
Production of a Novel Fucoidanase for the Green Synthesis of Gold Nanoparticles by Streptomyces sp. and Its Cytotoxic Effect on HeLa Cells


Abstract
:
1. Introduction
2. Results and Discussion
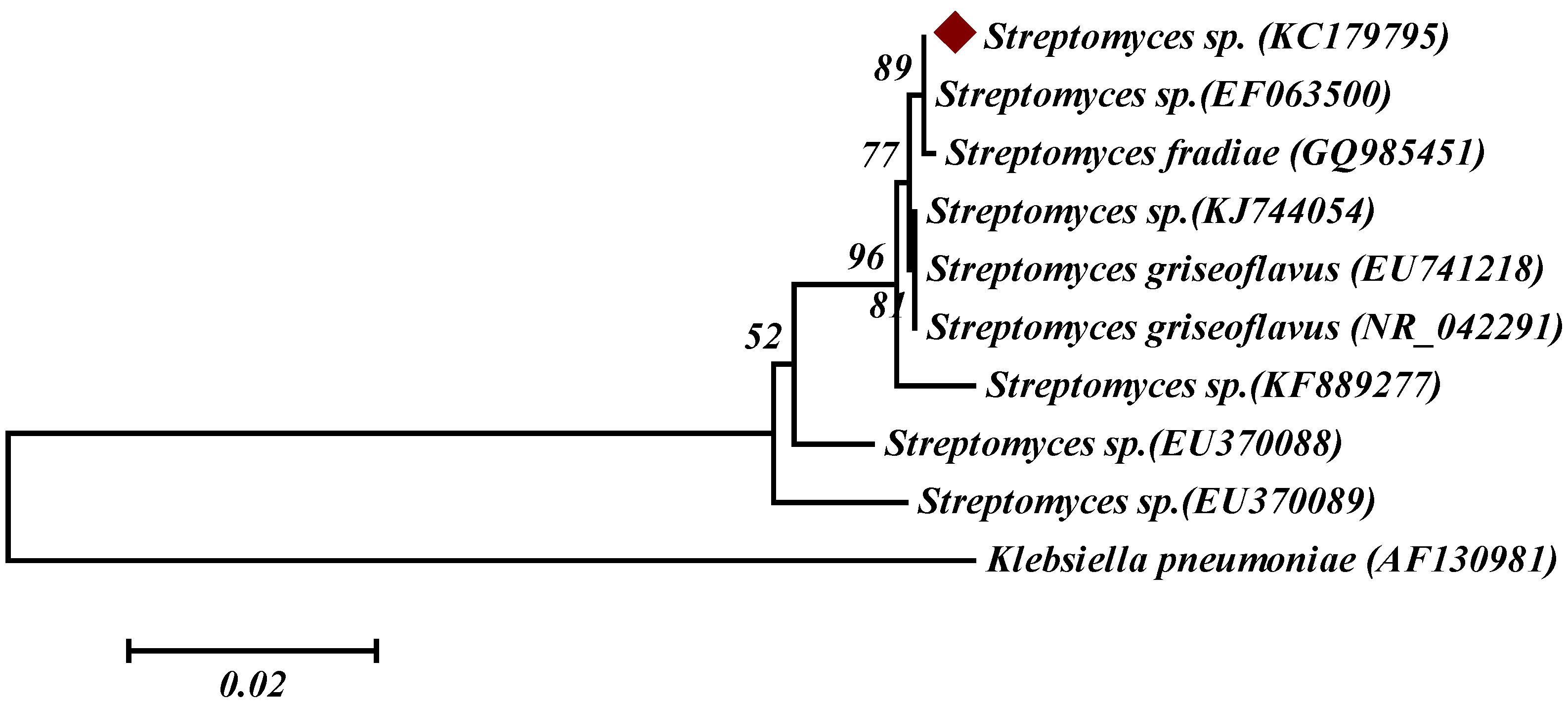
2.1. Isolation and Identification of Marine Actinobacterium

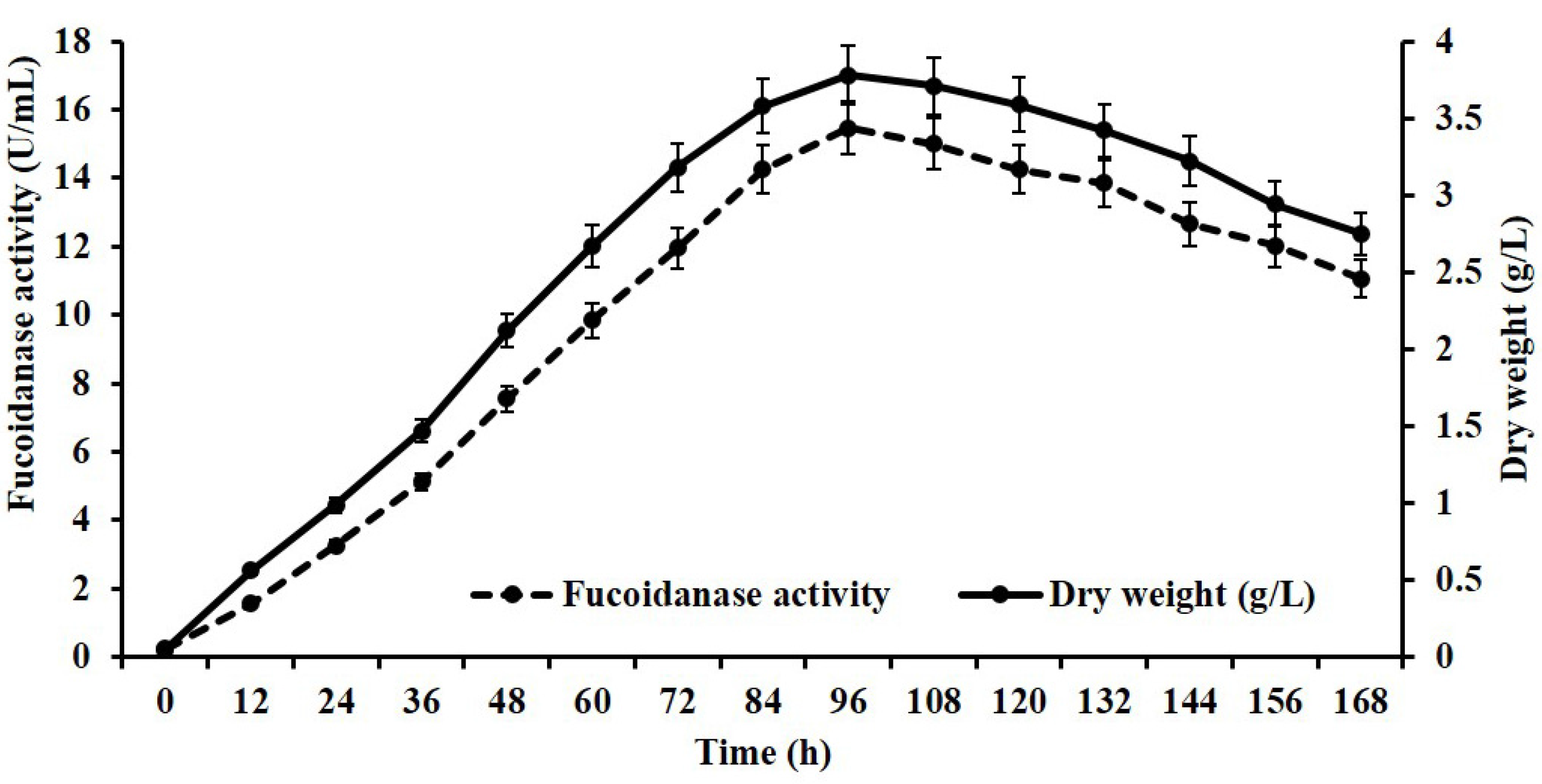
2.2. Fucoidanase Production

2.3. Plackett-Burman (PB) Design

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Variables | Concentration (g/L) | Effect | Coefficients | Standard Error | t-Value | p-Value | |
|---|---|---|---|---|---|---|---|---|
| −1 Level | +1 Level | |||||||
| X1 | Wheat bran | 1.0 | 5.0 | 2.003 | 1.002 | 0.1868 | 5.36 | 0.003 a,** |
| X2 | Kelp powder | 0.1 | 1.0 | −1.383 | −0.692 | 0.1868 | −3.70 | 0.014 b,* |
| X3 | Glucose | 0.1 | 0.5 | −2.023 | −1.012 | 0.1868 | −5.42 | 0.003 b,** |
| X4 | NaNO3 | 0.01 | 0.05 | 4.320 | 2.160 | 0.1868 | 11.56 | 0.000 a,** |
| X5 | MgSO4·7H2O | 0.01 | 0.05 | 2.860 | 1.430 | 0.1868 | 7.66 | 0.001 a,** |
| X6 | NaCl | 0.1 | 1.0 | 5.110 | 2.555 | 0.1868 | 13.68 | 0.000 a,** |
| Runs | Real Levels (Coded Levels)/Concentrations (g/L) | Fucoidanase Activity (U/mL) | ||||||
|---|---|---|---|---|---|---|---|---|
| X1 | X2 | X3 | X4 | X5 | X6 | Observed a | Predicted | |
| 1 | 5 (+1) | 0.1 (−1) | 0.5 (+1) | 0.01 (−1) | 0.01 (−1) | 0.1 (−1) | 2.83 | 2.78 |
| 2 | 5 (+1) | 1.0 (+1) | 0.1 (−1) | 0.05 (+1) | 0.01 (−1) | 0.1 (−1) | 7.71 | 7.74 |
| 3 | 1 (−1) | 1.0 (+1) | 0.5 (+1) | 0.01 (−1) | 0.05 (+1) | 0.1 (−1) | 2.42 | 2.25 |
| 4 | 5 (+1) | 0.1 (−1) | 0.5 (+1) | 0.05 (+1) | 0.01 (−1) | 1.0 (+1) | 12.56 | 12.21 |
| 5 | 5 (+1) | 1.0 (+1) | 0.1 (−1) | 0.05 (+1) | 0.05 (+1) | 0.1 (−1) | 10.62 | 10.60 |
| 6 | 5 (+1) | 1.0 (+1) | 0.5 (+1) | 0.01 (−1) | 0.05 (+1) | 1.0 (+1) | 8.91 | 9.36 |
| 7 | 1 (−1) | 1.0 (+1) | 0.5 (+1) | 0.05 (+1) | 0.01 (−1) | 1.0 (+1) | 8.25 | 8.82 |
