
Structural Analysis and Anti-Complement Activity of Polysaccharides from Kjellmaniella crsaaifolia

Abstract
:1. Introduction
2. Results and Discussion
2.1. Preparation of Polysaccharides

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Sugar | Fuc (%) | UA (%) | SO4 (%) | Protein (%) | Monosaccharides (Molar Ratio) | Mw (kDa) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Man | Rha | GlcA | Glc | Gal | Xyl | Fuc | |||||||
| KCW | 58.06 | 37.07 | – | 24.21 | 0.45 | 0.12 | 0.05 | 0.09 | 0.06 | 0.08 | 0.02 | 1 | 168.7 |
| DKCW | 53.56 | 30.75 | – | 22.52 | 0.74 | 0.12 | 0.10 | 0.12 | 0.06 | 0.06 | – | 1 | 5.2 |
| ds-DKCW | 82.42 | 38.79 | 3.72 | 8.93 | 1.22 | 0.23 | 0.11 | 0.19 | 0.16 | 0.21 | 0.20 | 1 | 3.5 |
| KCA | 53.40 | 40.43 | – | 35.49 | – | 0.05 | 0.02 | 0.06 | 0.02 | 0.03 | 0.02 | 1 | 153.7 |
| DKCA | 45.76 | 35.87 | – | 32.37 | – | 0.04 | 0.04 | 0.05 | 0.02 | 0.02 | – | 1 | 9.7 |
| ds-DKCA | 65.58 | 48.97 | 3.51 | 12.72 | 0.77 | 0.13 | 0.05 | 0.12 | 0.27 | 0.14 | 0.03 | 1 | 2.3 |
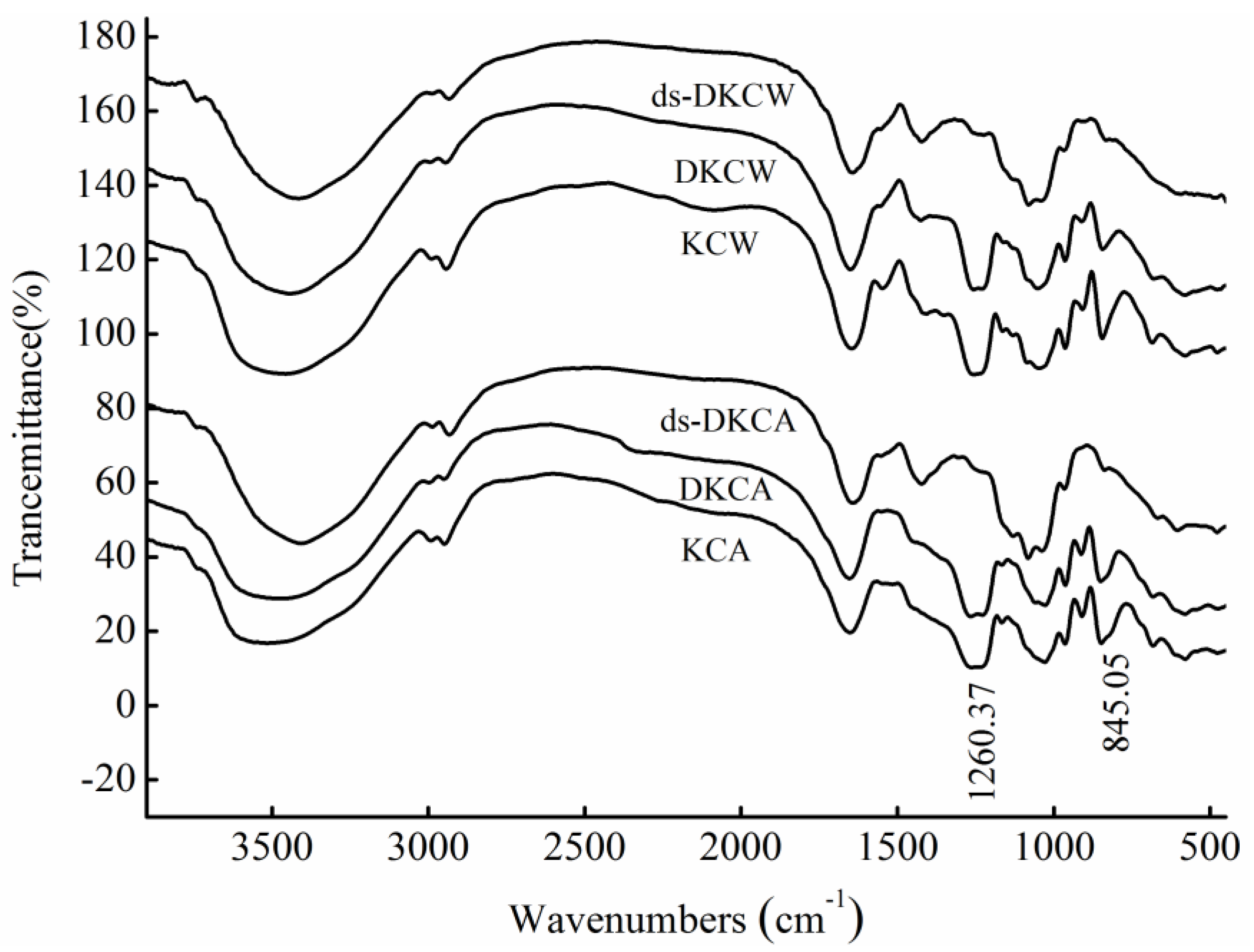
2.2. IR Analysis

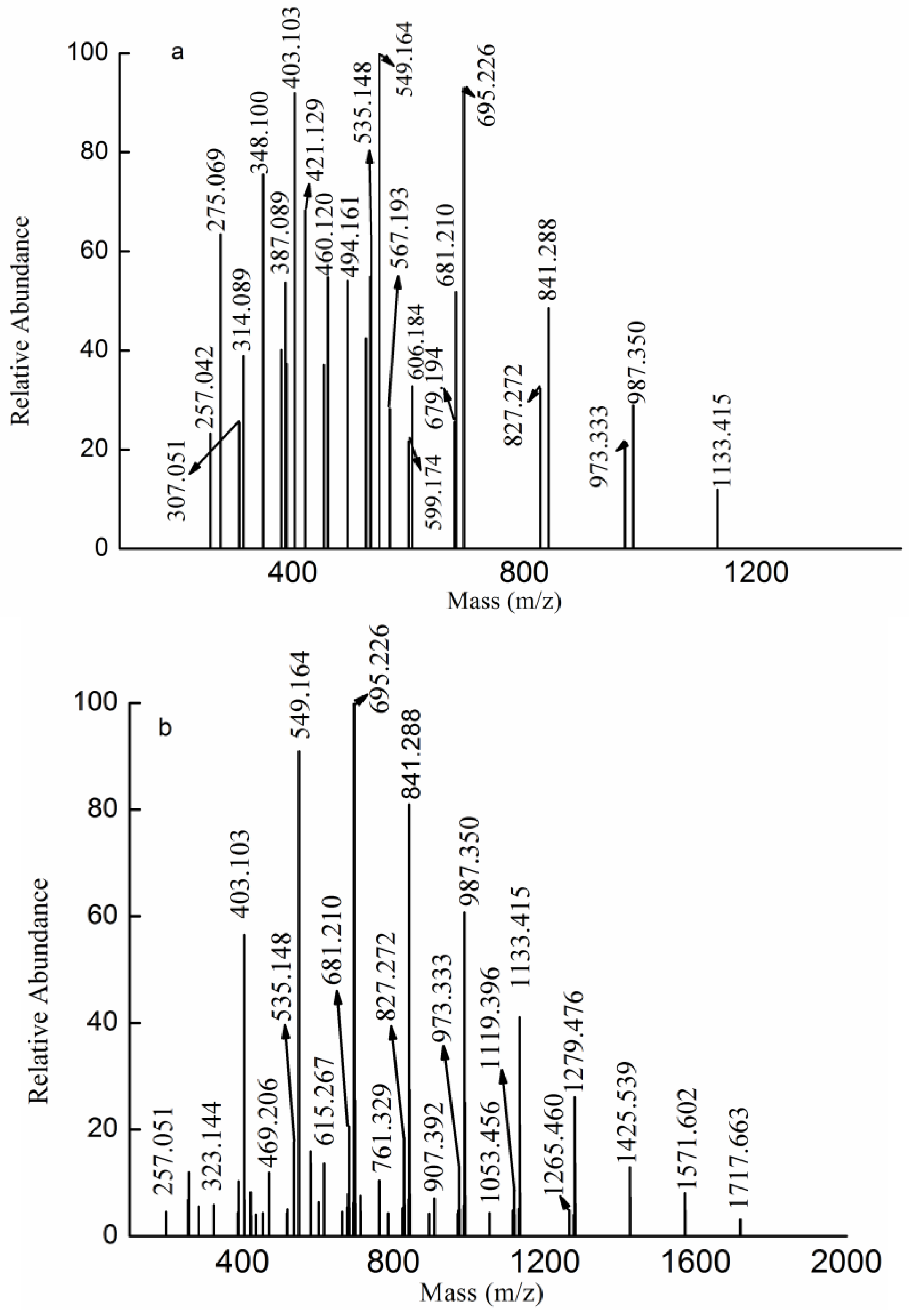
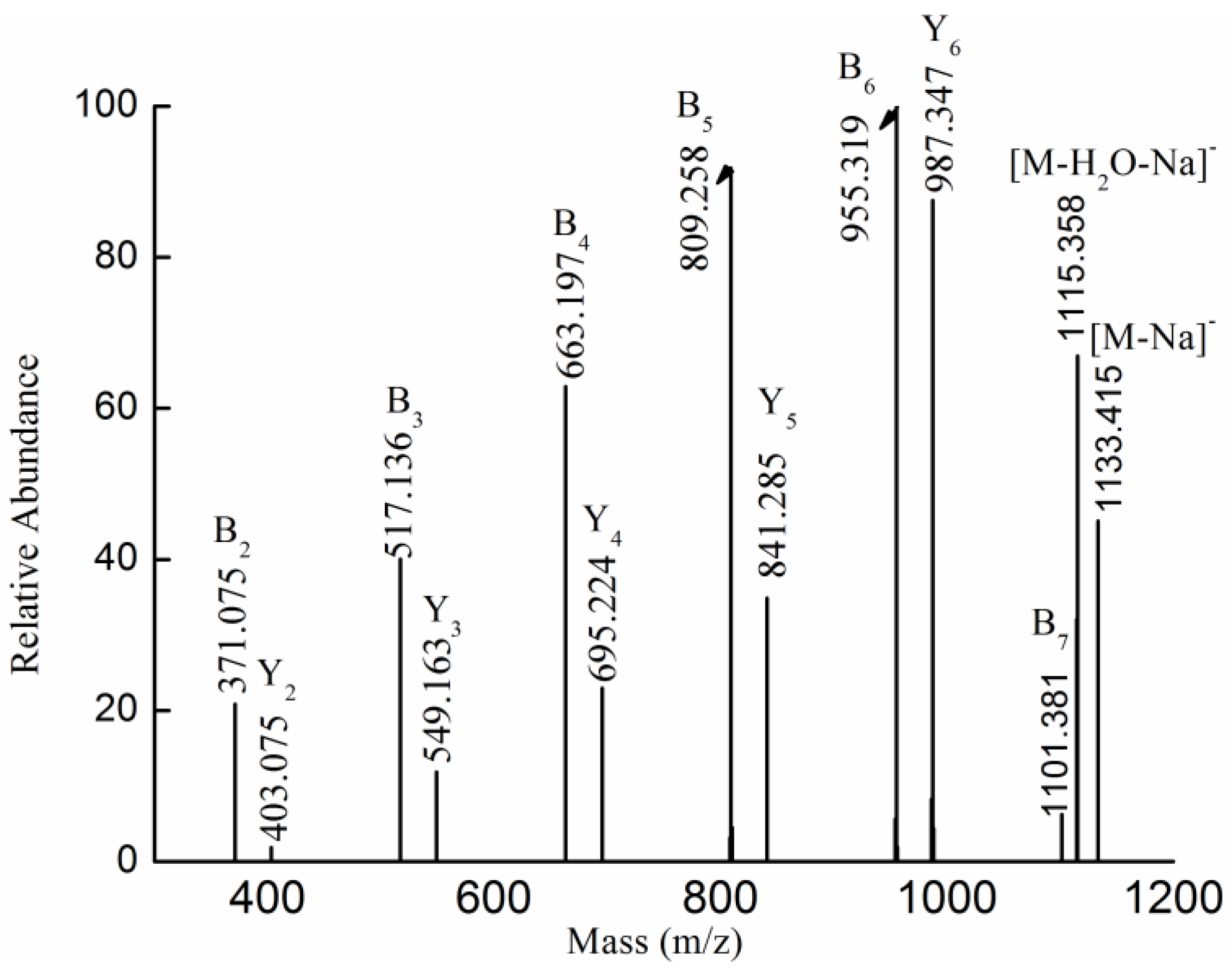
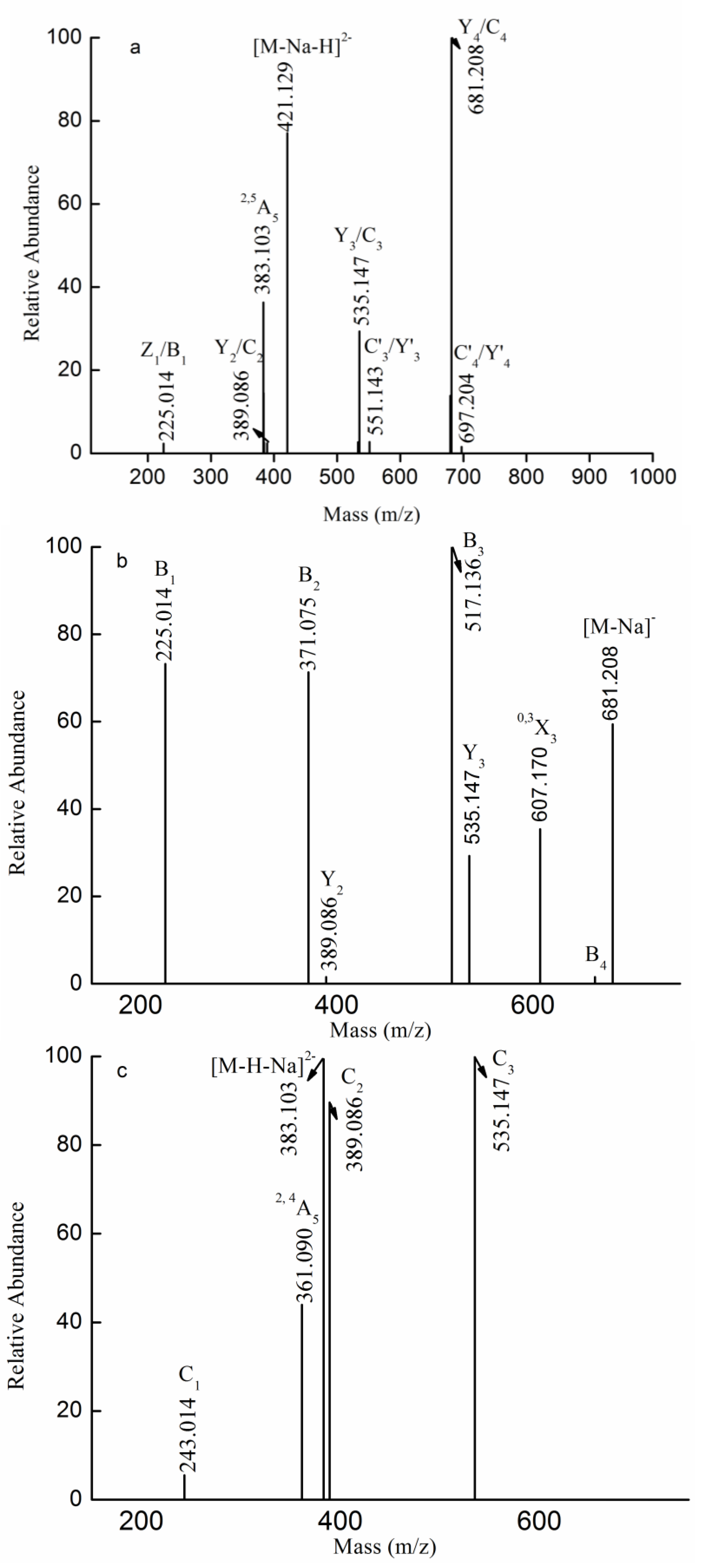
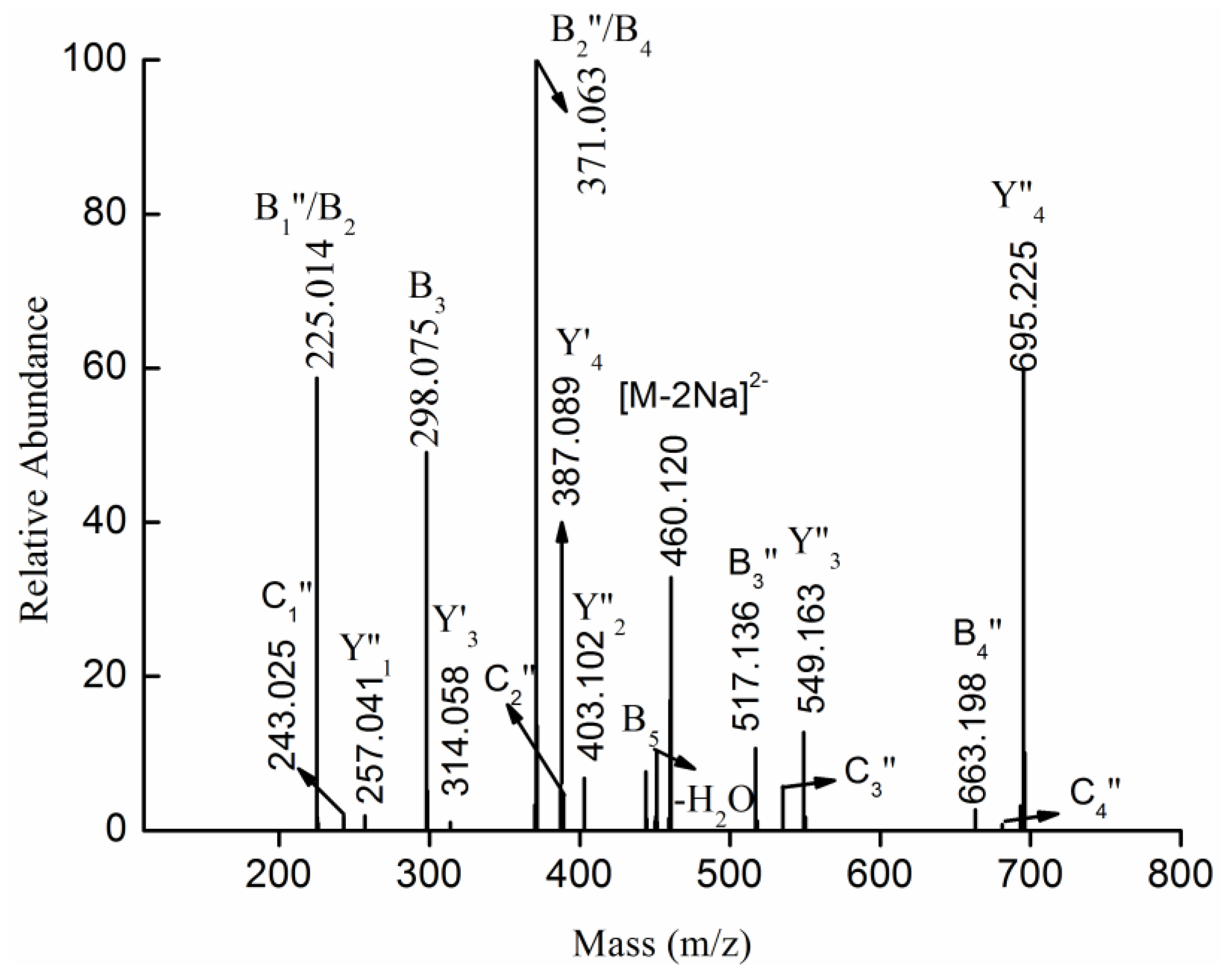
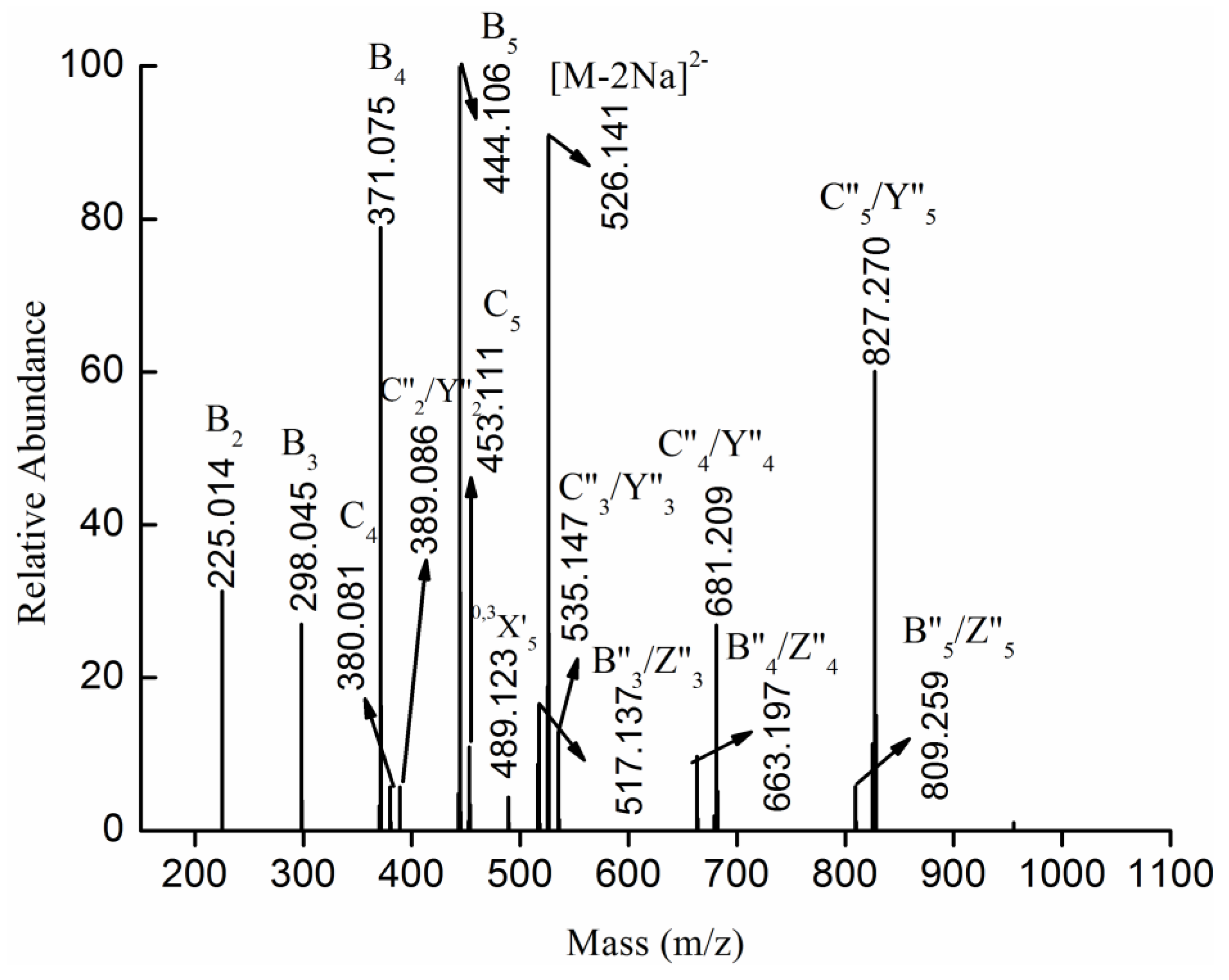
2.3. MS Analysis of Structure





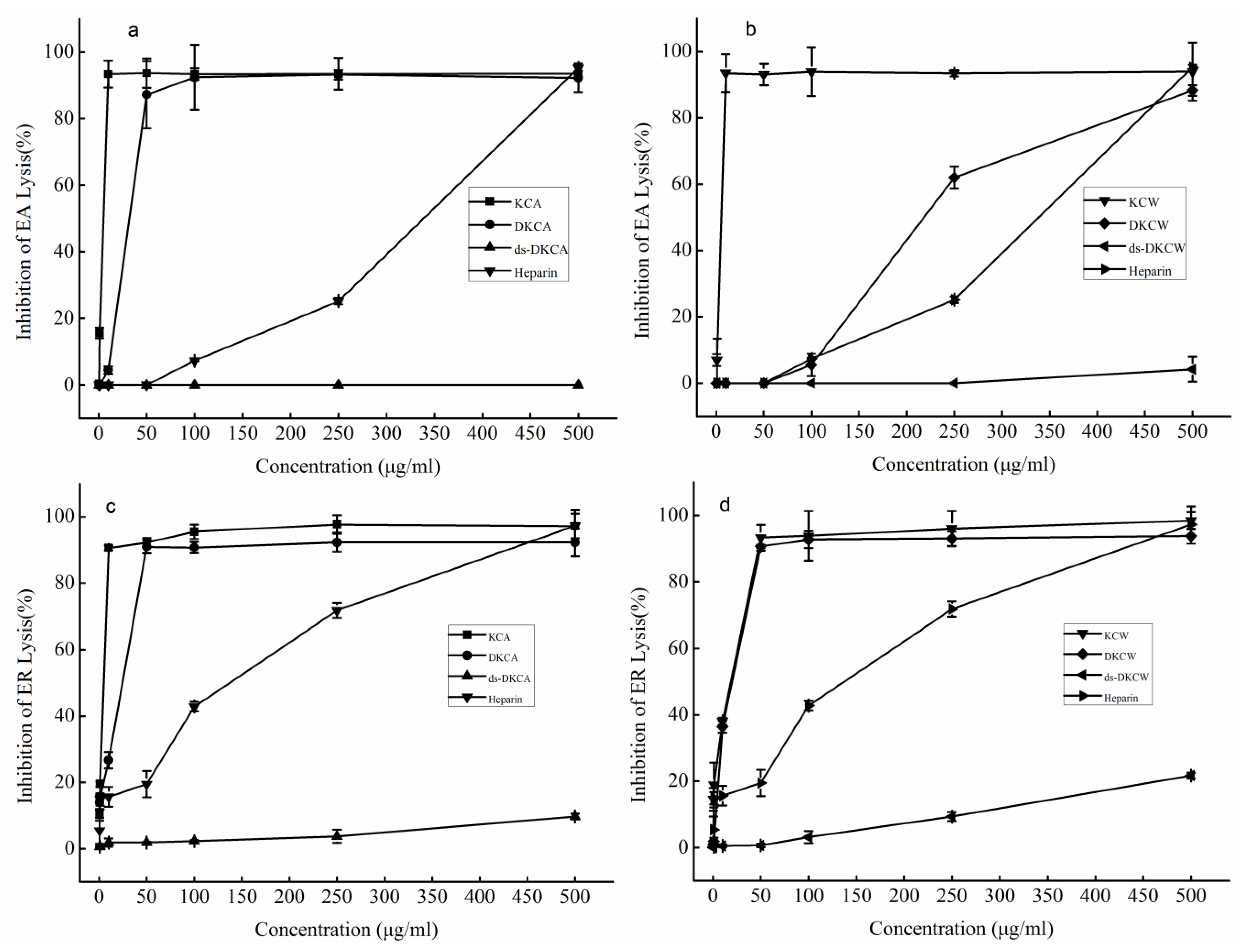
2.4. Anti-Complement Activity

3. Experimental Section
