaneously at a frequency of 1 clone out of 10
9. Then, we analyzed one revertant, M2R, and observed that it was identical to strain SF214 for its pigmentation (
Figure 6A) and for its efficiency in producing spores (
Figure 6B). Reversion of both phenotypes at a frequency of one clone out of 10
9 makes it extremely unlikely that the two phenotypes are due to more than one mutation.
Figure 5.
Biofilm formation (A) and sporulation efficiencies (B) of strain SF214 and M4 mutant. (A) Cells were grown at the indicated temperatures in rich (LB; light gray bars) or sporulation-inducing (DS; dark gray bars) media for 48 h. Cells were then removed, wells were stained, washed and the OD (570 nm) determined. (B) At the indicated times a sample of each growing culture (SF214 and M4 strain in black and white squares, respectively) was collected and split into two aliquots that were diluted and plated on LB plates. One of the aliquots was heat-treated (20 min at 80 °C) to kill all vegetative cells before plating. For each time point, the sporulation efficiency is the percentage of cells growing after the heat treatment, considering as 100% the number of cells growing without the heat treatment.
Figure 5.
Biofilm formation (A) and sporulation efficiencies (B) of strain SF214 and M4 mutant. (A) Cells were grown at the indicated temperatures in rich (LB; light gray bars) or sporulation-inducing (DS; dark gray bars) media for 48 h. Cells were then removed, wells were stained, washed and the OD (570 nm) determined. (B) At the indicated times a sample of each growing culture (SF214 and M4 strain in black and white squares, respectively) was collected and split into two aliquots that were diluted and plated on LB plates. One of the aliquots was heat-treated (20 min at 80 °C) to kill all vegetative cells before plating. For each time point, the sporulation efficiency is the percentage of cells growing after the heat treatment, considering as 100% the number of cells growing without the heat treatment.
Figure 6.
Pigmentation (A) and sporulation efficiency (B) of wild type and mutants of strain SF214. (A) Cells of wild type (wt), unpigmented (M4), over-pigmented (M2) mutants, and spontaneous revertant of M2 (M2R) were grown for 36 h at 25 °C on LB plate. (B) At the indicated times a sample of each growing culture (SF214 and M2R strain in black and white squares, respectively) was collected and split into two aliquots that were diluted and plated on LB plates. One of the aliquots was heat-treated to kill all vegetative cells before plating. For each time point, the sporulation efficiency is the percentage of cells growing after the heat treatment, considering as 100% the number of cells growing without the heat treatment.
Figure 6.
Pigmentation (A) and sporulation efficiency (B) of wild type and mutants of strain SF214. (A) Cells of wild type (wt), unpigmented (M4), over-pigmented (M2) mutants, and spontaneous revertant of M2 (M2R) were grown for 36 h at 25 °C on LB plate. (B) At the indicated times a sample of each growing culture (SF214 and M2R strain in black and white squares, respectively) was collected and split into two aliquots that were diluted and plated on LB plates. One of the aliquots was heat-treated to kill all vegetative cells before plating. For each time point, the sporulation efficiency is the percentage of cells growing after the heat treatment, considering as 100% the number of cells growing without the heat treatment.
