1. Introduction
Since ancient times in China, dry calcareous shells have been used for Traditional Chinese Medicine. “Chinese Medicinal Herbal” recorded that sea urchins have the function: “Ruan jian san jie, resolving phlegm, eliminating swelling, expectorate sputum accumulation” [
1]. According to a modern pharmacological study, it was demonstrated that sea urchin extracts have biological effects, such as anti-cancer, antioxidant, anti-leukemia, anti-fatigue and anti-inflammatory effects [
2,
3,
4]. For example, the compound “Bonellinin” extracted from sea urchin gonad could inhibit the growth of cancer cells [
5]. Polyhydroxylated naphthoquinone pigments from the shells of purple sea urchin (
Anthocidaris crassispina) possessed a strong antiradical activity against 1,1-diphenyl-2-picrylhydrazyl (DPPH), superoxide radical anion, and hydrogen peroxide [
6]. For example, polyhydroxylated 1,4-naphthoquinone exhibited dose-dependent antioxidant activities on DPPH radicals [
7,
8] and inhibited the model of ocular allergic inflammation surpassing the reference drug olopatadine [
9]; 6-ethyl-2,3,5,7,8-pentahydroxy-1,4-naphthoquinone (Echinochrome A) existing in sea urchin shells and spines could be used as the auxiliary agent for reducing cardio toxic agent-induced damage [
10].
Sea urchin extracts can have many pharmacological effects, but what are the main chemical components? Sea urchins consist of shells and gonads. The sea urchin shells contain minerals (>90%), proteins, polysaccharides and pigments, while sea urchin gonads consist of, for instance, polysaccharides, fatty acid, and proteins [
11]. To this end, scholars have paid more attention to the pharmacological activities and chemical constituents of sea urchin gonads. Pozharitskaya
et al. found that the fractions of ESD1 and ESD2 extracted from gonads of green sea urchin (
Strongylocentrotus droebachiensis) could be the functional food supplement for the regulation of inflammation and diabetes [
12]. It was shown that polysaccharides from the gonad could inhibit tumor transplant and improve immune activity [
13]. Ke
et al. reported
Strongylocentrotus nudus egg polysaccharides (SEP) displayed antitumor activity mediated by the activation and cytotoxicity of NK cells via TLR2/4
in vivo [
14]. A water-soluble polysaccharide was extracted from sea urchin intestine, which could significantly inhibit the cell growth [
15]. At a dose of 50 μg/mL, the sulfated polysaccharide XB-1 prepared from shells of sea urchin
Hemicentrotus pulcherrimus could reduce the number of HeLa, MCF-7 and A549 cells to 42.08%, 18.42% and 63.20% [
16]. However, there are few reports examining the compositions and inflammatory bioactivities of sea urchin shell polysaccharides.
More than 800 species of sea urchins have been identified in the world, including approximately 100 species in China. In this paper, three common commercial sea urchins, Strongylocentrotus nudus, Anthocidaris crassispina and Glyptocidaris crenularis, were studied from Shandong Peninsula, coastal areas of Zhejiang, Fujian and Guangdong Province, and Liaodong Peninsula, respectively. A quantitative analysis of the composition of shell polysaccharides hydrolysate combined with a chromatographic fingerprint was first established by HPLC-DAD, and the anti-inflammatory activities of polysaccharides of sea urchin shells were reported, which better clarify the structures and anti-inflammatory activities among three species of sea urchin shell.
