
Functional and Structural Characterization of FAU Gene/Protein from Marine Sponge Suberites domuncula

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
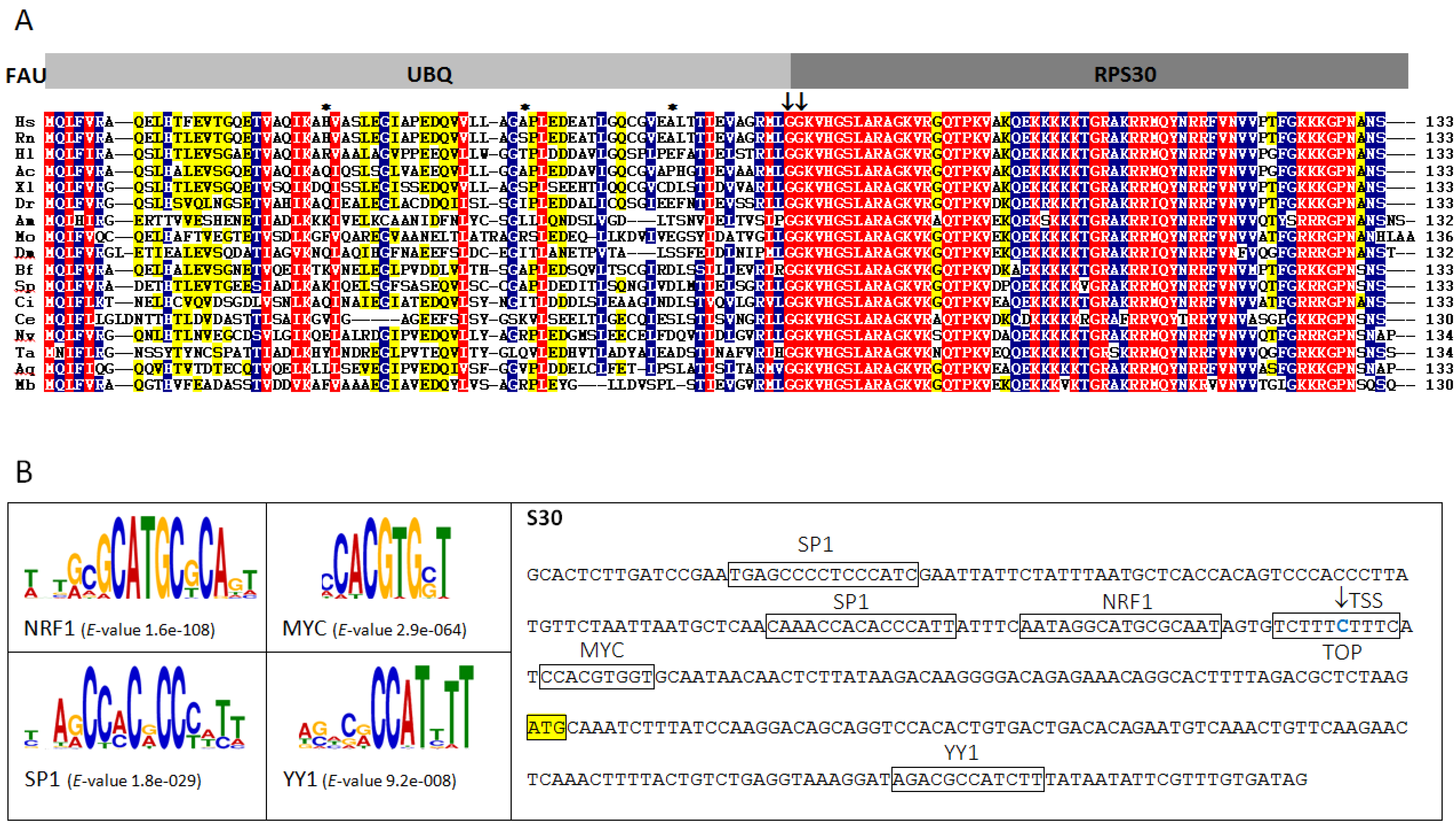
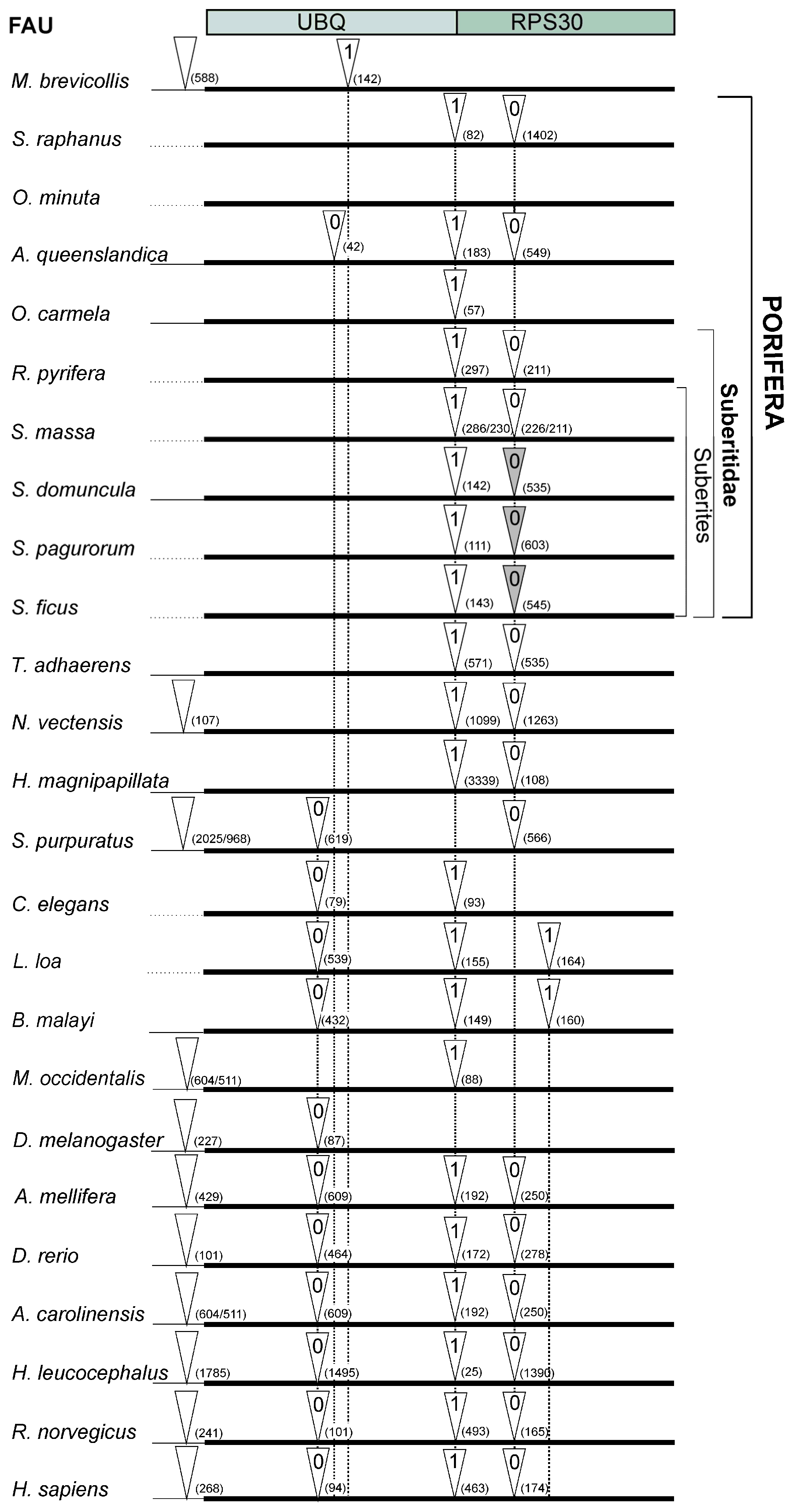
2.1. Structure and Evolution of Metazoan FAU Gene

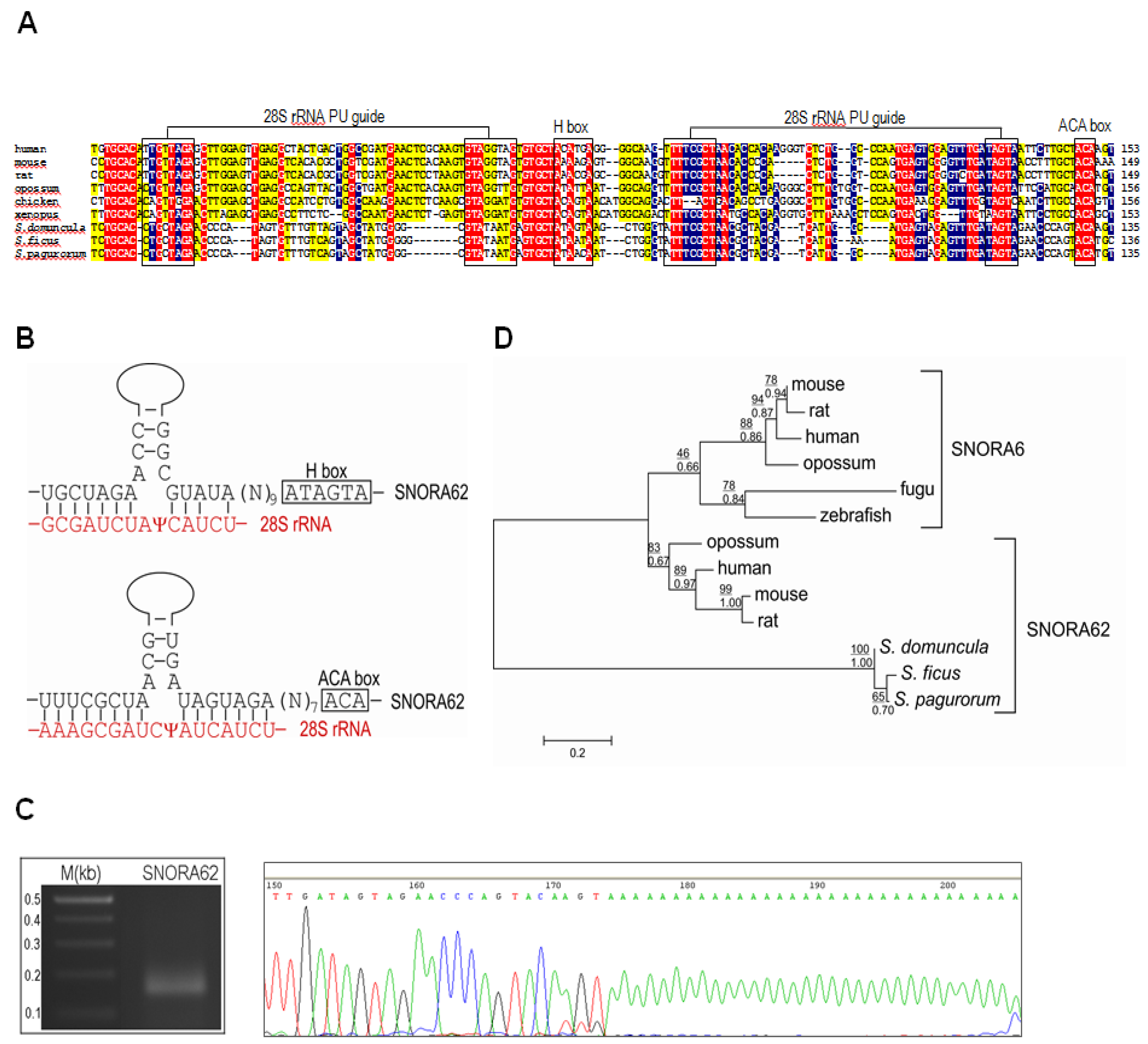
2.2. Identification of snoRNAs in FAU Introns of Sponges


2.3. Subcellular Localization of Sponge FAU


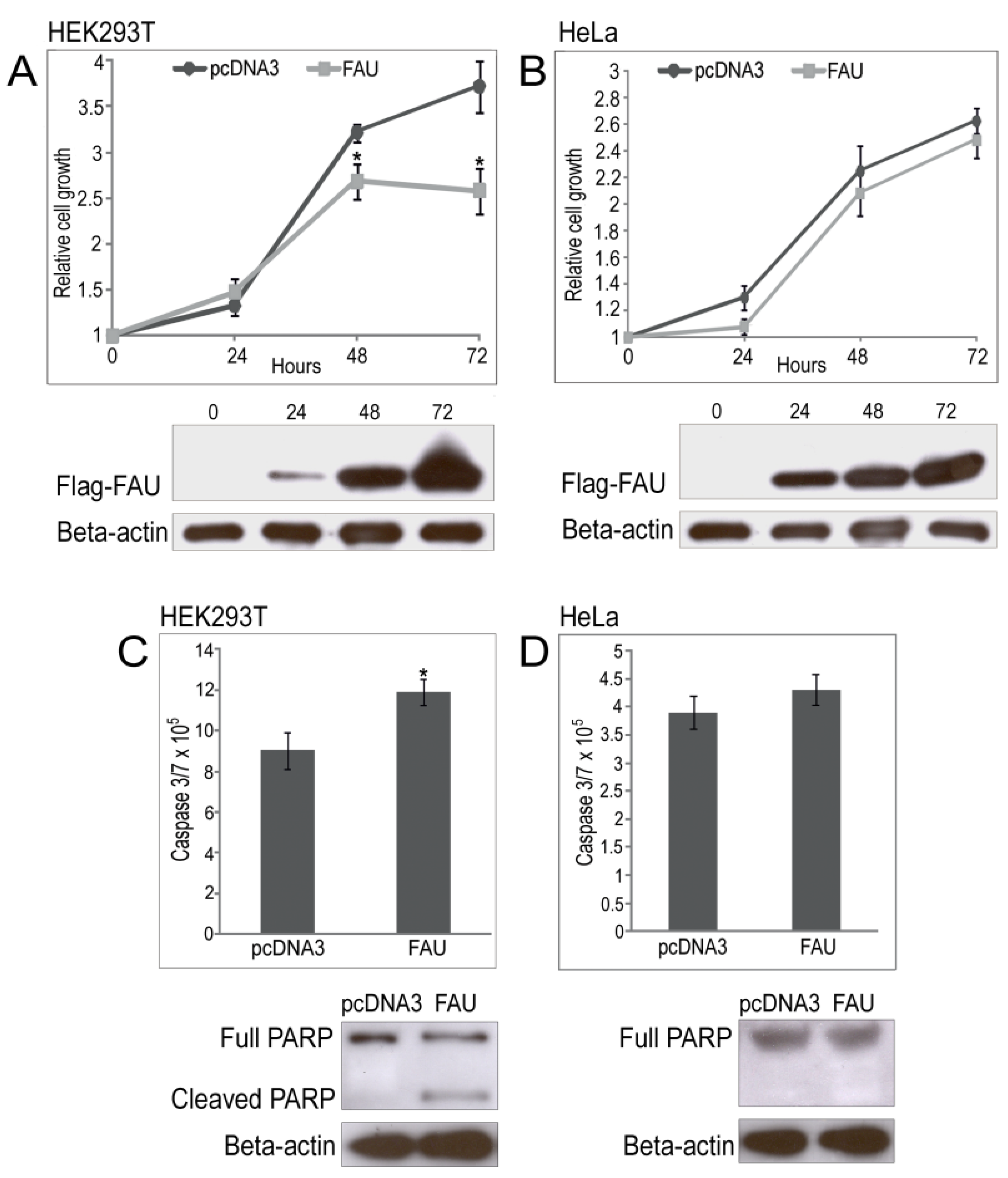
2.4. Overexpression of Sponge FAU in Human Cell Lines
3. Experimental Section
3.1. Materials
3.2. Sequence Analyses
3.3. Isolation of Genomic DNA and Sequencing of FAU Gene
3.4. Isolation and Characterization of Small RNAs
3.5. Cell Culture
3.6. Plasmid Constructions
3.7. Transient Transfections and Laser Scanning Confocal Microscopy
3.8. Immunoblotting
3.9. Cell Viability Assay
3.10. Apoptosis Assay
3.11. Statistical Analyses
Supplementary Files
Supplementary File 1Acknowledgment
Author Contributions
Conflicts of Interest
References
- Davies, P.C.; Lineweaver, C.H. Cancer tumors as Metazoa 1.0: Tapping genes of ancient ancestors. Phys. Biol. 2011, 8, 015001. [Google Scholar] [PubMed]
- Srivastava, M.; Simakov, O.; Chapman, J.; Fahey, B.; Gauthier, M.E.; Mitros, T.; Richards, G.S.; Conaco, C.; Dacre, M.; Hellsten, U.; et al. The Amphimedon queenslandica genome and the evolution of animal complexity. Nature 2010, 466, 720–726. [Google Scholar] [PubMed]
- Perina, D.; Bosnar, M.H.; Bago, R.; Mikoc, A.; Harcet, M.; Dezeljin, M.; Cetkovic, H. Sponge non-metastatic Group I NME gene/protein-structure and function is conserved from sponges to humans. BMC Evol. Biol. 2011, 11, 87. [Google Scholar] [CrossRef] [PubMed]
