
New Insights on the Terpenome of the Red Seaweed Laurencia dendroidea (Florideophyceae, Rhodophyta)

Abstract
:1. Introduction
2. Results

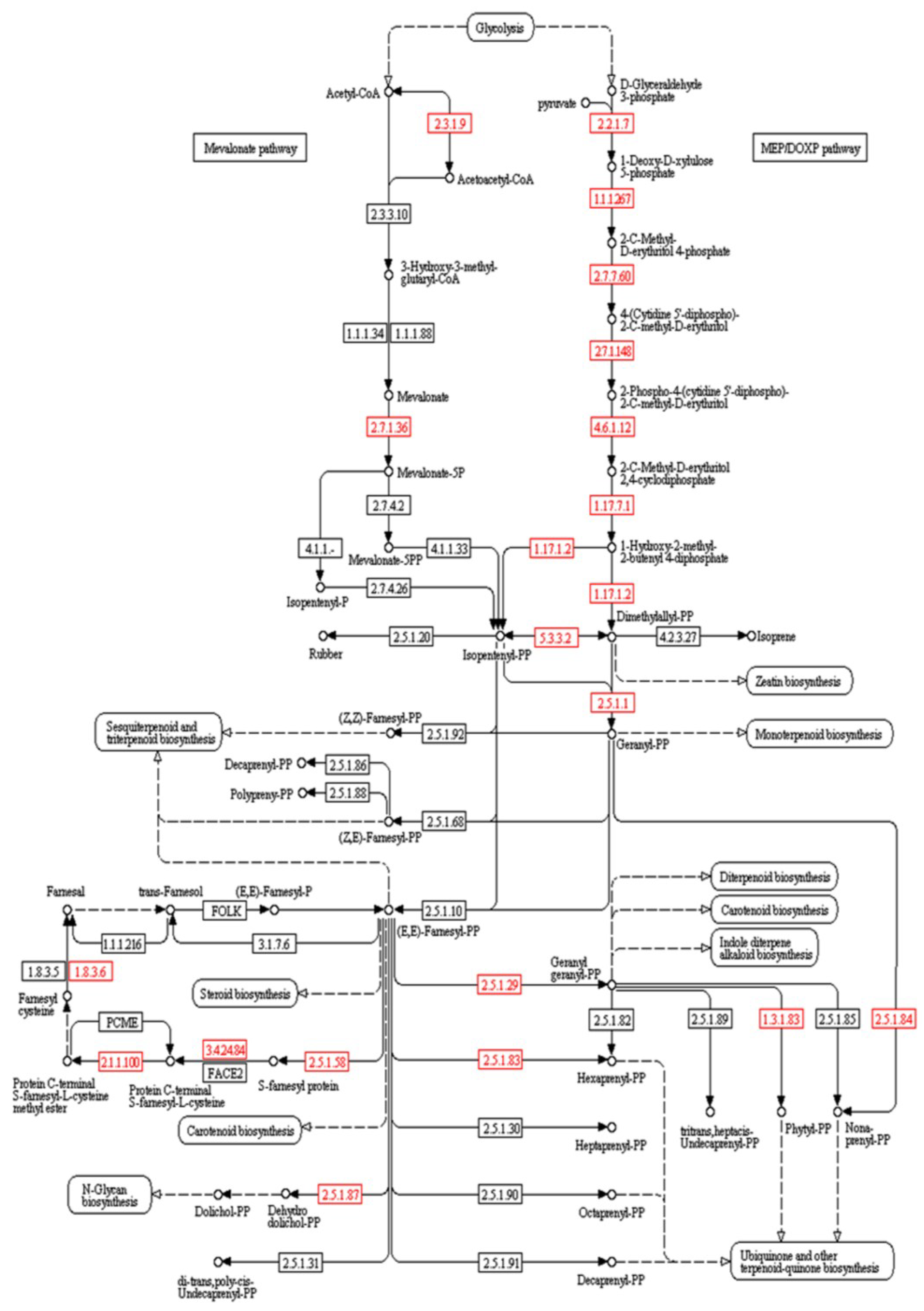{kind=link}
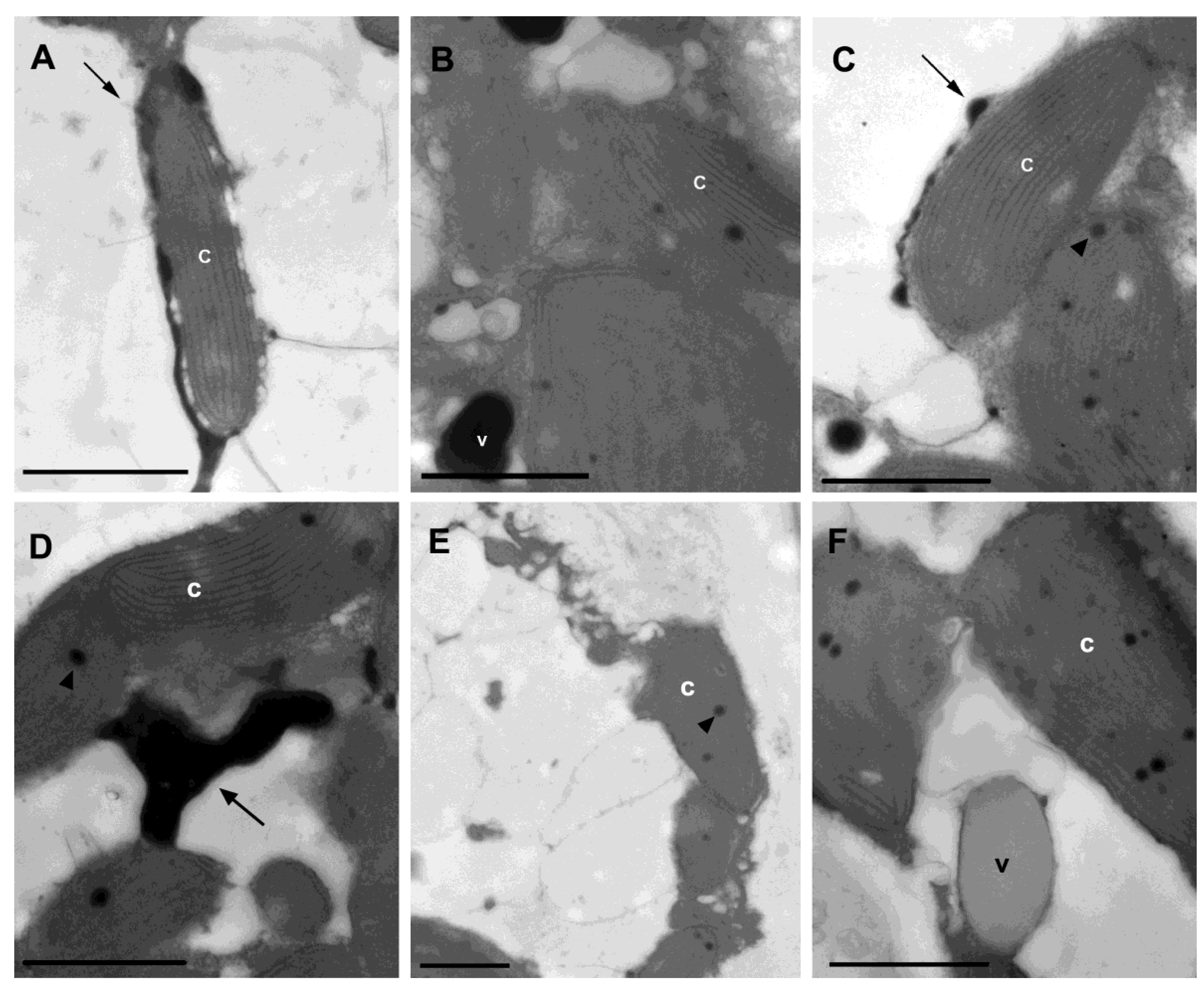{kind=link}
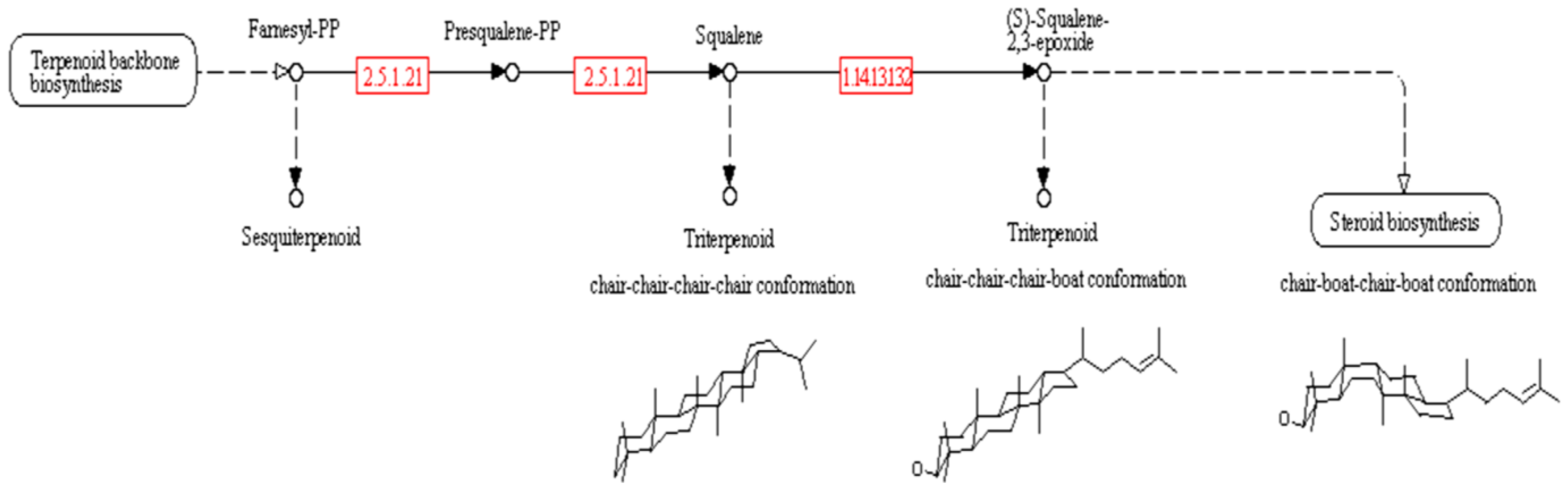{kind=link}
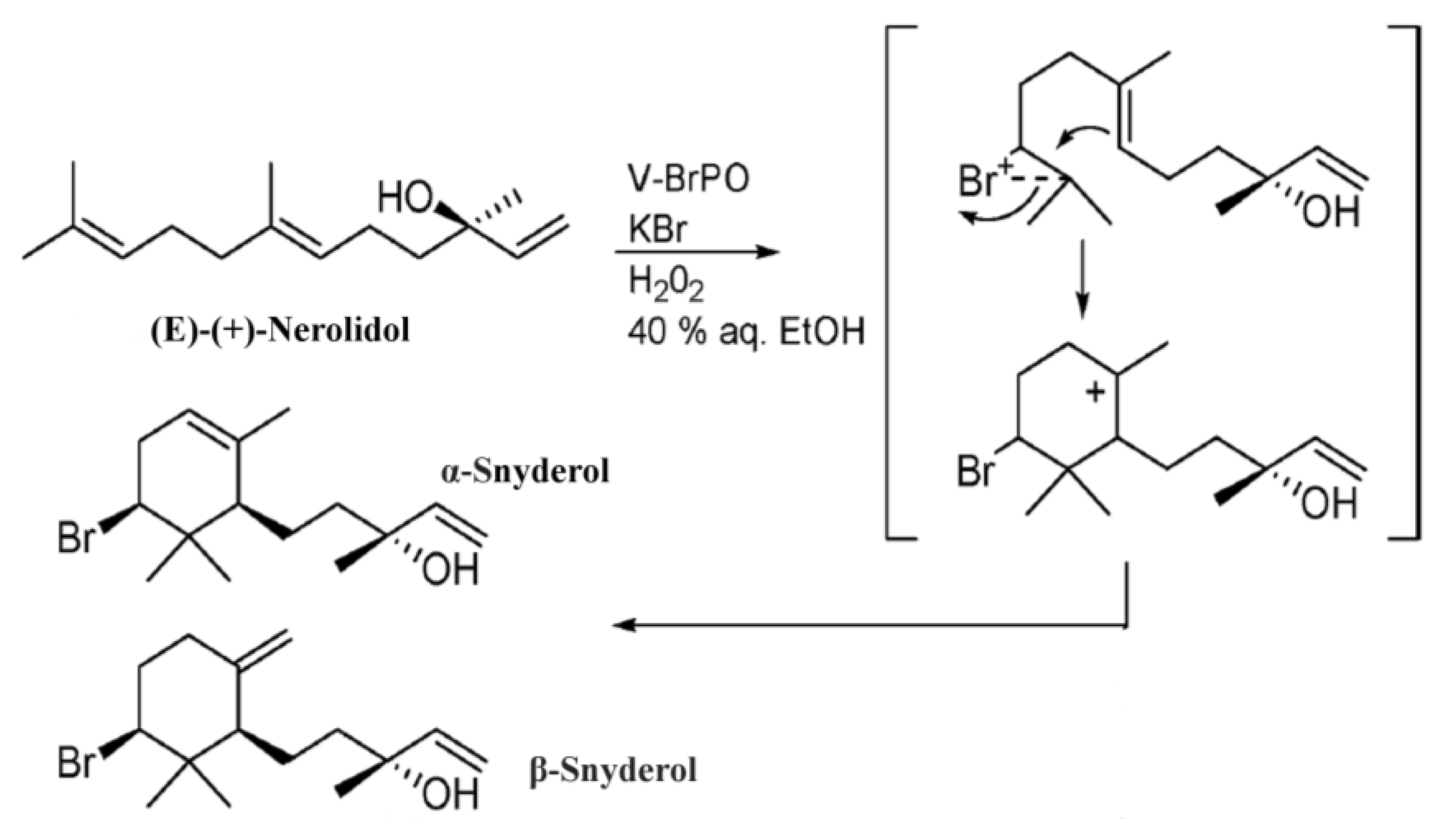{kind=link}
| Sample | Number of Sequences | Total Nucleotides (bp) | Average Size (bp) ± SD |
|---|---|---|---|
| Light | 5,825,960 | 749,118,981 | 128.0 ± 30 |
| Dark | 70,560 | 15,816,057 | 224.0 ± 45 |
| Assembled sequences | 54,255 | 19,504,276 | 359.5 ± 589 |


| Gene Name | EC Number | Blast e-Value | Similarity | Gene Product | Terpene Class | Role | Reference |
|---|---|---|---|---|---|---|---|
| (3R)-linalool synthase | 4.2.3.26 | 3e-07 | 68% | Linalool | Monoterpene | Defense (ER); antibiotic, antifungal anticonvulsant, antitumor (BP) | [33,34,35,38,39] |
| (+)-trans-carveol dehydrogenase | 1.1.1.275 | 2e-08 | 54% | (+)-(S)-carvone | Monoterpene | Anti-herbivore, antifungal (ER); anticonvulsant, antibiotic, cytotoxic, anti-sprouting agent in potatoes (BP). | [40,41,42,43] |
| (+)-pulegone reductase | 1.3.1.81 | 3e-39 | 52% | (+)-pulegone | Monoterpene | Defense (ER); analgesic, antibacterial, antifungal, insecticide, acaricidal (BP) | [38,39,44,45,46,47] |
| (−)-isopiperitenone reductase | 1.3.1.82 | 1e-04 | 56% | (−)-isopiperitenone (intermediate compound to (−)-menthone) | Monoterpene | Defense (ER); acaricidal, antibiotic (BP) | [38,39,47,48] |
| secologanin synthase | 1.3.3.9 | 2e-16 | 52% | Secologanin | Monoterpene | Precursor to indole alkaloids; antimicrobial (BP) | [48,49] |
| farnesyl-diphosphate farnesyltransferase | 2.5.1.21 | 3e-12 | 64% | Squalene | Triterpene | Precursor to triterpene | [50] |
| squalene monooxygenase | 1.14.13.132 | 3e-96 | 69% | (S)-squalene-2,3-epoxide | Triterpene | Precursor to triterpene | [50] |
| squalene-hopene/tetraprenyl-beta-curcumene cyclase | 4.2.1.129 | 7e-04 | 62% | hopan-22-ol | Triterpene | Precursor to triterpene with chair-chair-chair-chair conformation | [51] |
| lupeol synthase 1 | 5.4.99.41 | 2e-04 | 41% | lupeol,β-amyrin | Triterpene | Antibacterial, anti-fungal, anti-inflammatory, antineoplastic, antihypertensive, antiurolithiatic (BP) | [52,53,54] |
| nerolidol synthase | 4.2.3.48 | 9e-04 | 62% | Nerolidol | Sesquiterpene | Precursor to α- and β-snyderols | [20,45] |
| alpha-bisabolene synthase | 4.2.3.38 | 6e-04 | 43% | (E)-alpha-bisabolene | Sesquiterpene | Precursor to (−)-elatol and caespitol; Defense (ER); antileishmanial, anti-trypanosomal, antibiotic, anti-tumor (BP) | [6,7,21,22,26,55,56,57] |
| germacrene-A synthase | 4.2.3.23 | 6e-20 | 43% | (+)-(R)-gemacrene A | Sesquiterpene | Precursor to germacrene-type sesquiterpenes | [58,59] |
| germacrene A oxidase | 1.1.1.314 | 3e-9 | 52% | germacra-1(10),4,11(13)-trien-12-oate | Sesquiterpene | Precursor to germacrene-type sesquiterpenes | [58,59] |
| aristolochene synthase | 4.2.3.9 | 5e-04 | 44% | Aristolochene | Sesquiterpene | Precursor to germacrene-type sesquiterpenes | [58,59] |
| 5-epiaristolochene 1,3-dihydroxylase | 1.14.13.119 | 3e-20 | 53% | Capsidiol | Sesquiterpene | Plant defense (ER); Antibiotic, prostaglandin inhibitor (BP) | [52,60,61,62] |
| pentalenene synthase | 4.2.3.7 | 1e-07 | 42% | Pentalenene | Sesquiterpene | Precursor to humulene-type sesquiterpene | [63] |
| (+)-delta-cadinene synthase | 4.2.3.13 | 3e-05 | 48% | Precursor to (−)-δ-cadinene and (+)-α-cadinol | Sesquiterpene | Plant defense (ER), antibiotic (BP) | [60,61] |
| zerumbone synthase | 1.1.1.326 | 5e-08 | 43% | Zerumbone | Sesquiterpene | Antitumor, anti Alzheimer’s disease (BP) | [52,62] |
| gibberellin 20-oxidase | 1.14.11.12 | 2e-12 | 43% | gibberellin 44 | Diterpene | Endogenous growth regulators (ER) | [53,54] |
| gibberellin 2-oxidase | 1.14.11.13 | 7e-11 | 45% | 2beta-hydroxygibberellin 1 | Diterpene | Endogenous growth regulators (ER) | [53,54] |
| abietadienol/abietadienal oxidase | 1.14.13.109 | 1e-13 | 46% | diterpene acids | Diterpene | Intermediate to diverse diterpene skeletons | [64] |

3. Discussion
3.1. Monoterpene Synthase Genes
3.2. Triterpene Synthase Genes