3.1. Preparation of Polysaccharides
3.2. Preparation of Low Molecular Weight Fucoidans and Their Desulfated Mixtures
3.3. Composition Methods
3.4. Spectroscopic Analysis
3.5. Anti-Complement Activity
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yan, H.; Kakuta, S.; Nishihara, M.; Sugi, M.; Adachi, Y.; Ohno, N.; Iwakura, Y.; Tsuji, N.M. Kjellmaniella crassifolia miyabe (Gagome) extract modulates intestinal and systemic immune responses. Biosci. Biotechnol. Biochem. 2011, 75, 2178–2183. [Google Scholar] [CrossRef] [PubMed]
- Katayama, S.; Nishio, T.; Kishimura, H.; Saeki, H. Immunomodulatory properties of highly viscous polysaccharide extract from the Gagome alga (Kjellmaniella crassifolia). Plant Foods Hum. Nutr. 2012, 67, 76–81. [Google Scholar] [CrossRef]
- Sakai, T.; Kimura, H.; Kojima, K.; Shimanaka, K.; Ikai, K.; Kato, I. Marine bacterial sulfated fucoglucuronomannan (SFGM) lyase digests brown algal SFGM into trisaccharides. Mar. Biotechnol. (N.Y.) 2003, 5, 70–78. [Google Scholar] [CrossRef]
- Wu, J.; Yu, G.; Li, M.; Zhao, X.; Wang, C.; Liu, T.; Gu, C. Extraction, isolation and structural characterization of polysaccharides from Kjellmanjella crassifolia. J. Ocean Univ. China 2011, 41, 127–130. [Google Scholar]
- Wang, P.; Zhao, X.; Lv, Y.; Liu, Y.; Lang, Y.; Wu, J.; Liu, X.; Li, M.; Yu, G. Analysis of structural heterogeneity of fucoidan from Hizikia fusiforme by ES-CID-MS/MS. Carbohydr. Polym. 2012, 90, 602–607. [Google Scholar] [CrossRef] [PubMed]