- Perina, D.; Bosnar, M.H.; Mikoc, A.; Muller, W.E.; Cetkovic, H. Characterization of NME6-like gene/protein from marine sponge Suberites domuncula. Naunyn Schmiedeberg Arch. Pharmacol. 2011, 384, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Domazet-Loso, T.; Tautz, D. Phylostratigraphic tracking of cancer genes suggests a link to the emergence of multicellularity in metazoa. BMC Biol. 2010, 8, 66. [Google Scholar] [CrossRef] [PubMed]
- Domazet-Loso, T.; Klimovich, A.; Anokhin, B.; Anton-Erxleben, F.; Hamm, M.J.; Lange, C.; Bosch, T.C. Naturally occurring tumours in the basal metazoan Hydra. Nat. Commun. 2014, 5, 4222. [Google Scholar] [CrossRef] [PubMed]
- Perry, R.P. Balanced production of ribosomal proteins. Gene 2007, 401, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Kressler, D.; Hurt, E.; Bassler, J. Driving ribosome assembly. Biochim. Biophys. Acta 2010, 1803, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Gardner, P.P.; Bateman, A.; Poole, A.M. SnoPatrol: How many snoRNA genes are there? J. Biol. 2010, 9, 4. [Google Scholar] [CrossRef] [PubMed]
- Scott, M.S.; Ono, M. From snoRNA to miRNA: Dual function regulatory non-coding RNAs. Biochimie 2011, 93, 1987–1992. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.J. Mammalian small nucleolar RNAs are mobile genetic elements. PLoS Genet. 2006, 2, e205. [Google Scholar] [CrossRef] [PubMed]
- Lecompte, O.; Ripp, R.; Thierry, J.C.; Moras, D.; Poch, O. Comparative analysis of ribosomal proteins in complete genomes: An example of reductive evolution at the domain scale. Nucleic Acids Res. 2002, 30, 5382–5390. [Google Scholar] [CrossRef] [PubMed]