| 8 | 1 (−1) | 0.1 (−1) | 0.5 (+1) | 0.05 (+1) | 0.05 (+1) | 0.1 (−1) | 8.39 | 7.95 |
| 9 | 1 (−1) | 0.1 (−1) | 0.1 (−1) | 0.05 (+1) | 0.05 (+1) | 1.0 (+1) | 14.86 | 15.09 |
| 10 | 5 (+1) | 0.1 (−1) | 0.1 (−1) | 0.01 (−1) | 0.05 (+1) | 1.0 (+1) | 12.81 | 12.77 |
| 11 | 1 (−1) | 1.0 (+1) | 0.1 (−1) | 0.01 (−1) | 0.01 (−1) | 1.0 (+1) | 7.37 | 6.52 |
| 12 | 1 (−1) | 0.1 (−1) | 0.1 (−1) | 0.01 (−1) | 0.01 (−1) | 0.1 (−1) | 2.13 | 2.80 |
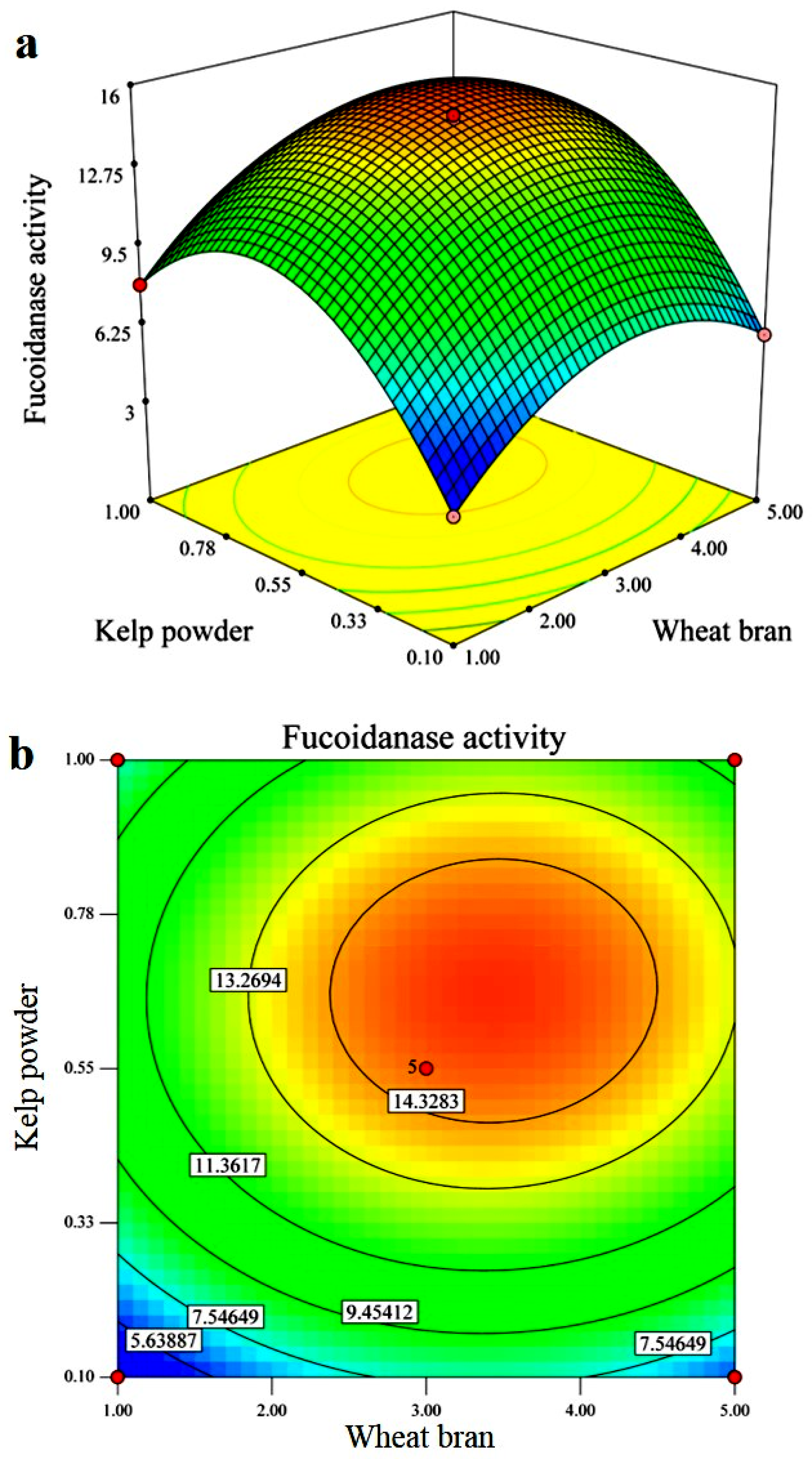
2.4. Response Surface Methodology (RSM)
| Trials | X1 | X2 | X6 | Fucoidanase Activity (U/mL) | |
|---|---|---|---|---|---|
| Observed a | Predicted | ||||
| 1 | 1 | 0.10 | 0.55 | 3.71 | 3.73 |
| 2 | 5 | 0.10 | 0.55 | 5.83 | 5.84 |
| 3 | 1 | 1.00 | 0.55 | 7.92 | 7.91 |
| 4 | 5 | 1.00 | 0.55 | 10.88 | 10.86 |
| 5 | 1 | 0.55 | 0.10 | 9.89 | 9.81 |
| 6 | 5 | 0.55 | 0.10 | 13.56 | 13.48 |
| 7 | 1 | 0.55 | 1.00 | 14.38 | 14.46 |
| 8 | 5 | 0.55 | 1.00 | 15.76 | 15.85 |
| 9 | 3 | 0.10 | 0.10 | 8.51 | 8.57 |
| 10 | 3 | 1.00 | 0.10 | 11.36 | 11.46 |
| 11 | 3 | 0.10 | 1.00 | 10.47 | 10.37 |
| 12 | 3 | 1.00 | 1.00 | 16.74 | 16.68 |
| 13 | 3 | 0.55 | 0.55 | 14.52 | 14.75 |
| 14 | 3 | 0.55 | 0.55 | 14.83 | 14.75 |
| 15 | 3 | 0.55 | 0.55 | 14.79 | 14.75 |
| 16 | 3 | 0.55 | 0.55 | 14.81 | 14.75 |
| 17 | 3 | 0.55 | 0.55 | 14.80 | 14.75 |
| Source | Sum of Squares | DF | Mean Square | F-Value | p-Value > F |
|---|---|---|---|---|---|
| Model | 226.45 | 9 | 25.16 | 1439.22 | <0.0001 ** |
| X1 | 12.83 | 1 | 12.83 | 733.73 | <0.0001 ** |
| X2 | 42.23 | 1 | 42.23 | 2415.50 | <0.0001 ** |
| X6 | 24.61 | 1 | 24.61 | 1407.44 | <0.0001 ** |
| X1X2 | 0.18 | 1 | 0.18 | 10.09 | <0.0156 * |
| X1X6 | 1.31 | 1 | 1.31 | 74.99 | <0.0001 ** |
| X2X6 | 2.92 | 1 | 2.92 | 167.26 | <0.0001 ** |
| X21 | 38.37 | 1 | 38.37 | 2194.81 | <0.0001 ** |
| X22 | 90.90 | 1 | 90.90 | 5199.32 | <0.0001 ** |