- Anastyuk, S.D.; Shevchenko, N.M.; Nazarenko, E.L.; Imbs, T.I.; Gorbach, V.I.; Dmitrenok, P.S.; Zvyagintseva, T.N. Structural analysis of a highly sulfated fucan from the brown alga Laminaria cichorioides by tandem MALDI and ESI mass spectrometry. Carbohydr. Res. 2010, 345, 2206–2212. [Google Scholar] [CrossRef] [PubMed]
- Daniel, R.; Chevolot, L.; Carrascal, M.; Tissot, B.; Mourão, P.A.S.; Abian, J. Electrospray ionization mass spectrometry of oligosaccharides derived from fucoidan of Ascophyllum nodosum. Carbohydr. Res. 2007, 342, 826–834. [Google Scholar] [CrossRef] [PubMed]
- Makrides, S.C. Therapeutic inhibition of the complement system. Pharmacol. Rev. 1998, 50, 59–88. [Google Scholar] [PubMed]
- Blondin, C.; Chaubet, F.; Nardella, A.; Sinquin, C.; Jozefonvicz, J. Relationships between chemical characteristics and anticomplementary activity of fucans. Biomaterials 1996, 17, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Clement, M.J.; Tissot, B.; Chevolot, L.; Adjadj, E.; Du, Y.; Curmi, P.A.; Daniel, R. NMR characterization and molecular modeling of fucoidan showing the importance of oligosaccharide branching in its anticomplementary activity. Glycobiology 2010, 20, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Tissot, B.; Montdargent, B.; Chevolot, L.; Varenne, A.; Descroix, S.; Gareil, P.; Daniel, R. Interaction