3. Discussion
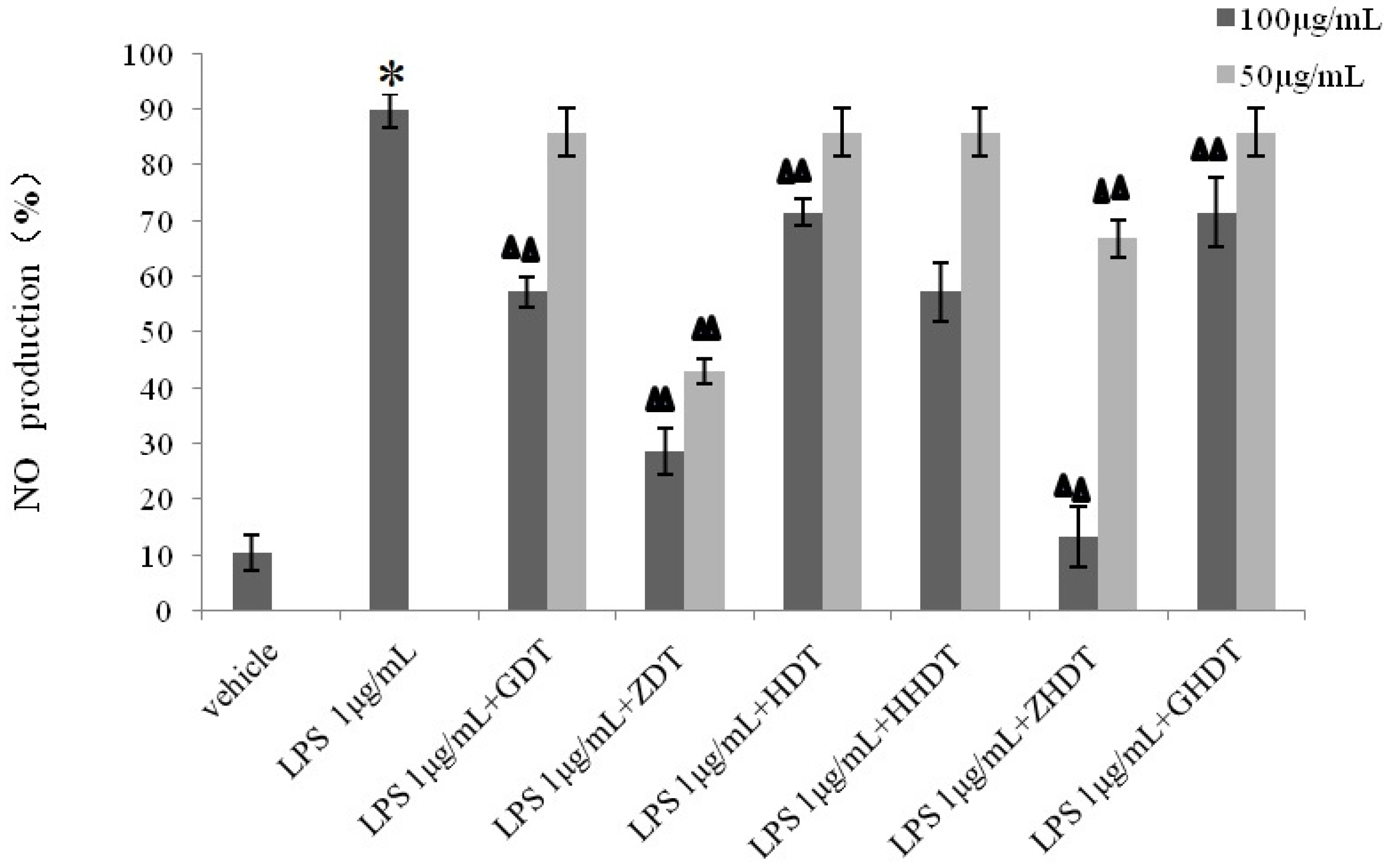
According to the record of an ancient book, sea urchin shells can diminish inflammation and relieve pain. However, there has been no research investigating the anti-inflammatory activity of sea urchin shells. In this paper, an inflammation model of murine macrophage cell line RAW264.7 was first established to explore the activity of three species of sea urchin shell polysaccharides. The experiment demonstrated that the polysaccharide of the purple sea urchin shell and gonad showed an obvious inhibitory effect on RAW264.7 cell growth. Polysaccharides from the purple sea urchin shell and gonad and
Strongylocentrotus nudus gonad had different inhibition actions on the proliferation of RAW264.7 cells stimulated by LPS. The inhibitory activity of purple sea urchin polysaccharides exceeded those of the other sea urchin, and the polysaccharide of