| X26 | 11.69 | 1 | 11.69 | 668.69 | <0.0001 ** |
| Residual | 0.12 | 7 | 0.017 | ||
| Lack-of-fit | 0.055 | 3 | 0.018 | 1.10 | 0.4456 NS |
| Pure error | 0.067 | 4 | 0.017 | ||
| Cor total | 226.57 | 16 |

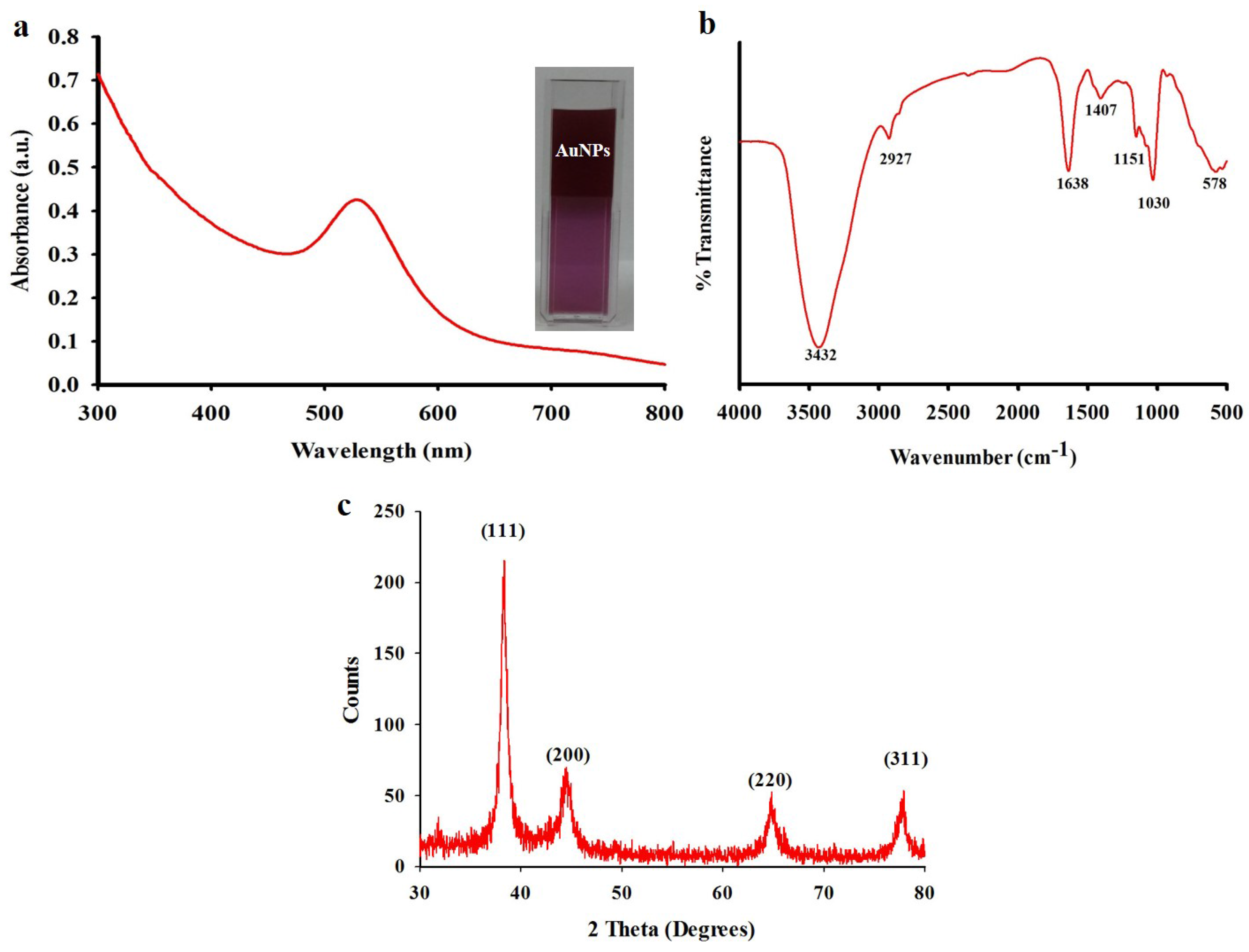
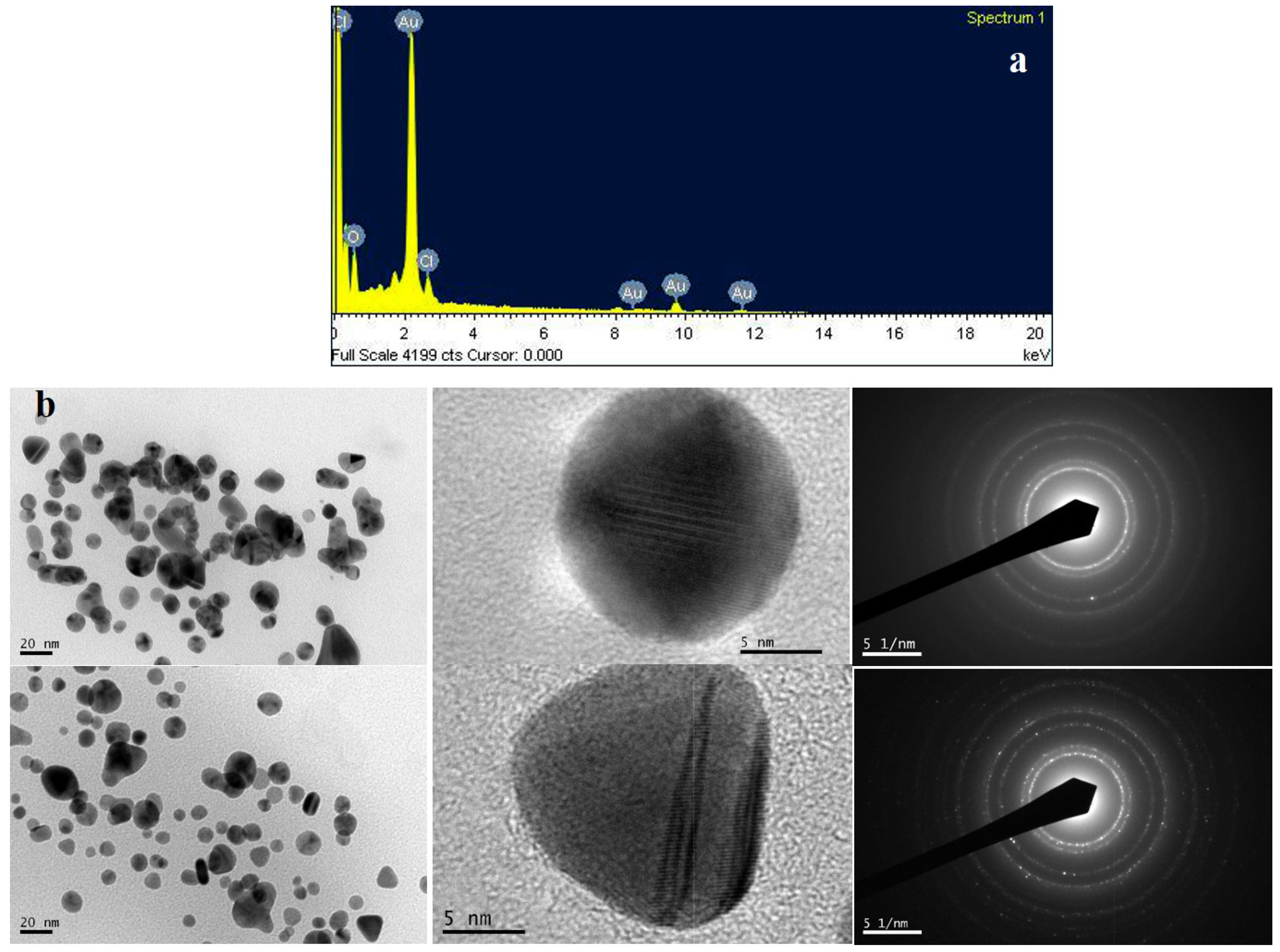
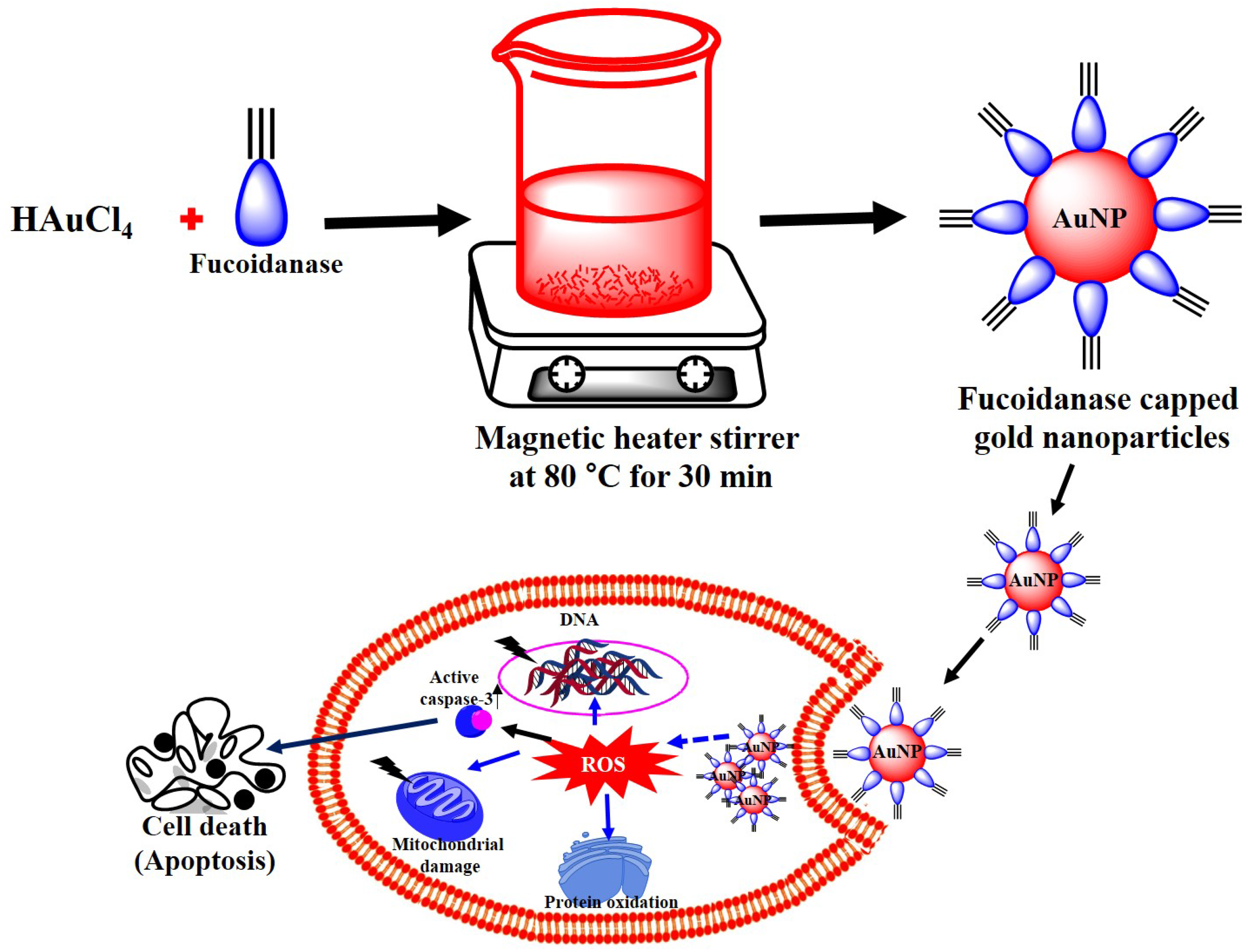
2.5. Biosynthesis and Characterization of Gold Nanoparticles


2.6. In Vitro Anticancer Activity of Gold Nanoparticles
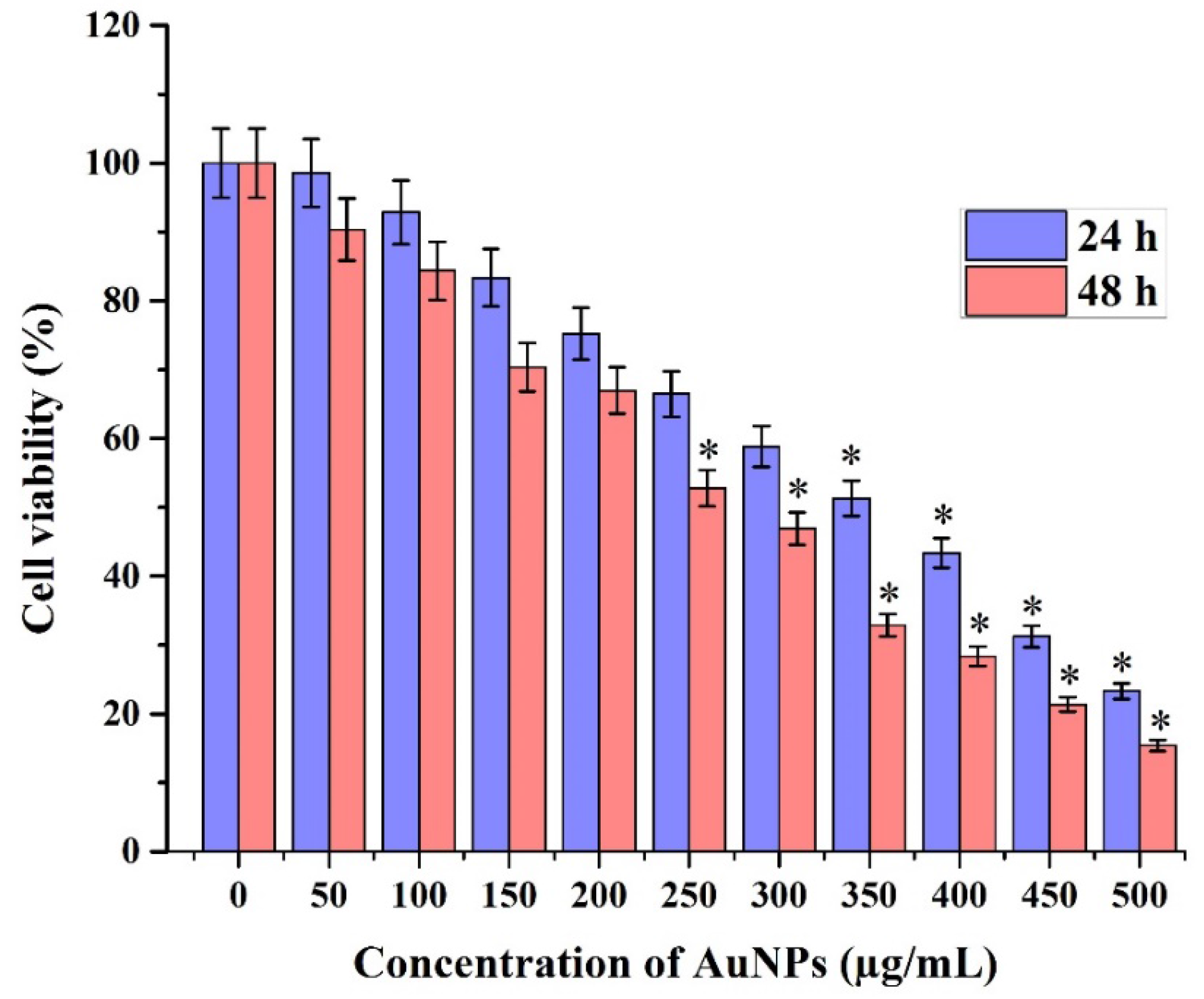
2.6.1. Cell Viability

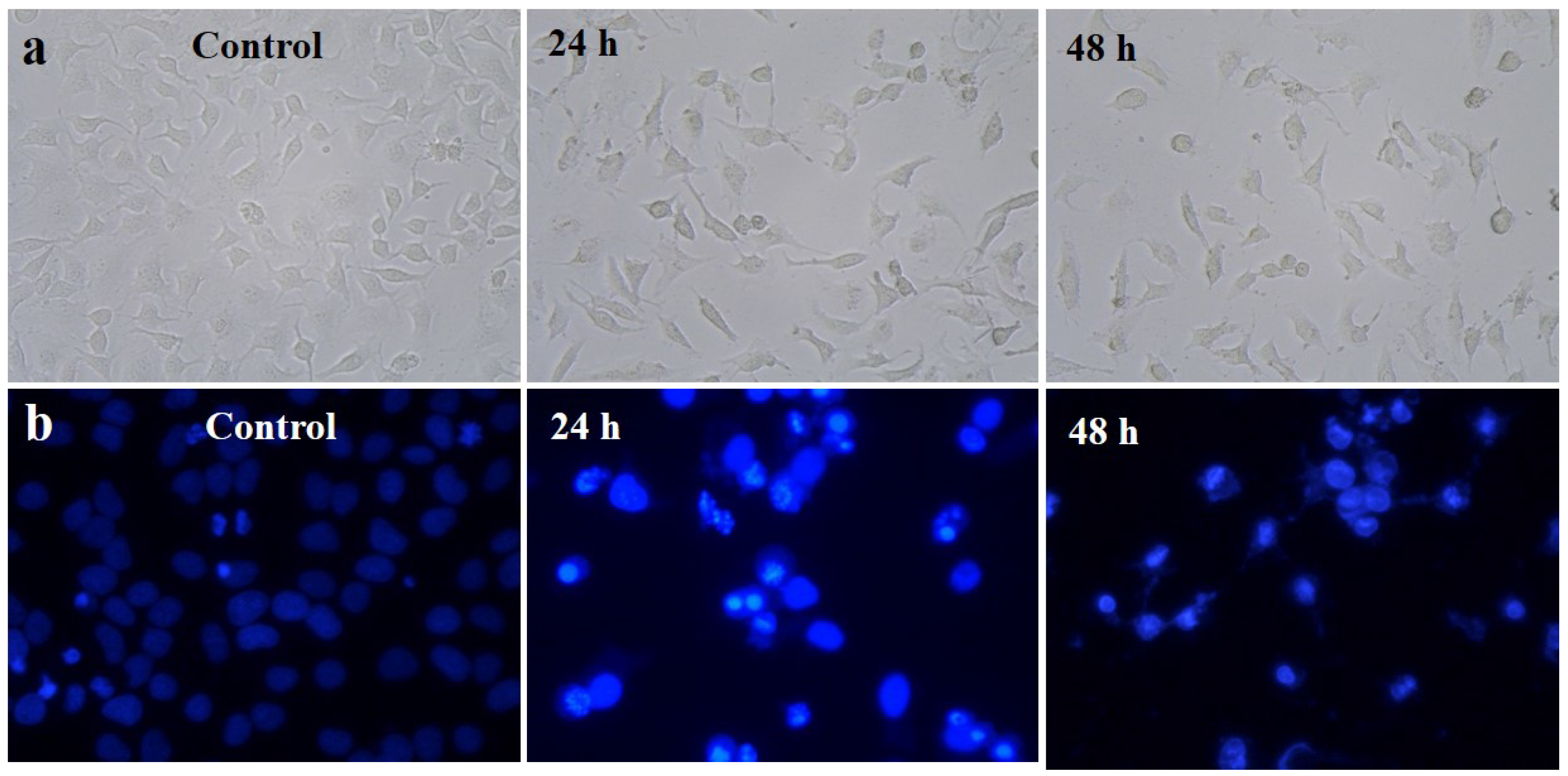
2.6.2. Morphological Observation

2.6.3. 4′,6-Diamidino-2-Phenylindole Dihydrochloride (DAPI) for Nuclear Staining

3. Materials and Methods
3.1. Chemicals
3.2. Isolation and Identification of Marine Actinobacterium
3.3. Fucoidanase Production
3.4. Enzyme Assay
3.4.1. Fucoidanase Activity
3.4.2. Fucosidase Activity
3.4.3. Amylase Assay
3.5. Experimental Design and Data Analysis
3.5.1. Plackett-Burman (PB) Design
3.5.2. Response Surface Methodology (RSM)
3.6. Purification of Fucoidanase
3.7. Biosynthesis and Characterization of Gold Nanoparticles
3.8. In Vitro Anticancer Activity of Gold Nanoparticles
3.8.1. Cell Viability
3.8.2. Morphological Observation
3.8.3. 4′,6-Diamidino-2-Phenylindole Dihydrochloride (DAPI) Staining
3.8.4. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Holtkamp, A.D.; Kelly, S.; Ulber, R.; Lang, S. Fucoidans and fucoidanases—Focus on techniques for molecular structure elucidation and modification of marine polysaccharides. Appl. Microbiol. Biotechnol. 2009, 82, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Senni, K.; Pereira, J.; Gueniche, F.; Delbarre-Ladrat, C.; Sinquin, C.; Ratiskol, J.; Godeau, G.; Fischer, A.M.; Helley, D.; Colliec-Jouault, S. Marine polysaccharides: A source of bioactive molecules for cell therapy and tissue engineering. Mar. Drugs 2011, 9, 1664–1681. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, I.; Queiroz, K.; Alves, L.; Santos, E.; Leite, E.; Rocha, H. Heterofucans from Dictyota menstrualis have anticoagulant activity. Braz. J. Med. Biol. Res. 2004, 37, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Vishchuk, O.S.; Ermakova, S.P.; Zvyagintseva, T.N. Sulfated polysaccharides from brown seaweeds Saccharina japonica and Undaria pinnatifida: Isolation, structural characteristics, and antitumor activity. Carbohydr. Res. 2011, 346, 2769–2776. [Google Scholar] [CrossRef] [PubMed]
- Shibata, H.; Kimura-Takagi, I.; Nagaoka, M.; Hashimoto, S.; Sawada, H.; Ueyama, S.; Yokokura, T. Inhibitory effect of Cladosiphon fucoidan on the adhesion of Helicobacter pylori to human gastric cells. J. Nutr. Sci. Vitaminol. 1999, 45, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, U.; Mateu, C.G.; Chattopadhyay, K.; Pujol, C.A.; Damonte, E.B.; Ray, B. Structure and antiviral activity of sulfated fucans from Stoechospermum marginatum. Phytochemistry 2006, 67, 2474–2482. [Google Scholar] [CrossRef] [PubMed]
- Cumashi, A.; Ushakova, N.A.; Preobrazhenskaya, M.E.; D’Incecco, A.; Piccoli, A.; Totani, L.; Tinari, N.; Morozevich, G.E.; Berman, A.E.; Bilan, M.I. A comparative study of the anti-inflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology 2007, 17, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Mourao, P.A. Use of sulfated fucans as anticoagulant and antithrombotic agents: Future perspectives. Curr. Pharm. Des. 2004, 10, 967–981. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Zhang, M.; Wu, K.; Liu, B.; Cai, J.; Pan, R. Purification and characteristics of fucoidanase obtained from Dendryphiella arenaria TM94. J. Appl. Phycol. 2011, 23, 197–203. [Google Scholar] [CrossRef]