of fucoidan with the proteins of the complement classical pathway. Biochim. Biophys. Acta 2003, 1651, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Samuelsena, A.B.; Lunda, I.; Djahromia, J.M.; Paulsena, B.S.; Wolda, J.K.; Knutsen, H.S. Structural features and anti-complementary activity of some heteroxylan polysaccharide fractions from the seeds of Plantago major L. Carbohydr. Polym. 1999, 38, 133–143. [Google Scholar] [CrossRef]
- Mauzac, M.; Maillet, F.; Jozefonvicz, J.; Kazatchkine, M.D. Anticomplementary activity of dextran derivatives. Biomaterials 1985, 6, 61–63. [Google Scholar] [CrossRef] [PubMed]
- Casu, B. Structure and biological activity of heparin and other glycosaminoglycans. Pharmacol. Res. Commun. 1979, 11, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Foley, S.A.; Mulloy, B.; Tuohy, M.G. An unfractionated fucoidan from Ascophyllum nodosum: Extraction, characterization, and apoptotic effects in vitro. J. Nat. Prod. 2011, 74, 1851–1861. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Xue, C.; Yin, L.A.; Tang, Q.; Yu, G.; Chai, W. Comparison of structures and anticoagulant activities of fucosylated chondroitin sulfates from different sea cucumbers. Carbohydr. Polym. 2011, 83, 688–696. [Google Scholar] [CrossRef]