Glyptocidaris crenularis has no anti-inflammatory effect. In experiments measuring the release of NO, we found that every genus of sea urchin shell polysaccharide showed different inhibitory functions: the inhibition rate of shell polysaccharides of
Anthocidaris Crassispina (70%) was greater than the inhibition rate of
Strongylocentrotus nudus shell polysaccharide (40%), which in turn was greater than the inhibition rate of shell polysaccharide of
Glyptocidaris crenularis (20%). Next, we analyzed the monosaccharide compositions of the polysaccharides of three types of sea urchin shells. As shown in
Table 1, the monosaccharide compositions were similar, but the contents of each monosaccharide were different. For example, the amount of fucose and glucuronic acid in
Strongylocentrotus nudus shells exceeded 25% and 5%, respectively. In addition, compared with other sea urchin shells, the amount of glucose of
Glyptocidaris crenularis was larger than others, which reached more than 30%. Thus, we could deduce that differences of the primary monosaccharide composition led to the varying anti-inflammatory activity of the polysaccharide of sea urchin shell.
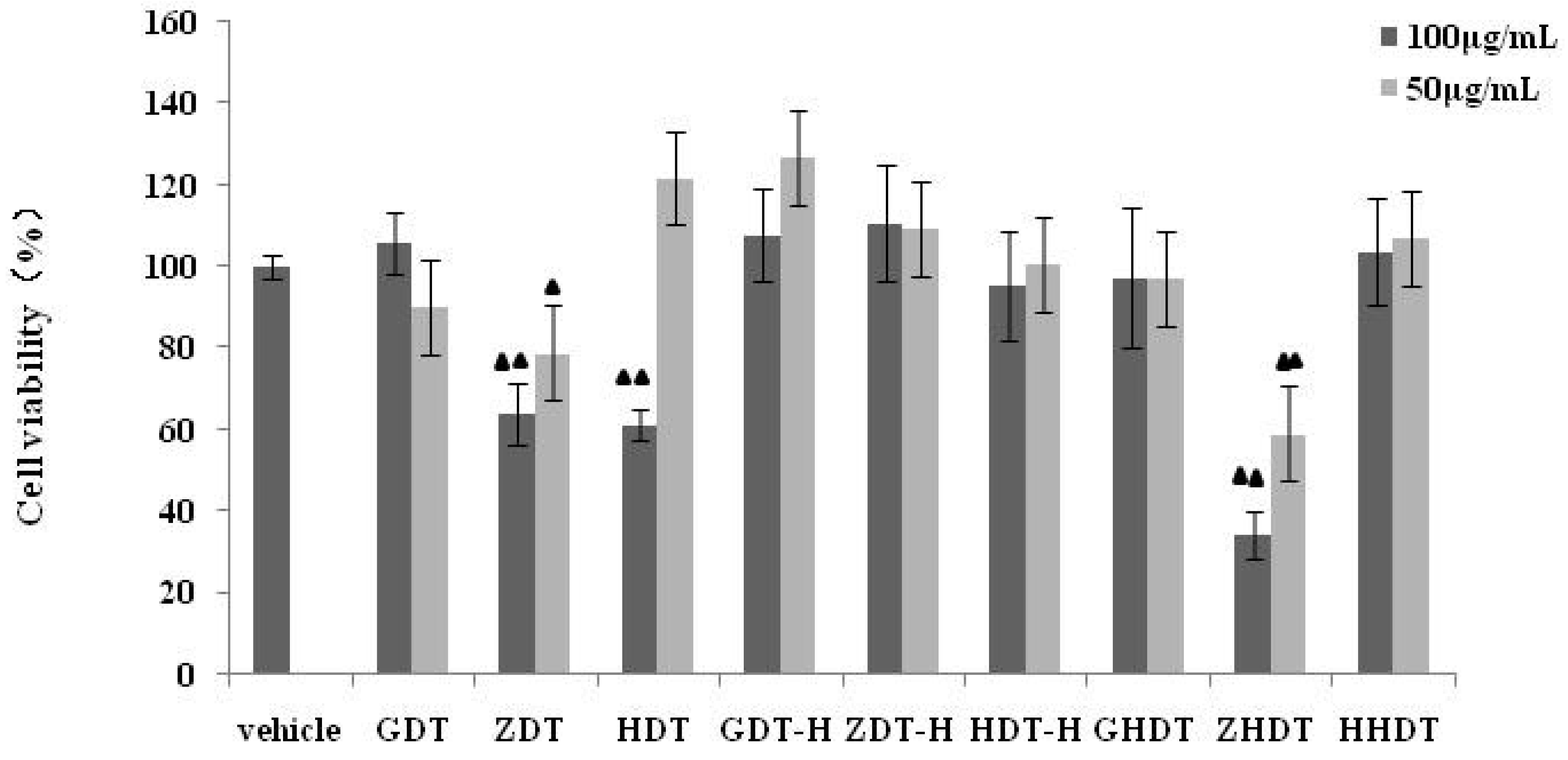
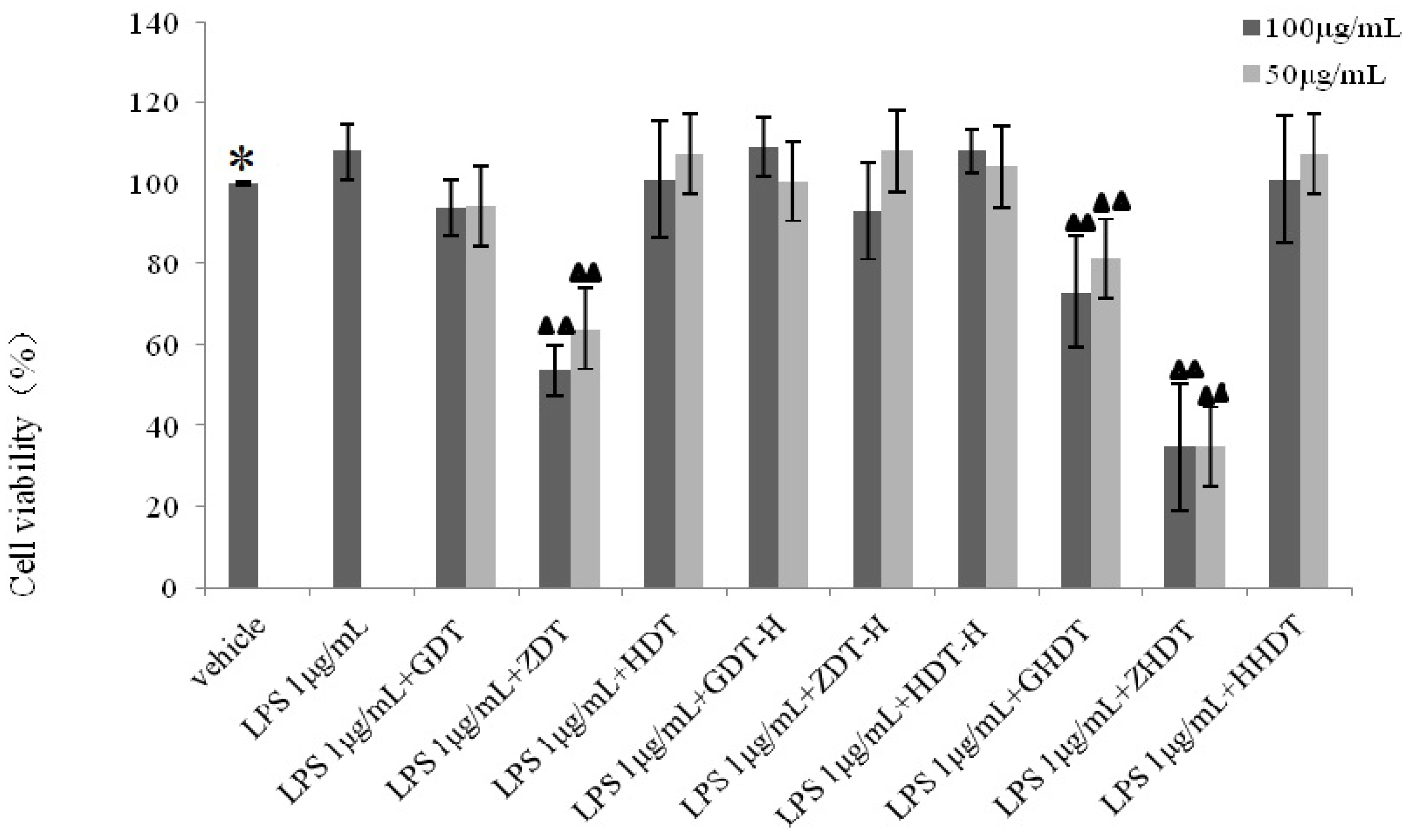
Based on the above experimental results, we summarized the difference of anti-inflammatory activity between the purple sea urchin shell and gonad polysaccharide. As shown in
Figure 6,
Figure 7 and
Figure 8, polysaccharides from the purple sea urchin shell (100 μg/mL) and gonads (100 μg/mL) produced inhibition rates of 40% and 60% on RAW264.7 cells without LPS stimulation. Meanwhile, the purple sea urchin shell polysaccharide and gonad polysaccharide reduced the amount of RAW264.7 cells stimulated by LPS to 50% and 60%, and effectively inhibited the release of NO with inhibition rates of 70% and 80%. However, the polysaccharide hydrolysate of the purple sea urchin shell exhibited no inhibition to RAW264.7 cell proliferation.
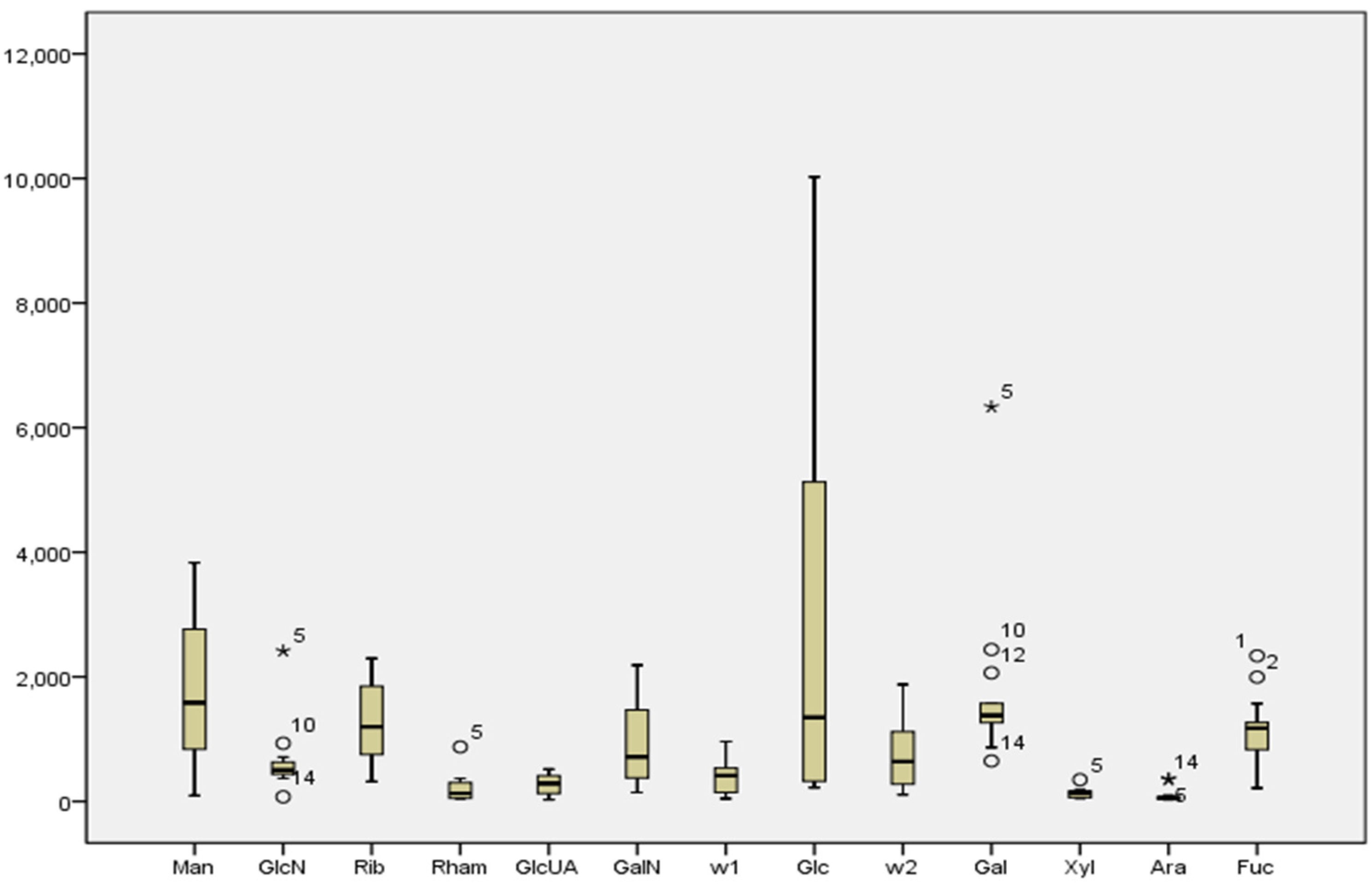