- Warner, J.R.; McIntosh, K.B. How common are extraribosomal functions of ribosomal proteins? Mol. Cell 2009, 34, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Kas, K.; Michiels, L.; Merregaert, J. Genomic structure and expression of the human FAU gene: Encoding the ribosomal protein S30 fused to a ubiquitin-like protein. Biochem. Biophys. Res. Commun. 1992, 187, 927–933. [Google Scholar] [CrossRef]
- Michiels, L.; van der Rauwelaert, E.; van Hasselt, F.; Kas, K.; Merregaert, J. FAU cDNA encodes a ubiquitin-like-S30 fusion protein and is expressed as an antisense sequence in the Finkel-Biskis-Reilly murine sarcoma virus. Oncogene 1993, 8, 2537–2546. [Google Scholar] [PubMed]
- Pickard, M.R.; Green, A.R.; Ellis, I.O.; Caldas, C.; Hedge, V.L.; Mourtada-Maarabouni, M.; Williams, G.T. Dysregulated expression of FAU and MELK is associated with poor prognosis in breast cancer. Breast Cancer Res. 2009, 11, R60. [Google Scholar] [CrossRef]
- Pickard, M.R.; Edwards, S.E.; Cooper, C.S.; Williams, G.T. Apoptosis regulators FAU and Bcl-G are down-regulated in prostate cancer. Prostate 2010, 70, 1513–1523. [Google Scholar] [CrossRef] [PubMed]
- Moss, E.L.; Mourtada-Maarabouni, M.; Pickard, M.R.; Redman, C.W.; Williams, G.T. FAU regulates carboplatin resistance in ovarian cancer. Genes Chromosomes Cancer 2010, 49, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Pickard, M.R.; Mourtada-Maarabouni, M.; Williams, G.T. Candidate tumour suppressor FAU regulates apoptosis in human cells: An essential role for Bcl-G. Biochim. Biophys. Acta 2011, 1812, 1146–1153. [Google Scholar] [CrossRef] [PubMed]
- Alba, M.M.; Castresana, J. Inverse relationship between evolutionary rate and age of mammalian genes. Mol. Biol. Evol. 2005, 22, 598–606. [Google Scholar] [CrossRef] [PubMed]