- Duarte, M.E. R.; Cardoso, M.A.; Noseda, M.D.; Cerezo, A.S. Structural studies on fucoidans from the brown seaweed Sargassum stenophyllum. Carbohydr. Res. 2001, 333, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Anastyuk, S.D.; Imbs, T.I.; Shevchenko, N.M.; Dmitrenok, P.S.; Zvyagintseva, T.N. ESIMS analysis of fucoidan preparations from Costaria costata, extracted from alga at different life-stages. Carbohydr. Polym. 2012, 90, 993–1002. [Google Scholar] [CrossRef] [PubMed]
- Anastyuk, S.D.; Shevchenko, N.M.; Nazarenko, E.L.; Dmitrenok, P.S.; Zvyagintseva, T.N. Structural analysis of a fucoidan from the brown alga Fucus evanescens by MALDI-TOF and tandem ESI mass spectrometry. Carbohydr. Res. 2009, 344, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Tissot, B.; Salpin, J.Y.; Martinez, M.; Gaigeot, M.P.; Daniel, R. Differentiation of the fucoidan sulfated l-fucose isomers constituents by CE-ESIMS and molecular modeling. Carbohydr. Res. 2006, 341, 598–609. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Wang, J.; Ren, S.; Song, N.; Zhang, Q. Structural analysis of a heteropolysaccharide from Saccharina japonica by electrospray mass spectrometry in tandem with collision-induced dissociation tandem mass spectrometry (ESI-CID-MS/MS). Mar. Drugs 2012, 10, 2138–2152. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Zhang, W.; Wang, J.; Ren, S.; Song, N.; Zhang, Q. Structural analysis of heteropolysaccharide from Saccharina japonica and its derived oligosaccharides. Int. J. Biol. Macromol. 2013, 62C, 697–704. [Google Scholar] [CrossRef]