3.3. Sesquiterpene Synthase Genes

3.4. Diterpene Synthase Genes
4. Experimental Section
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hay, M.E. Marine chemical ecology: Chemical signals and cues structure marine populations, communities, and ecosystems. Ann. Rev. Mar. Sci. 2009, 1, 193–212. [Google Scholar] [CrossRef] [PubMed]
- Amsler, C.D. Chemical ecology of seaweeds. In Seaweed Biology; Wiencke, C., Bischof, K., Eds.; Ecological Studies; Springer (Berlin Heidelberg): Berlin, Germany, 2012; Volume 219, pp. 177–188. [Google Scholar]
- Rasher, D.B.; Hay, M.E. Chemically rich seaweeds poison corals when not controlled by herbivores. Proc. Natl. Acad. Sci. USA 2010, 107, 9683–9688. [Google Scholar] [CrossRef] [PubMed]
- Irie, T.; Suzuki, M.; Masamune, T. Laurencin, a constituent from Laurencia species. Tetrahedron Lett. 1965, 6, 1091–1099. [Google Scholar] [CrossRef]
- Liang, Y.; Li, X.-M.; Li, C.-S.; Sun, H.; Wang, B.-G. Laurane-, cyclolaurane-, and cuparane-type sesquiterpenes from the marine red alga Laurencia okamurai. Nat. Prod. Commun. 2014, 9, 323–324. [Google Scholar] [PubMed]
- Pereira, R.C.; da Gama, B.AP.; Teixeira, V.L.; Yoneshigue-Valentin, Y. Ecological roles of natural products of the Brazilian red seaweed Laurencia obtusa. Braz. J. Biol. 2003, 63, 665–72. [Google Scholar] [CrossRef] [PubMed]
- Da Gama, B.A; Pereira, R.C.; Soares, A.R.; Teixeira, V.L.; Yoneshigue-Valentin, Y. Is the mussel test a good indicator of antifouling activity? A comparison between laboratory and field assays. Biofouling 2003, 19 (Suppl.), 161–169. [Google Scholar]
- Sudatti, D.B.; Rodrigues, S.V.; Pereira, R.C. Quantitative GC-ECD analysis of halogenated metabolites: Determination of surface and within-thallus elatol of Laurencia obtusa. J. Chem. Ecol. 2006, 32, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.S.; Sudatti, D.B.; Fujii, M.T.; Rodrigues, S.V.; Pereira, R.C. Inter- and intrapopulation variation in the defensive chemistry of the red seaweed Laurencia dendroidea (Ceramiales, Rhodophyta). Phycologia 2013, 52, 130–136. [Google Scholar] [CrossRef]
- Sudatti, D.B.; Fujii, M.T.; Rodrigues, S.V.; Turra, A.; Pereira, R.C. Effects of abiotic factors on growth and chemical defenses in cultivated clones of Laurencia dendroidea J. Agardh (Ceramiales, Rhodophyta). Mar. Biol. 2011, 158, 1439–1446. [Google Scholar] [CrossRef]
- Sakemi, S.; Higa, T.; Jefford, C.W.; Bernardinelli, G. Venustatriol. A new, anti-viral, triterpene tetracyclic ether from Laurencia venusta. Tetrahedron Lett. 1986, 27, 4287–4290. [Google Scholar] [CrossRef]
- Vairappan, C.S.; Daitoh, M.; Suzuki, M.; Abe, T.; Masuda, M. Antibacterial halogenated metabolites from the Malaysian Laurencia species. Phytochemistry 2001, 58, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Vairappan, C.S.; Kawamoto, T.; Miwa, H.; Suzuki, M. Potent antibacterial activity of halogenated compounds against antibiotic-resistant bacteria. Planta Med. 2004, 70, 1087–1090. [Google Scholar] [CrossRef] [PubMed]
- Topcu, G.; Aydogmus, Z.; Imre, S.; Gören, A.C.; Pezzuto, J.M.; Clement, J.A.; Kingston, D.G.I. Brominated sesquiterpenes from the red alga Laurencia obtusa. J. Nat. Prod. 2003, 66, 1505–1508. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, A.O.; Veiga-Santos, P.; Ueda-Nakamura, T.; Filho, B.P.D.; Sudatti, D.B.; Bianco, E.M.; Pereira, R.C.; Nakamura, C.V. Effect of elatol, isolated from red seaweed Laurencia dendroidea, on Leishmania amazonensis. Mar. Drugs 2010, 8, 2733–2743. [Google Scholar] [CrossRef] [PubMed]
- Veiga-Santos, P.; Pelizzaro-Rocha, K.J.; Santos, A.O.; Ueda-Nakamura, T.; Dias Filho, B.P.; Silva, S.O.; Sudatti, D.B.; Bianco, E.M.; Pereira, R.C.; Nakamura, C.V. In vitro anti-trypanosomal activity of elatol isolated from red seaweed Laurencia dendroidea. Parasitology 2010, 137, 1661–1670. [Google Scholar] [CrossRef] [PubMed]
- Chatter, R.; Ben Othman, R.; Rabhi, S.; Kladi, M.; Tarhouni, S.; Vagias, C.; Roussis, V.; Guizani-Tabbane, L.; Kharrat, R. In vivo and in vitro anti-inflammatory activity of neorogioltriol, a new diterpene extracted from the red algae Laurencia glandulifera. Mar. Drugs 2011, 9, 1293–1306. [Google Scholar] [CrossRef] [PubMed]