- Kusaykin, M.; Chizhov, A.; Grachev, A.; Alekseeva, S.; Bakunina, I.Y.; Nedashkovskaya, O.; Sova, V.; Zvyagintseva, T. A comparative study of specificity of fucoidanases from marine microorganisms and invertebrates. J. Appl. Phycol. 2006, 18, 369–373. [Google Scholar] [CrossRef]
- Bakunina, I.Y.; Nedashkovskaya, O.; Alekseeva, S.; Ivanova, E.; Romanenko, L.; Gorshkova, N.; Isakov, V.; Zvyagintseva, T.; Mikhailov, V. Degradation of fucoidan by the marine proteobacterium Pseudoalteromonas citrea. Microbiology 2002, 71, 41–47. [Google Scholar] [CrossRef]
- Sakai, T.; Kawai, T.; Kato, I. Isolation and characterization of a fucoidan-degrading marine bacterial strain and its fucoidanase. Mar. Biotechnol. 2004, 6, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Bakunina, I.Y.; Shevchenko, L.; Nedashkovskaya, O.; Shevchenko, N.; Alekseeva, S.; Mikhailov, V.; Zvyagintseva, T. Screening of marine bacteria for fucoidanases. Microbiology 2000, 69, 303–308. [Google Scholar] [CrossRef]
- Rodríguez-Jasso, R.M.; Mussatto, S.I.; Sepúlveda, L.; Agrasar, A.T.; Pastrana, L.; Aguilar, C.N.; Teixeira, J.A. Fungal fucoidanase production by solid-state fermentation in a rotating drum bioreactor using algal biomass as substrate. Food Bioprod. Process. 2013, 91, 587–594. [Google Scholar] [CrossRef]
- Qianqian, W.; Shuang, M.; Hourong, X.; Min, Z.; Jingmin, C. Purification and the secondary structure of fucoidanase from Fusarium sp. LD8. Evid. Based Complement. Altern. Med. 2011, 2011, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Alexeeva, Y.; Ivanova, E.; Bakunina, I.Y.; Zvaygintseva, T.; Mikhailov, V. Optimization of glycosidases production by Pseudoalteromonas issachenkonii KMM 3549T. Lett. Appl. Microbiol. 2002, 35, 343–346. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.H. Gold nanoparticle-based immunochromatographic test for identification of Staphylococcus aureus from clinical specimens. Clin. Chim. Acta 2006, 373, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Manivasagan, P.; Venkatesan, J.; Kang, K.H.; Sivakumar, K.; Park, S.J.; Kim, S.K. Production of α-amylase for the biosynthesis of gold nanoparticles using Streptomyces sp. MBRC-82. Int. J. Biol. Macromol. 2015, 72, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Manivasagan, P.; Alam, M.S.; Kang, K.H.; Kwak, M.; Kim, S.K. Extracellular synthesis of gold bionanoparticles by Nocardiopsis sp. and evaluation of its antimicrobial, antioxidant and cytotoxic activities. Bioprocess Biosyst. Eng. 2015, 38, 1167–1177. [Google Scholar] [CrossRef] [PubMed]
- Manivasagan, P.; Kang, K.H.; Kim, D.G.; Kim, S.K. Production of polysaccharide-based bioflocculant for the synthesis of silver nanoparticles by Streptomyces sp. Int. J. Biol. Macromol. 2015, 77, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Manivasagan, P.; Venkatesan, J.; Sivakumar, K.; Kim, S.K. Actinobacterial enzyme inhibitors—A review. Crit. Rev. Microbiol. 2015, 41, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Shanmugasundaram, T.; Radhakrishnan, M.; Gopikrishnan, V.; Pazhanimurugan, R.; Balagurunathan, R. A study of the bactericidal, anti-biofouling, cytotoxic and antioxidant properties of actinobacterially synthesised silver nanoparticles. Colloid Surf. B Biointerfaces 2013, 111, 680–687. [Google Scholar] [CrossRef] [PubMed]
- Gopal, J.V.; Thenmozhi, M.; Kannabiran, K.; Rajakumar, G.; Velayutham, K.; Rahuman, A.A. Actinobacteria mediated synthesis of gold nanoparticles using Streptomyces sp. VITDDK3 and its antifungal activity. Mater. Lett. 2013, 93, 360–362. [Google Scholar] [CrossRef]
- Amin, M.; Iram, F.; Iqbal, M.S.; Saeed, M.Z.; Raza, M.; Alam, S. Arabinoxylan-mediated synthesis of gold and silver nanoparticles having exceptional high stability. Carbohydr. Polym. 2013, 92, 1896–1900. [Google Scholar] [CrossRef] [PubMed]