- Jin, W.; Zhang, W.; Wang, J.; Ren, S.; Song, N.; Duan, D.; Zhang, Q. Characterization of laminaran and a highly sulfated polysaccharide from Sargassum fusiforme. Carbohydr. Res. 2014, 385, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Guo, Z.; Wang, J.; Zhang, W.; Zhang, Q. Structural analysis of sulfated fucan from Saccharina japonica by electrospray ionization tandem mass spectrometry. Carbohydr. Res. 2013, 369, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Saad, O.M.; Leary, J.A. Delineating mechanisms of dissociation for isomeric heparin disaccharides using isotope labeling and ion trap tandem mass spectrometry. Am. Soc. Mass Spectrom. 2004, 15, 1274–1286. [Google Scholar] [CrossRef]
- Crepon, B.; Maillet, F.; Kazatchkine, M.D.; Jozefonvicz, J. Molecular weight dependency of the acquired anticomplementary and anticoagulant activities of specifically substituted dextrans. Biomaterials 1987, 8, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Cofrancsco, E.; Radaelli, F.; Pogliania, E. Correlation of sulfate content and degree of carboxylation of heparin and related glycosaminoglycans with anticomplement activity. Relationships to the anticoagulant and platelet-aggregating activities. Thromb. Res. 1979, 14, 179–187. [Google Scholar]