- Kladi, M.; Xenaki, H.; Vagias, C.; Papazafiri, P.; Roussis, V. New cytotoxic sesquiterpenes from the red algae Laurencia obtusa and Laurencia microcladia. Tetrahedron 2006, 62, 182–189. [Google Scholar] [CrossRef]
- Kim, M.-M.; Mendis, E.; Kim, S.-K. Laurencia okamurai extract containing laurinterol induces apoptosis in melanoma cells. J. Med. Food 2008, 11, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Lhullier, C.; Falkenberg, M.; Ioannou, E.; Quesada, A.; Papazafiri, P.; Horta, P.A.; Schenkel, E.P.; Vagias, C.; Roussis, V. Cytotoxic halogenated metabolites from the Brazilian red alga Laurencia catarinensis. J. Nat. Prod. 2010, 73, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Da Gama, B.A.P.; Pereira, R.C.; Carvalho, A.G.V; Coutinho, R.; Yoneshigue-Valentin, Y. The effects of seaweed secondary metabolites on biofouling. Biofouling 2002, 18, 13–20. [Google Scholar] [CrossRef]
- White, D.E.; Stewart, I.C.; Grubbs, R.H.; Stoltz, B.M. The catalytic asymmetric total synthesis of elatol. J. Am. Chem. Soc. 2008, 130, 810–811. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.C.; Costa-Lotufo, L.V. Bioprospecting for bioactives from seaweeds: Potential, obstacles and alternatives. Rev. Bras. Farmacogn. 2012, 22, 894–905. [Google Scholar] [CrossRef]
- Ignea, C.; Cvetkovic, I.; Loupassaki, S.; Kefalas, P.; Johnson, C.B.; Kampranis, S.C.; Makris, A.M. Improving yeast strains using recyclable integration cassettes, for the production of plant terpenoids. Microb. Cell Fact. 2011, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- Martin, V.J.J.; Pitera, D.J.; Withers, S.T.; Newman, J.D.; Keasling, J.D. Engineering a mevalonate pathway in Escherichia coli for production of terpenoids. Nat. Biotechnol. 2003, 21, 796–802. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, L.S.; Gregoracci, G.B.; Silva, G.G.Z.; Salgado, L.T.; Filho, G.A.; Alves-Ferreira, M.; Pereira, R.C.; Thompson, F.L. Transcriptomic analysis of the red seaweed Laurencia dendroidea (Florideophyceae, Rhodophyta) and its microbiome. BMC Genomics 2012, 13, 487. [Google Scholar] [CrossRef] [PubMed]
- Lohr, M.; Schwender, J.; Polle, J.E.W. Isoprenoid biosynthesis in eukaryotic phototrophs: A spotlight on algae. Plant Sci. 2012, 185–186, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Fenical, W. Halogenation in the Rhodophyta, a Review. J. Phycol. 1975, 11, 245–259. [Google Scholar]
- Cabrita, M.T.; Vale, C.; Rauter, A.P. Halogenated compounds from marine algae. Mar. Drugs 2010, 8, 2301–2317. [Google Scholar] [CrossRef] [PubMed]
- Young, D.N.; Howard, B.M.; Fenical, W. Subcellular localization of brominated secondarymetabolites in the red alga Laurencia snyderae. J. Phycol. 1980, 16, 182–185. [Google Scholar] [CrossRef]
- Salgado, L.T.; Viana, N.B.; Andrade, L.R.; Leal, R.N.; da Gama, B.A.P.; Attias, M.; Pereira, R.C.; Amado Filho, G.M. Intra-cellular storage, transport and exocytosis of halogenated compounds in marine red alga Laurencia obtusa. J. Struct. Biol. 2008, 162, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Reis, V.M.; Oliveira, L.S.; Passos, R.M.F.; Viana, N.B.; Mermelstein, C.; Sant’anna, C.; Pereira, R.C.; Paradas, W.C.; Thompson, F.L.; Amado-Filho, G.M.; et al. Traffic of secondary metabolites to cell surface in the red alga Laurencia dendroidea depends on a two-step transport by the cytoskeleton. PLoS One 2013, 8, e63929. [Google Scholar] [CrossRef]
- Paradas, W.C.; Salgado, L.T.; Sudatti, D.B.; Crapez, M.A; Fujii, M.T.; Coutinho, R.; Pereira, R.C.; Amado Filho, G.M. Induction of halogenated vesicle transport in cells of the red seaweed Laurencia obtusa. Biofouling 2010, 26, 277–286. [Google Scholar] [CrossRef]
- Nakamura, Y.; Sasaki, N.; Kobayashi, M.; Ojima, N.; Yasuike, M.; Shigenobu, Y.; Satomi, M.; Fukuma, Y.; Shiwaku, K.; Tsujimoto, A.; et al. The first symbiont-free genome sequence of marine red alga, Susabi-nori (Pyropia yezoensis). PLoS One 2013, 8, e57122. [Google Scholar] [CrossRef] [PubMed]
- Collén, J.; Porcel, B.; Carré, W.; Ball, S.G.; Chaparro, C.; Tonon, T.; Barbeyron, T.; Michel, G.; Noel, B.; Valentin, K.; et al. Genome structure and metabolic features in the red seaweed Chondrus crispus shed light on evolution of the Archaeplastida. Proc. Natl. Acad. Sci. USA 2013, 110, 5247–5252. [Google Scholar] [CrossRef] [PubMed]