It was determined that compositions of purple sea urchin shell polysaccharide were similar to its gonad polysaccharides, and both of them were neutral heteropolysaccharides. In
Figure 1 and
Figure 5, they contained common monosaccharide fragments, such as Man, Rib, Rham, GalN, Glc, Gal, Ara and Fuc. These common fragments decided the structural similarity between shell polysaccharide and gonad polysaccharide. Shell polysaccharide contained glucosamine and two unknown components. In general, glucosamine plays a role in boosting the growth of bone cells, and therefore mainly exists in the animal shell. Two unknown components need to be further confirmed. According to the literature, CR3, the human macrophage surface polysaccharide receptor, has specificity to mannose, dextran and
N-acetyl-glucosamine. This specific binding can promote the phagocytosis by macrophages and inhibit the generation of the NOS enzyme [
17,
18]. As is widely known, both sea urchin shell and gonad polysaccharides contained glucose and mannose. Therefore, the commonalities of the structure and content of the monosaccharide compositions determine the anti-inflammatory activity, but the differences of their contents explain the differences in anti-inflammatory activity. In addition, the connection of the glycosidic bond and spatial structure between monosaccharide and monosaccharide affected the activities of various species of sea urchin polysaccharides, and the anti-inflammatory activities of the polysaccharide of the purple sea urchin gonad and shell. In experiments of anti-inflammatory activity, we found the polysaccharide of purple sea urchin shell and gonads were cytotoxic to RAW264.7 cells. Determining whether the anti-inflammatory activities of the polysaccharide of the purple sea urchin were produced directly by itself or indirectly by inhibiting cell growth relies on further study of the anti-inflammatory mechanism.