- Roepcke, S.; Zhi, D.; Vingron, M.; Arndt, P.F. Identification of highly specific localized sequence motifs in human ribosomal protein gene promoters. Gene 2006, 365, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Perina, D.; Korolija, M.; Roller, M.; Harcet, M.; Jelicic, B.; Mikoc, A.; Cetkovic, H. Over-represented localized sequence motifs in ribosomal protein gene promoters of basal metazoans. Genomics 2011, 98, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Meyuhas, O. Synthesis of the translational apparatus is regulated at the translational level. Eur. J. Biochem. 2000, 267, 6321–6330. [Google Scholar] [CrossRef] [PubMed]
- Shibui-Nihei, A.; Ohmori, Y.; Yoshida, K.; Imai, J.; Oosuga, I.; Iidaka, M.; Suzuki, Y.; Mizushima-Sugano, J.; Yoshitomo-Nakagawa, K.; Sugano, S. The 5′ terminal oligopyrimidine tract of human elongation factor 1A-1 gene functions as a transcriptional initiator and produces a variable number of US at the transcriptional level. Gene 2003, 311, 137–145. [Google Scholar] [CrossRef]
- Perry, R.P. The architecture of mammalian ribosomal protein promoters. BMC Evol. Biol. 2005, 5, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Riggelen, J.; Yetil, A.; Felsher, D.W. MYC as a regulator of ribosome biogenesis and protein synthesis. Nat. Rev. Cancer 2010, 10, 301–309. [Google Scholar]
- Oskarsson, T.; Trumpp, A. The Myc trilogy: Lord of RNA polymerases. Nat. Cell Biol. 2005, 7, 215–217. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, R.; Suzuki, Y.; Takeuchi, N.; Wakaguri, H.; Ueda, T.; Sugano, S.; Nakai, K. Comprehensive detection of human terminal oligo-pyrimidine (TOP) genes and analysis of their characteristics. Nucleic Acids Res. 2008, 36, 3707–3715. [Google Scholar] [CrossRef] [PubMed]