- Kapraun, D.F. Nuclear DNA content estimates in multicellular green, red and brown algae: Phylogenetic considerations. Ann. Bot. 2005, 95, 7–44. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. Kegg Pathway Database: Terpenoid backbone biosynthesis-reference pathway. Available online: http://www.genome.jp/kegg-bin/show_pathway?map00900 (accessed on 15 September 2014).
- De Sousa, D.P.; Nóbrega, F.F.F.; de Lima, M.R.V; de Almeida, R.N. Pharmacological activity of (R)-(+)-pulegone, a chemical constituent of essential oils. Z. Naturforsch. C. 2011, 66, 353–359. [Google Scholar]
- Arruda, T.A.; Antunes, R.M.P.; Catão, R.M.R.; Lima, E.O.; Sousa, D.P.; Nunes, X.P.; Pereira, M.S.V.; Barbosa-Filho, J.M.; da Cunha, E.V.L. Preliminary study of the antimicrobial activity of Mentha x villosa Hudson essential oil, rotundifolone and its analogues. Rev. Bras. Farmacogn. 2006, 16, 307–311. [Google Scholar] [CrossRef]
- Khan, A.; Ahmad, A.; Manzoor, N.; Khan, L.A. Antifungal activities of Ocimum sanctum essential oil and its lead molecules. Nat. Prod. Commun. 2010, 5, 345–349. [Google Scholar] [PubMed]
- Elisabetsky, E.; Brum, L.F.; Souza, D.O. Anticonvulsant properties of linalool in glutamate-related seizure models. Phytomedicine 1999, 6, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.W.; Crock, J.; Lu, S.; Croteau, R.; Chen, X.Y. (3R)-Linalool synthase from Artemisia annua L.: cDNA isolation, characterization, and wound induction. Arch. Biochem. Biophys. 1999, 372, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, S.; Hosokawa-Shinonaga, Y.; Tamaoki, D.; Yamada, S.; Akimitsu, K.; Gomi, K. Jasmonate induction of the monoterpene linalool confers resistance to rice bacterial blight and its biosynthesis is regulated by JAZ protein in rice. Plant. Cell Environ. 2014, 37, 451–461. [Google Scholar] [CrossRef] [PubMed]
- De Sousa, D.P.; de Farias Nóbrega, F.F.; de Almeida, R.N. Influence of the chirality of (R)-(−)- and (S)-(+)-carvone in the central nervous system: A comparative study. Chirality 2007, 19, 264–268. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, C.; da Fonseca, M. Carvone: Why and how should one bother to produce this terpene. Food Chem. 2006, 95, 413–422. [Google Scholar] [CrossRef]
- Frank, T.; Biert, K.; Speiser, B. Feeding deterrent effect of carvone, a compound from caraway seeds, on the slug Arion lusitanicus. Ann. Appl. Biol. 2002, 141, 93–100. [Google Scholar] [CrossRef]
- Smid, E.J.; de Witte, Y.; Gorris, L.G.M. Secondary plant metabolites as control agents of postharvest Penicillium rot on tulip bulbs. Postharvest Biol. Technol. 1995, 6, 303–312. [Google Scholar] [CrossRef]
- Franzios, G.; Mirotsou, M.; Hatziapostolou, E.; Kral, J.; Scouras, Z.G.; Mavragani-Tsipidou, P. Insecticidal and genotoxic activities of mint essential oils. J. Agric. Food Chem. 1997, 45, 2690–2694. [Google Scholar] [CrossRef]
- McGarvey, D.J.; Croteau, R. Terpenoid metabolism. Plant Cell 1995, 7, 1015–1026. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2012, 29, 144–222. [Google Scholar] [CrossRef] [PubMed]
- Pearson, W.R. An introduction to sequence similarity (“homology”) searching. In Curr. Protoc. Bioinform.; 2013. Available online: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3820096/pdf/nihms519883.pdf (accessed on 15 September 2014). [Google Scholar]
- Vairappan, C.S.; Anangdan, S.P.; Tan, K.L.; Matsunaga, S. Role of secondary metabolites as defense chemicals against ice-ice disease bacteria in biofouler at carrageenophyte farms. J. Appl. Phycol. 2009, 22, 305–311. [Google Scholar] [CrossRef]
- Vairappan, C.S. Potent antibacterial activity of halogenated metabolites from Malaysian red algae, Laurencia majuscula (Rhodomelaceae, Ceramiales). Biomol. Eng. 2003, 20, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Vairappan, C.S.; Kamada, T.; Lee, W.-W.; Jeon, Y.-J. Anti-inflammatory activity of halogenated secondary metabolites of Laurencia snackeyi (Weber-van Bosse) Masuda in LPS-stimulated RAW 264.7 macrophages. J. Appl. Phycol. 2013, 25, 1805–1813. [Google Scholar] [CrossRef]