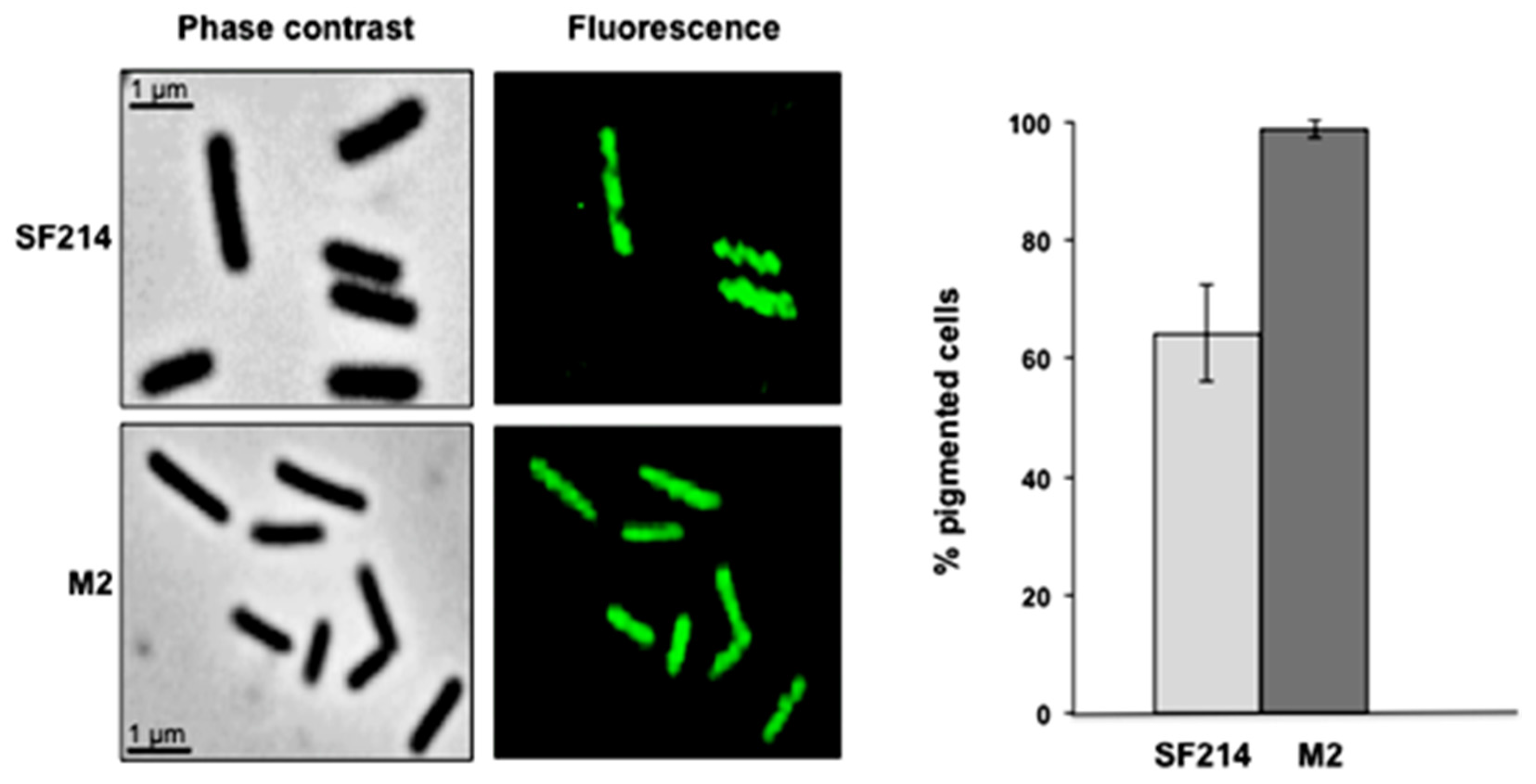
A fluorescence microscopy analysis of strains SF214 and M2 showed that the number of fluorescent cells of the mutant was strongly increased. Almost all mutant cells were fluorescent at conditions (25 °C in LB medium) in which slightly over 60% of the wild-type cells were pigmented (
Figure 7). Pigment production in SF214 is a bimodal process [
18] and the number of fluorescent cells is known to vary in response to environmental conditions [
18]. The increase in the number of fluorescent cells in M2 is an indication that the mutation carried by the M2 strain is not in a gene coding for a pigment biosynthetic enzyme but rather in a regulatory factor influencing the matrix synthesis/sporulation switch [
5].
Figure 7.
Pigmentation of strain SF214 and its mutant M2. Growing cells of strain SF214 and of the M2 mutant were analyzed by phase contrast and fluorescence microscopy. A representative microscopy field for each strain is shown by phase contrast and fluorescence microscopy. Over 1000 cells of 20 different microscopy fields for each strain were observed and the percentage of fluorescent cells reported.
Figure 7.
Pigmentation of strain SF214 and its mutant M2. Growing cells of strain SF214 and of the M2 mutant were analyzed by phase contrast and fluorescence microscopy. A representative microscopy field for each strain is shown by phase contrast and fluorescence microscopy. Over 1000 cells of 20 different microscopy fields for each strain were observed and the percentage of fluorescent cells reported.