- Perina, D.; Korolija, M.; Mikoc, A.; Roller, M.; Plese, B.; Imesek, M.; Morrow, C.; Batel, R.; Cetkovic, H. Structural and functional characterization of ribosomal protein gene introns in sponges. PLoS ONE 2012, 7, e42523. [Google Scholar] [CrossRef] [PubMed]
- Yoshihama, M.; Uechi, T.; Asakawa, S.; Kawasaki, K.; Kato, S.; Higa, S.; Maeda, N.; Minoshima, S.; Tanaka, T.; Shimizu, N.; et al. The human ribosomal protein genes: Sequencing and comparative analysis of 73 genes. Genome Res. 2002, 12, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Sela, N.; Kim, E.; Ast, G. The role of transposable elements in the evolution of non-mammalian vertebrates and invertebrates. Genome Biol. 2010, 11, R59. [Google Scholar] [CrossRef] [PubMed]
- Dieci, G.; Preti, M.; Montanini, B. Eukaryotic snoRNAs: A paradigm for gene expression flexibility. Genomics 2009, 94, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Ganot, P.; Caizergues-Ferrer, M.; Kiss, T. The family of box ACA small nucleolar RNAs is defined by an evolutionarily conserved secondary structure and ubiquitous sequence elements essential for RNA accumulation. Genes Dev. 1997, 11, 941–956. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Li, S. Genome-wide analyses of retrogenes derived from the human box H/ACA snoRNAs. Nucleic Acids Res. 2007, 35, 559–571. [Google Scholar] [PubMed]
- Bompfunewerer, A.F.; Flamm, C.; Fried, C.; Fritzsch, G.; Hofacker, I.L.; Lehmann, J.; Missal, K.; Mosig, A.; Muller, B.; Prohaska, S.J.; et al. Evolutionary patterns of non-coding RNAs. Theory Biosci. 2005, 123, 301–369. [Google Scholar] [CrossRef] [PubMed]