- Phillips, D.R.; Rasbery, J.M.; Bartel, B.; Matsuda, S.P. Biosynthetic diversity in plant triterpene cyclization. Curr. Opin. Plant Biol. 2006, 9, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Nakano, S.-I.; Ohashi, S.; Hoshino, T. Squalene-hopene cyclase: Insight into the role of the methyl group on the squalene backbone upon the polycyclization cascade. Enzymatic cyclization products of squalene analogs lacking a 26-methyl group and possessing a methyl group at C7 or C11. Org. Biomol. Chem. 2004, 2, 2012–2022. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Raouf, N.; Al-Enazi, N.M.; Al-Homaidan, A.A.; Ibraheem, I.B. M.; Al-Othman, M.R.; Hatamleh, A.A. Antibacterial β-amyrin isolated from Laurencia microcladia. Arab. J. Chem. 2013, 8, 32–37. [Google Scholar] [CrossRef]
- Chang, M.-Y.; Shen, Y.-L. Linalool exhibits cytotoxic effects by activating antitumor immunity. Molecules 2014, 19, 6694–6706. [Google Scholar] [CrossRef] [PubMed]
- Güven, K.C.; Coban, B.; Sezik, E.; Erdugan, H.; Kaleağasıoğlu, F. Alkaloids of marine macroalgae. In Natural Products. Phytochemistry, Botany and Metabolism of Alkaloids, Phenolics and Terpenes; Ramawat, K.G., Mérillon, J.-M., Eds.; Springer (Berlin Heidelberg): Berlin, Germany, 2013; pp. 25–37. [Google Scholar]
- Vázquez, L.H.; Palazon, J.; Navarro-Ocaña, A. The pentacyclic triterpenes α-, β-amyrins: A review of sources and biological activities. In Phytochemicals-A Global Perspective of Their Role in Nutrition and Health; Rao, V., Ed.; InTech: Rijeka, Croatia, 2012. [Google Scholar]
- Gallo, M.; Sarachine, M. Biological activities of lupeol. Int. J. Biomed. Pharm. Sci. 2009, 3, 46–66. [Google Scholar]
- Carter-Franklin, J.N.; Butler, A. Vanadium bromoperoxidase-catalyzed biosynthesis of halogenated marine natural products. J. Am. Chem. Soc. 2004, 126, 15060–15066. [Google Scholar] [CrossRef] [PubMed]
- Cane, D.E. Enzymic formation of sesquiterpenes. Chem. Rev. 1990, 90, 1089–1103. [Google Scholar] [CrossRef]
- Yu, X.-Q.; He, W.-F.; Liu, D.-Q.; Feng, M.-T.; Fang, Y.; Wang, B.; Feng, L.-H.; Guo, Y.-W.; Mao, S.-C. A seco-laurane sesquiterpene and related laurane derivatives from the red alga Laurencia okamurai Yamada. Phytochemistry 2014, 103, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. Kegg Pathway Database: Sesquiterpenoid and triterpenoid biosynthesis—reference pathway. Available online: http://www.genome.jp/kegg-bin/show_pathway?map00909 (accessed on 15 September 2014).
- Leivar, P.; González, V.M.; Castel, S.; Trelease, R.N.; López-Iglesias, C.; Arró, M.; Boronat, A.; Campos, N.; Ferrer, A.; Fernàndez-Busquets, X. Subcellular localization of Arabidopsis 3-hydroxy-3-methylglutaryl-coenzyme A reductase. Plant Physiol. 2005, 137, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Brooker, J.D.; Russell, D.W. Subcellular localization of 3-hydroxy-3-methylglutaryl coenzyme A reductase in Pisum sativum seedlings. Arch. Biochem. Biophys. 1975, 167, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Arebalo, R.E.; Mitchell, E.D. Cellular distribution of 3-hydroxy-3-methylglutaryl coenzyme a reductase and mevalonate kinase in leaves of Nepeta cataria. Phytochemistry 1984, 23, 13–18. [Google Scholar] [CrossRef]
- Kuhn, D.N.; Knauf, M.; Stumpf, P.K. Subcellular localization of acetyl-CoA synthetase in leaf protoplasts of Spinacia oleracea. Arch. Biochem. Biophys. 1981, 209, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Lombard, J.; Moreira, D. Origins and early evolution of the mevalonate pathway of isoprenoid biosynthesis in the three domains of life. Mol. Biol. Evol. 2011, 28, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Lange, B.M.; Rujan, T.; Martin, W.; Croteau, R. Isoprenoid biosynthesis: The evolution of two ancient and distinct pathways across genomes. Proc. Natl. Acad. Sci. USA 2000, 97, 13172–13177. [Google Scholar] [CrossRef] [PubMed]
- Laule, O.; Fürholz, A.; Chang, H.-S.; Zhu, T.; Wang, X.; Heifetz, P.B.; Gruissem, W.; Lange, M. Crosstalk between cytosolic and plastidial pathways of isoprenoid biosynthesis in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2003, 100, 6866–6871. [Google Scholar] [CrossRef] [PubMed]