A phenotypic analysis of M2 mutant cells showed that they are thinner (average width of 0.58 ± 0.1 and 0.45 ± 0.08 μm for SF214 and M2 strains, respectively) than wild-type cells. This phenotype has been associated with alteration in the efficiency of biofilm formation in
B. subtilis [
23,
24] as well as in other microorganisms [
25,
26]. To verify whether these phenotypes are also linked to matrix synthesis in
B. pumilus, we analyzed biofilm synthesis in strains SF214 and M2. As shown in
Figure 8, biofilm production was clearly reduced in M2 in a rich (LB) medium at 25 and 37 °C, while at 42 °C the situation was reversed, with the mutant producing more biofilm than strain SF214 (
Figure 8). When cells were grown in a sporulation-inducing (DS) medium, wild-type and mutant strains produced similar amounts of biofilm at all three temperatures tested (
Figure 8). In the experiments shown in
Figure 8 we also analyzed the amount of biofilm produced by the revertant strain M2R, which, as expected, behaved like the wild-type strains at all conditions tested (
Figure 8). Experiments shown in
Figure 8 then suggest that the observed changes in cell morphology are linked to a defect in matrix synthesis in the M2 mutant. In conclusion, the mutation carried by M2 increased pigment synthesis, blocked sporulation, and affected biofilm formation. Therefore, as in the case of the M4 mutant reported above, for the M2 mutant it is also more likely that the mutation is in a gene coding for regulatory factor rather than in one of the genes of the pigment biosynthetic pathway.
Figure 8.
Biofilm formation efficiency of strain SF214, M2 mutant, and M2R revertant. Cells were grown at the indicated temperatures in rich (LB; light gray bars) or sporulation-inducing (DS; dark gray bars) media for 48 h. Cells were then removed, wells were stained and washed, and the OD (570 nm) was determined.
Figure 8.
Biofilm formation efficiency of strain SF214, M2 mutant, and M2R revertant. Cells were grown at the indicated temperatures in rich (LB; light gray bars) or sporulation-inducing (DS; dark gray bars) media for 48 h. Cells were then removed, wells were stained and washed, and the OD (570 nm) was determined.
2.4. In Search of the Mutations
Due to the difficulties in manipulating strain SF214, the availability of genome data, and the possibility that the mutation carried by each of the two mutants could be in one of the regulatory factors determining the switch between sporulation and matrix production, we decided to analyze the DNA sequences of the genes coding for the main regulators controlling the spore/matrix switch indicated in
Figure 1. The analysis of strain SF214 genome showed that it contains homologs of the main regulators identified in
B. subtilis (
Table 3). The identity of the predicted products of the genes analyzed varied from 42% to 93% between
B. subtilis and
B. pumilus, while proteins of the two
B. pumilus strains were almost identical (
Table 3). Of the analyzed gene products, SinI and Spo0B are the least conserved ones between the two species (
Table 3).
Table 3.
Genes and putative proteins involved in the regulation of matrix synthesis and sporulation in B. subtilis 168, B. pumilus ATCC 7061T, and B. pumilus SF214.
Table 3.
Genes and putative proteins involved in the regulation of matrix synthesis and sporulation in B. subtilis 168, B. pumilus ATCC 7061T, and B. pumilus SF214.
| Genes | Putative Encoded Protein | Protein Identity (%) |
|---|
| B. subtilis 168 vs. B. pumilus ATCC 7061T | B. pumilus ATCC 7061T vs. B. pumilus SF214 |
|---|
| sinI | sinR antagonist | 45 | 96 |
| sinR | transcriptional regulator | 93 | 98 |
| spo0A | sporulation protein A | 88 | 100 |
| spo0B | phosphotransferase B | 58 | 98 |
| spo0F | phosphotransferase F | 92 | 100 |
All genes indicated in
Table 3 were sequenced in both M2 and M4 mutants (data not shown). No differences were identified in any of the genes analyzed, indicating that none of these genes was responsible for the observed phenotypes. These negative results indicate that other factors are involved in or influence the fate of stationary cells of the SF214 strain. These could either be
B. pumilus SF214-specific proteins or homologs of proteins characterized in
B. subtilis as affecting biofilm synthesis but not analyzed in this study. These include AbrB [
10], RemA-B [
27,
28], Veg [
29], and SlrA [
30], and also proteins only indirectly related to biofilm, such as collagen-like proteins [
31], components of the RapP-PhrP quorum sensing system [
32], and ribosomal proteins S11 and S21 [
33]. In addition, mutations in genes coding for alpha-phosphoglucomutase [
23], CcpA, glutamate synthase, GltAB, and the aminopeptidase AmpS [
34] have all been shown to have pleiotropic effects and also influence biofilm formation. All those genes are potential targets of the mutations carried by strains M2 and M4, making unrealistic the sequence-based approach to identify the mutated gene(s). In conclusion, the factor(s) controlling the biofilm-matrix switch in SF214 remains elusive and its identification is a challenging future task that will necessarily require the development of genetic tools to manipulate this pigmented marine microorganism.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