- Brogna, S.; Sato, T.A.; Rosbash, M. Ribosome components are associated with sites of transcription. Mol. Cell 2002, 10, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Giam, M.; Okamoto, T.; Mintern, J.D.; Strasser, A.; Bouillet, P. Bcl-2 family member Bcl-G is not a proapoptotic protein. Cell Death Dis. 2012, 3, e404. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.T.; Farzaneh, F. Are snoRNAs and snoRNA host genes new players in cancer? Nat. Rev. Cancer 2012, 12, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Mannoor, K.; Liao, J.; Jiang, F. Small nucleolar RNAs in cancer. Biochim. Biophys. Acta 2012, 1826, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Moodley, K.; Weiss, S.F. Down-regulation of the non-integrin laminin receptor reduces cellular viability by inducing apoptosis in lung and cervical cancer cells. PLoS ONE 2013, 8, e57409. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.P.; Chung, H.W.; Liu, K.C.; Wu, R.S.; Yang, J.S.; Tang, N.Y.; Lo, C.; Hsia, T.C.; Yu, C.C.; Chueh, F.S.; et al. Diallyl sulfide induces cell cycle arrest and apoptosis in HeLa human cervical cancer cells through the p53, caspase- and mitochondria-dependent pathways. Int. J. Oncol. 2011, 38, 1605–1613. [Google Scholar] [PubMed]
- Mei, Y.P.; Liao, J.P.; Shen, J.; Yu, L.; Liu, B.L.; Liu, L.; Li, R.Y.; Ji, L.; Dorsey, S.G.; Jiang, Z.R.; et al. Small nucleolar RNA 42 acts as an oncogene in lung tumorigenesis. Oncogene 2012, 31, 2794–2804. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.Y.; Rodriguez, C.; Guo, P.; Sun, X.; Talbot, J.T.; Zhou, W.; Petros, J.; Li, Q.; Vessella, R.L.; Kibel, A.S.; et al. SnoRNA U50 is a candidate tumor-suppressor gene at 6q14.3 with a mutation associated with clinically significant prostate cancer. Hum. Mol. Genet. 2008, 17, 1031–1042. [Google Scholar] [CrossRef] [PubMed]