- Adam, K.P.; Thiel, R.; Zapp, J. Incorporation of 1-[1-(13)C]Deoxy-d-xylulose in chamomile sesquiterpenes. Arch. Biochem. Biophys. 1999, 369, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Tholl, D.; Bohlmann, J.; Pichersky, E. The family of terpene synthases in plants: A mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom. Plant J. 2011, 66, 212–229. [Google Scholar] [CrossRef] [PubMed]
- Degenhardt, J.; Köllner, T.G.; Gershenzon, J. Monoterpene and sesquiterpene synthases and the origin of terpene skeletal diversity in plants. Phytochemistry 2009, 70, 1621–1637. [Google Scholar] [CrossRef] [PubMed]
- Weinberger, F.; Coquempot, B.; Forner, S.; Morin, P.; Kloareg, B.; Potin, P. Different regulation of haloperoxidation during agar oligosaccharide-activated defence mechanisms in two related red algae, Gracilaria sp. and Gracilaria chilensis. J. Exp. Bot. 2007, 58, 4365–4372. [Google Scholar] [CrossRef] [PubMed]
- Payo, D.A.; Colo, J.; Calumpong, H.; de Clerck, O. Variability of non-polar secondary metabolites in the red alga Portieria. Mar. Drugs 2011, 9, 2438–2468. [Google Scholar] [CrossRef] [PubMed]
- Arnold, T.M.; Targett, N.M.; Tanner, C.E.; Hatch, W.I.; Ferrari, K.E. Evidence for methyl jasmonate-induced phlorotannin production in Fucus vesiculosus (Phaeophyceae). J. Phycol. 2001, 37, 1026–1029. [Google Scholar] [CrossRef]
- Collén, J.; Hervé, C.; Guisle-Marsollier, I.; Léger, J.J.; Boyen, C. Expression profiling of Chondrus crispus (Rhodophyta) after exposure to methyl jasmonate. J. Exp. Bot. 2006, 57, 3869–3881. [Google Scholar] [CrossRef] [PubMed]
- Salehi, P.; Sonboli, A.; Asghari, B. Chemical composition of the essential oil of Stachys acerosa and its antibacterial and antioxidant activities. Chem. Nat. Compd. 2007, 43, 339–341. [Google Scholar] [CrossRef]
- Eisner, T.; McCormick, K.D.; Sakaino, M.; Eisner, M.; Smedley, S.R.; Aneshansley, D.J.; Deyrup, M.; Myers, R.L.; Meinwald, J. Chemical defense of a rare mint plant. Chemoecology 1990, 1, 30–37. [Google Scholar] [CrossRef]
- Banchio, E.; Zygadlo, J.; Valladares, G.R. Effects of mechanical wounding on essential oil composition and emission of volatiles from Minthostachys mollis. J. Chem. Ecol. 2005, 31, 719–727. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Ramos, I.; Castañera, P. Acaricidal activity of natural monoterpenes on Tyrophagus putrescentiae (Schrank), a mite of stored food. J. Stored Prod. Res. 2000, 37, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Vimal, M.; Vijaya, P.P.; Mumtaj, P.; Farhath, M.S. Antibacterial activity of selected compounds of essential oils from indigenous plants. J. Chem. Pharm. Res. 2013, 5, 248–253. [Google Scholar]
- Carter, G.T.; Rinehart, K.L.; Li, L.H.; Kuentzel, S.L.; Connor, J.L. Brominated indoles from Laurencia brongniartii. Tetrahedron Lett. 1978, 19, 4479–4482. [Google Scholar] [CrossRef]
- Güven, K.C.; Percot, A.; Sezik, E. Alkaloids in marine algae. Mar. Drugs 2010, 8, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Manrı́quez, C.P.; Souto, M.L.; Gavı́n, J.A.; Norte, M.; Fernández, J.J. Several new squalene-derived triterpenes from Laurencia. Tetrahedron 2001, 57, 3117–3123. [Google Scholar] [CrossRef]
- Pacheco, F.C.; Villa-Pulgarin, J.A.; Mollinedo, F.; Martín, M.N.; Fernández, J.J.; Daranas, A.H. New polyether triterpenoids from Laurencia viridis and their biological evaluation. Mar. Drugs 2011, 9, 2220–2235. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, Y.; Suzuki, M.; Masuda, M. Enshuol, a novel squalene-derived pentacyclic triterpene alcohol from a new species of the red algal genus Laurencia. Chem. Lett. 1995, 1043–1044. [Google Scholar] [CrossRef]
- Arruda, D.C.; D’Alexandri, F.L.; Katzin, A.M.; Uliana, S.R.B. Antileishmanial activity of the terpene nerolidol. Antimicrob. Agents Chemother. 2005, 49, 1679–1687. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.P.N.; Oliveira, G.L.S.; de Carvalho, R.B.F.; de Sousa, D.P.; Freitas, R.M.; Pinto, P.L.S.; de Moraes, J. Antischistosomal activity of the terpene nerolidol. Molecules 2014, 19, 3793–3803. [Google Scholar] [CrossRef] [PubMed]