- Ganeshkumar, M.; Sathishkumar, M.; Ponrasu, T.; Dinesh, M.G.; Suguna, L. Spontaneous ultra fast synthesis of gold nanoparticles using Punica granatum for cancer targeted drug delivery. Colloid Surf. B Biointerfaces 2013, 106, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Suman, T.; Rajasree, S.R.; Kanchana, A.; Elizabeth, S.B. Biosynthesis, characterization and cytotoxic effect of plant mediated silver nanoparticles using Morinda citrifolia root extract. Colloid Surf. B Biointerfaces 2013, 106, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Dhas, T.S.; Kumar, V.G.; Karthick, V.; Govindaraju, K.; Narayana, T.S. Biosynthesis of gold nanoparticles using Sargassum swartzii and its cytotoxicity effect on HeLa cells. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2014, 133, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Venkatpurwar, V.; Shiras, A.; Pokharkar, V. Porphyran capped gold nanoparticles as a novel carrier for delivery of anticancer drug: In vitro cytotoxicity study. Int. J. Pharm. 2011, 409, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Lokina, S.; Suresh, R.; Giribabu, K.; Stephen, A.; Sundaram, R.L.; Narayanan, V. Spectroscopic investigations, antimicrobial, and cytotoxic activity of green synthesized gold nanoparticles. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2014, 129, 484–490. [Google Scholar] [CrossRef] [PubMed]
- Muthuvel, A.; Adavallan, K.; Balamurugan, K.; Krishnakumar, N. Biosynthesis of gold nanoparticles using Solanum nigrum leaf extract and screening their free radical scavenging and antibacterial properties. Biomed. Prev. Nutr. 2014, 4, 325–332. [Google Scholar] [CrossRef]