- Grimson, A.; Srivastava, M.; Fahey, B.; Woodcroft, B.J.; Chiang, H.R.; King, N.; Degnan, B.M.; Rokhsar, D.S.; Bartel, D.P. Early origins and evolution of microRNAs and Piwi-interacting RNAs in animals. Nature 2008, 455, 1193–1197. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Elkan, C. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc. Int. Conf. Intell. Syst. Mol. Biol. 1994, 2, 28–36. [Google Scholar] [PubMed]
- Gupta, S.; Stamatoyannopoulos, J.A.; Bailey, T.L.; Noble, W.S. Quantifying similarity between motifs. Genome Biol. 2007, 8, R24. [Google Scholar] [CrossRef] [PubMed]
- Matys, V.; Fricke, E.; Geffers, R.; Gossling, E.; Haubrock, M.; Hehl, R.; Hornischer, K.; Karas, D.; Kel, A.E.; Kel-Margoulis, O.V.; et al. TRANSFAC: Transcriptional regulation, from patterns to profiles. Nucleic Acids Res. 2003, 31, 374–378. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.H.; Zhang, X.C.; Huang, Z.P.; Zhou, H.; Huang, M.B.; Zhang, S.; Chen, Y.Q.; Qu, L.H. snoSeeker: An advanced computational package for screening of guide and orphan snoRNA genes in the human genome. Nucleic Acids Res. 2006, 34, 5112–5123. [Google Scholar] [CrossRef]
- Gardner, P.P.; Daub, J.; Tate, J.G.; Nawrocki, E.P.; Kolbe, D.L.; Lindgreen, S.; Wilkinson, A.C.; Finn, R.D.; Griffiths-Jones, S.; Eddy, S.R.; et al. Rfam: Updates to the RNA families database. Nucleic Acids Res. 2009, 37, D136–D140. [Google Scholar] [CrossRef] [PubMed]
- Lestrade, L.; Weber, M.J. snoRNA-LBME-db, a comprehensive database of human H/ACA and C/D box snoRNAs. Nucleic Acids Res. 2006, 34, D158–D162. [Google Scholar] [CrossRef] [PubMed]