- Klopell, F.C.; Lemos, M.; Sousa, J.P.B.; Comunello, E.; Maistro, E.L.; Bastos, J.K.; de Andrade, S.F. Nerolidol, an antiulcer constituent from the essential oil of Baccharis dracunculifolia DC (Asteraceae). Z. Naturforsch. C. 2007, 62, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Schever, P. Marine Natural Products V1: Chemical and Biological Perspectives; Academic Press: New York, NY, USA, 2012; Volume 1, p. 318. [Google Scholar]
- Fujii, M.T.; Cassano, V.; Senties, A.; Diaz-Larrea, J.; Machin-Sanchez, M.; Candelaria Gil-Rodriguez, M. Comparative analysis of the corps en cerise in several species of Laurencia (Ceramiales, Rhodophyta) from the Atlantic Ocean. Rev. Bras. Farm. J. Pharmacogn. 2012, 22, 795. [Google Scholar] [CrossRef]
- Davyt, D.; Fernandez, R.; Suescun, L.; Mombrú, A.W.; Saldaña, J.; Domínguez, L.; Fujii, M.T.; Manta, E. Bisabolanes from the red alga Laurencia scoparia. J. Nat. Prod. 2006, 69, 1113–1116. [Google Scholar] [CrossRef] [PubMed]
- Rising, K.A.; Starks, C.M.; Noel, J.P.; Chappell, J. Demonstration of germacrene A as an intermediate in 5-epi-aristolochene synthase catalysis. J. Am. Chem. Soc. 2000, 122, 1861–1866. [Google Scholar] [CrossRef]
- Brennan, M.R.; Erickson, K.L. Austradiol acetate and austradiol diacetate, 4,6-dihydroxy-(+)-selinane derivatives from an Australian Laurencia sp. J. Org. Chem. 1982, 47, 3917–3921. [Google Scholar] [CrossRef]
- Guellaa, G.; Skropetaa, D.; Mancinia, L.; Pietrab, F. The First 6,8-Cycloeudesmane sesquiterpene from a amrine organism: The red seaweed Laurencia microcladia from the Baia di Calenzana, Elba Island. Z. Naturforsch 2001, 57B, 1147–1151. [Google Scholar]
- Guella, G.; Skropeta, D.; Mancini, I.; Pietra, F. Calenzanane sesquiterpenes from the red seaweed Laurencia microcladia from the Bay of Calenzana, Elba Island: Acid-Catalyzed stereospecific conversion of calenzanol into indene- and guaiazulene-type sesquiterpenes. Chemistry 2003, 9, 5770–5777. [Google Scholar] [CrossRef] [PubMed]
- Bailey, J.A.; Burden, R.S.; Vincent, G.G. Capsidiol: An antifungal compound produced in Nicotiana tabacum and Nicotiana clevelandii following infection with tobacco necrosis virus. Phytochemistry 1975, 14, 597. [Google Scholar] [CrossRef]
- Bohlmann, J.; Stauber, E.J.; Krock, B.; Oldham, N.J.; Gershenzon, J.; Baldwin, I.T. Gene expression of 5-epi-aristolochene synthase and formation of capsidiol in roots of Nicotiana attenuata and N. sylvestris. Phytochemistry 2002, 60, 109–116. [Google Scholar] [CrossRef] [PubMed]
- De Marino, S.; Borbone, N.; Gala, F.; Zollo, F.; Fico, G.; Pagiotti, R.; Iorizzi, M. New constituents of sweet Capsicum annuum L. fruits and evaluation of their biological activity. J. Agric. Food Chem. 2006, 54, 7508–7516. [Google Scholar] [CrossRef] [PubMed]
- Nasiri, A.; Holth, A.; Björk, L. Effects of the sesquiterpene capsidiol on isolated guinea-pig ileum and trachea, and on prostaglandin synthesis in vitro. Planta Med. 1993, 59, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Kelecom, A. Synthesis of marine natural products in Brazil. J. Braz. Chem. Soc. 1998, 9, 101–118. [Google Scholar] [CrossRef]
- Qiao, Y.-Y.; Ji, N.-Y.; Wen, W.; Yin, X.-L.; Xue, Q.-Z. A new epoxy-cadinane sesquiterpene from the marine brown alga Dictyopteris divaricata. Mar. Drugs 2009, 7, 600–604. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Y.; Chen, Y.; Heinstein, P.; Davisson, V.J. Cloning, expression, and characterization of (+)-delta-cadinene synthase: A catalyst for cotton phytoalexin biosynthesis. Arch. Biochem. Biophys. 1995, 324, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Pérez-López, A.; Cirio, A.T.; Rivas-Galindo, V.M.; Aranda, R.S.; de Torres, N.W. Activity against Streptococcus pneumoniae of the Essential Oil and δ-Cadinene Isolated from Schinus molle fruit. J. Essent. Oil Res. 2011, 23, 25–28. [Google Scholar] [CrossRef]
- Kirana, C.; McIntosh, G.H.; Record, I.R.; Jones, G.P. Antitumor activity of extract of Zingiber aromaticum and its bioactive sesquiterpenoid zerumbone. Nutr. Cancer 2003, 45, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Bustamam, A.; Ibrahim, S.; Al-Zubairi, A.S. Zerumbone: A natural compound with anti-cholinesterase activity. Am. J. Pharmacol. Toxicol. 2008, 3, 209–211. [Google Scholar] [CrossRef]