- Maillet, F.; Maurice Petitou; Jean Choay; Kazatchkine, M.D. Structure-function relationships in the inhibitory effect of heparin on complement activation: Independency of the anti-coagulant and anti-complementary sites on the heparin molecule. Mol. Immunol. 1988, 25, 917–923. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Zhang, W.; Wang, J.; Zhang, Q. The neuroprotective activities and antioxidant activities of the polysaccharides from Saccharina japonica. Int. J. Biol. Macromol. 2013, 58, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, K.; Inoue, Y.; Tokuyasu, T. An improved method for the preparation of chondroitin by solvolytic desulfation of chondroitin sulfates. J. Biochem. 1979, 86, 1323–1329. [Google Scholar] [PubMed]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Gibbons, M.N. The determination of methylpentoses. Analyst 1955, 80, 267–276. [Google Scholar] [CrossRef]
- Kawai, Y.; Seno, N.; Anno, K. A modified method for chondrosulfatase assay. Anal. Biochem. 1969, 32, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Bitter, T.; Muir, H.M. A modified uronic acid carbazole reaction. Anal. Biochem. 1962, 4, 330–334. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, Q.; Wang, J.; Shi, X.; Zhang, Z. Analysis of the monosaccharide composition of fucoidan by precolumn derivation HPLC. Chin. J. Oceanol. Limnol. 2009, 27, 1–5. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, J.; Jin, W.; Zhang, Q. The antioxidant activities and neuroprotective effect of polysaccharides from the starfish Asterias rollestoni. Carbohydr. Polym. 2013, 95, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Zhang, Y.; Zhang, J.; Chen, D. Isolation and characterization of an anti-complementary polysaccharide D3-S1 from the roots of Bupleurum smithii. Int. Immunopharmacol. 2007, 7, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Klerx, J.P.A.M.; Beukelman, C.J.; Dijk, H.V.; Willers, J.M.N. Microassay for colorimetric estimation of complement activity in guinea pig, human and mouse serum. J. Immunol. Methods 1983, 63, 215–220. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Jin, W.; Sun, D.; Zhao, L.; Wang, J.; Duan, D.; Zhang, Q. Structural Analysis and Anti-Complement Activity of Polysaccharides from Kjellmaniella crsaaifolia. Mar. Drugs 2015, 13, 1360-1374. https://doi.org/10.3390/md13031360
Zhang W, Jin W, Sun D, Zhao L, Wang J, Duan D, Zhang Q. Structural Analysis and Anti-Complement Activity of Polysaccharides from Kjellmaniella crsaaifolia. Marine Drugs. 2015; 13(3):1360-1374. https://doi.org/10.3390/md13031360
Chicago/Turabian StyleZhang, Wenjing, Weihua Jin, Delin Sun, Luyu Zhao, Jing Wang, Delin Duan, and Quanbin Zhang. 2015. "Structural Analysis and Anti-Complement Activity of Polysaccharides from Kjellmaniella crsaaifolia" Marine Drugs 13, no. 3: 1360-1374. https://doi.org/10.3390/md13031360