- Raghunandan, D.; Ravishankar, B.; Sharanbasava, G.; Mahesh, D.B.; Harsoor, V.; Yalagatti, M.S.; Bhagawanraju, M.; Venkataraman, A. Anti-cancer studies of noble metal nanoparticles synthesized using different plant extracts. Cancer Nanotechnol. 2011, 2, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.; Kusumoto, Y.; Al-Mamun, M.A. Cytotoxicity and cancer (HeLa) cell killing efficacy of aqueous garlic (Allium sativum) extract. J. Sci. Res. 2011, 3, 375–382. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Liu, S.; Fang, Y.; Lv, M.; Wang, S.; Chen, L. Optimization of the production of organic solvent-stable protease by Bacillus sphaericus DS11 with response surface methodology. Bioresour. Technol. 2010, 101, 7924–7929. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, D.; Arulvasu, C.; Babu, G.; Manikandan, R.; Srinivasan, P. Biologically synthesized green silver nanoparticles from leaf extract of Vitex negundo L. induce growth-inhibitory effect on human colon cancer cell line HCT15. Process Biochem. 2013, 48, 317–324. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manivasagan, P.; Oh, J. Production of a Novel Fucoidanase for the Green Synthesis of Gold Nanoparticles by Streptomyces sp. and Its Cytotoxic Effect on HeLa Cells. Mar. Drugs 2015, 13, 6818-6837. https://doi.org/10.3390/md13116818
Manivasagan P, Oh J. Production of a Novel Fucoidanase for the Green Synthesis of Gold Nanoparticles by Streptomyces sp. and Its Cytotoxic Effect on HeLa Cells. Marine Drugs. 2015; 13(11):6818-6837. https://doi.org/10.3390/md13116818
Chicago/Turabian StyleManivasagan, Panchanathan, and Junghwan Oh. 2015. "Production of a Novel Fucoidanase for the Green Synthesis of Gold Nanoparticles by Streptomyces sp. and Its Cytotoxic Effect on HeLa Cells" Marine Drugs 13, no. 11: 6818-6837. https://doi.org/10.3390/md13116818
APA StyleManivasagan, P., & Oh, J. (2015). Production of a Novel Fucoidanase for the Green Synthesis of Gold Nanoparticles by Streptomyces sp. and Its Cytotoxic Effect on HeLa Cells. Marine Drugs, 13(11), 6818-6837. https://doi.org/10.3390/md13116818