- Takahashi, Y.; Daitoh, M.; Suzuki, M.; Abe, T.; Masuda, M. Halogenated metabolites from the new Okinawan red alga Laurencia yonaguniensis. J. Nat. Prod. 2002, 65, 395–398. [Google Scholar] [CrossRef] [PubMed]
- Vairappan, C.S.; Ishii, T.; Lee, T.K.; Suzuki, M.; Zhaoqi, Z. Antibacterial activities of a new brominated diterpene from Borneon Laurencia spp. Mar. Drugs 2010, 8, 1743–1749. [Google Scholar] [CrossRef] [PubMed]
- Iliopoulou, D.; Mihopoulos, N.; Vagias, C.; Papazafiri, P.; Roussis, V. Novel cytotoxic brominated diterpenes from the red alga Laurencia obtusa. J. Org. Chem. 2003, 68, 7667–7674. [Google Scholar] [CrossRef] [PubMed]
- Ro, D.-K.; Arimura, G.-I.; Lau, S.Y.W.; Piers, E.; Bohlmann, J. Loblolly pine abietadienol/abietadienal oxidase PtAO (CYP720B1) is a multifunctional, multisubstrate cytochrome P450 monooxygenase. Proc. Natl. Acad. Sci. USA 2005, 102, 8060–8065. [Google Scholar] [CrossRef] [PubMed]
- Crouch, I.J.; Staden, J. Evidence for the presence of plant growth regulators in commercial seaweed products. Plant Growth Regul. 1993, 13, 21–29. [Google Scholar] [CrossRef]
- Stirk, W.A.; Tarkowská, D.; Turečová, V.; Strnad, M.; van Staden, J. Abscisic acid, gibberellins and brassinosteroids in Kelpak®, a commercial seaweed extract made from Ecklonia maxima. J. Appl. Phycol. 2014, 26, 561–567. [Google Scholar] [CrossRef]
- Oliveira, E.C.; Paula, E.J.; Plastino, E.M.; Petti, R. Metodologias para cultivo de algas em laboratório. In Macroalgas de Interes Económico: Cultivo, Manejo y Industrialización; Ferrario, M.E., Sar, E., Eds.; Universidad Nacional de La Plata: La Plata, Argentina, 1996; pp. 175–198. [Google Scholar]
- Schmieder, R.; Edwards, R. Quality control and preprocessing of metagenomic datasets. Bioinformatics 2011, 27, 863–864. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [PubMed]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef] [PubMed]
- Cane, D.E.; Ikeda, H. Exploration and mining of the bacterial terpenome. Acc. Chem. Res. 2012, 45, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Von Stosch, H.A. Wirkung von Arsenit auf Meeresalgen in Kultur. In Proceedings of the 4th International Seaweed Symposium, Biarritz, France, 18–24 September 1961; pp. 142–150.
- Curry, K. Initiation of terpenoid synthesis in osmophores of Stanhopea anfracta (Orchidaceae): A cytochemical study. Am. J. Bot. 1987, 74, 1332–1338. [Google Scholar] [CrossRef]
- Croteau, R.; Kutchan, T.M.; Lewis, N.G. Natural Products (Secondary Metabolites). In Biochemistry & Molecular Biology of Plants; Buchanan, B.B., Gruissem, W., Jones, R.L., Eds.; American Society of Plant Physiologists: Maryland, MD, USA, 2000; pp. 1251–1318. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Oliveira, L.S.; Tschoeke, D.A.; De Oliveira, A.S.; Hill, L.J.; Paradas, W.C.; Salgado, L.T.; Thompson, C.C.; Pereira, R.C.; Thompson, F.L. New Insights on the Terpenome of the Red Seaweed Laurencia dendroidea (Florideophyceae, Rhodophyta). Mar. Drugs 2015, 13, 879-902. https://doi.org/10.3390/md13020879
De Oliveira LS, Tschoeke DA, De Oliveira AS, Hill LJ, Paradas WC, Salgado LT, Thompson CC, Pereira RC, Thompson FL. New Insights on the Terpenome of the Red Seaweed Laurencia dendroidea (Florideophyceae, Rhodophyta). Marine Drugs. 2015; 13(2):879-902. https://doi.org/10.3390/md13020879
Chicago/Turabian StyleDe Oliveira, Louisi Souza, Diogo Antonio Tschoeke, Aline Santos De Oliveira, Lilian Jorge Hill, Wladimir Costa Paradas, Leonardo Tavares Salgado, Cristiane Carneiro Thompson, Renato Crespo Pereira, and Fabiano L. Thompson. 2015. "New Insights on the Terpenome of the Red Seaweed Laurencia dendroidea (Florideophyceae, Rhodophyta)" Marine Drugs 13, no. 2: 879-902. https://doi.org/10.3390/md13020879
APA StyleDe Oliveira, L. S., Tschoeke, D. A., De Oliveira, A. S., Hill, L. J., Paradas, W. C., Salgado, L. T., Thompson, C. C., Pereira, R. C., & Thompson, F. L. (2015). New Insights on the Terpenome of the Red Seaweed Laurencia dendroidea (Florideophyceae, Rhodophyta). Marine Drugs, 13(2), 879-902. https://doi.org/10.3390/md13020879