- Hofacker, I.L.; Fontana, W.; Stadler, P.F.; Bonhoeffer, L.S.; Tacker, M.; Schuster, P. Fast folding and comparison of RNA secondary structures. Monatsh. Chem. 1994, 125, 167–188. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Posada, D.; Crandall, K.A. MODELTEST: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, M.; Kishino, H.; Yano, T. Dating of the human-Ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Graham, F.L.; van der Eb, A.J. A new technique for the assay of infectivity of human adenovirus 5 DNA. Virology 1973, 52, 456–467. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perina, D.; Korolija, M.; Hadžija, M.P.; Grbeša, I.; Belužić, R.; Imešek, M.; Morrow, C.; Marjanović, M.P.; Bakran-Petricioli, T.; Mikoč, A.; et al. Functional and Structural Characterization of FAU Gene/Protein from Marine Sponge Suberites domuncula. Mar. Drugs 2015, 13, 4179-4196. https://doi.org/10.3390/md13074179
Perina D, Korolija M, Hadžija MP, Grbeša I, Belužić R, Imešek M, Morrow C, Marjanović MP, Bakran-Petricioli T, Mikoč A, et al. Functional and Structural Characterization of FAU Gene/Protein from Marine Sponge Suberites domuncula. Marine Drugs. 2015; 13(7):4179-4196. https://doi.org/10.3390/md13074179
Chicago/Turabian StylePerina, Dragutin, Marina Korolija, Marijana Popović Hadžija, Ivana Grbeša, Robert Belužić, Mirna Imešek, Christine Morrow, Melanija Posavec Marjanović, Tatjana Bakran-Petricioli, Andreja Mikoč, and et al. 2015. "Functional and Structural Characterization of FAU Gene/Protein from Marine Sponge Suberites domuncula" Marine Drugs 13, no. 7: 4179-4196. https://doi.org/10.3390/md13074179
APA StylePerina, D., Korolija, M., Hadžija, M. P., Grbeša, I., Belužić, R., Imešek, M., Morrow, C., Marjanović, M. P., Bakran-Petricioli, T., Mikoč, A., & Ćetković, H. (2015). Functional and Structural Characterization of FAU Gene/Protein from Marine Sponge Suberites domuncula. Marine Drugs, 13(7), 4179-4196. https://doi.org/10.3390/md13074179