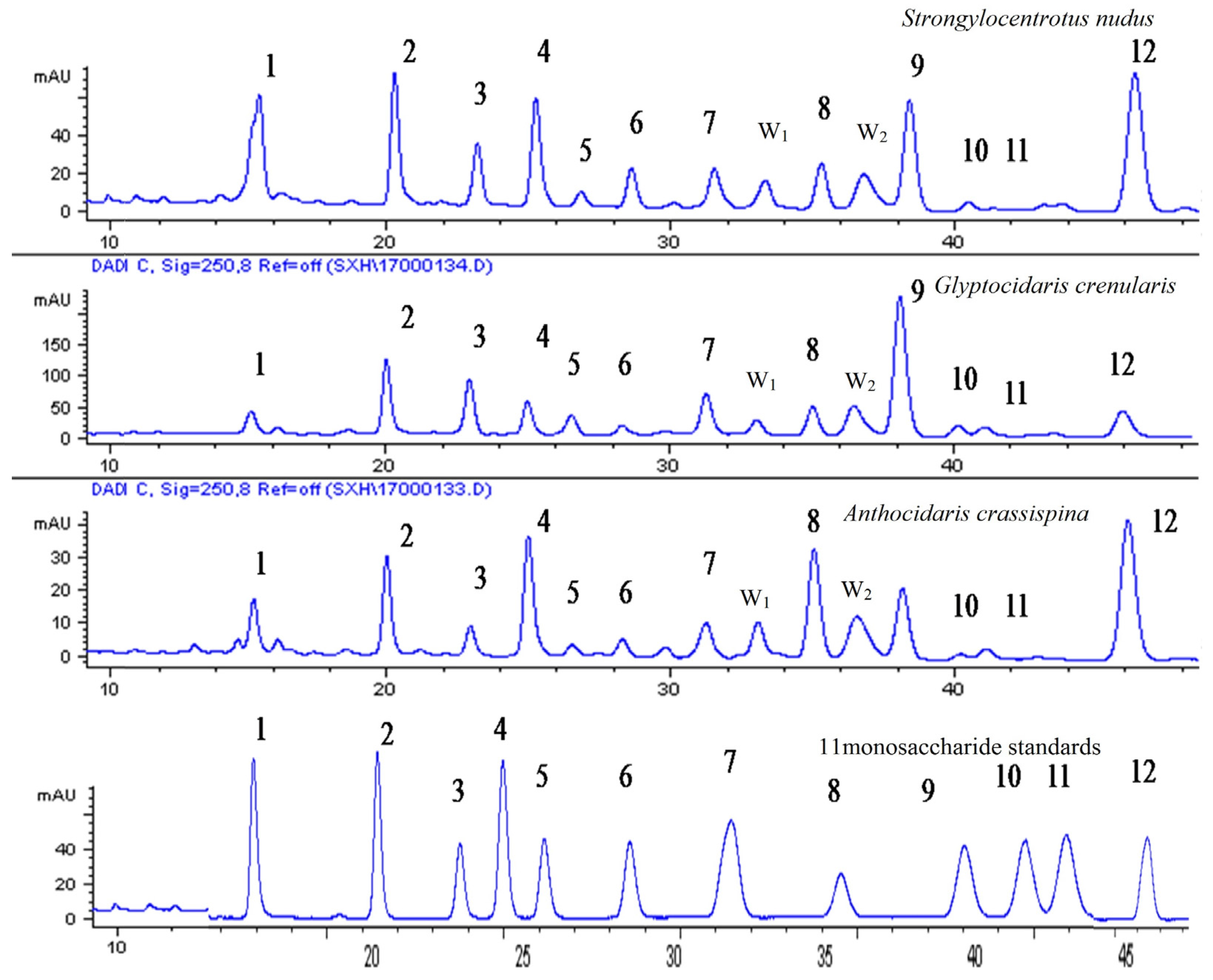
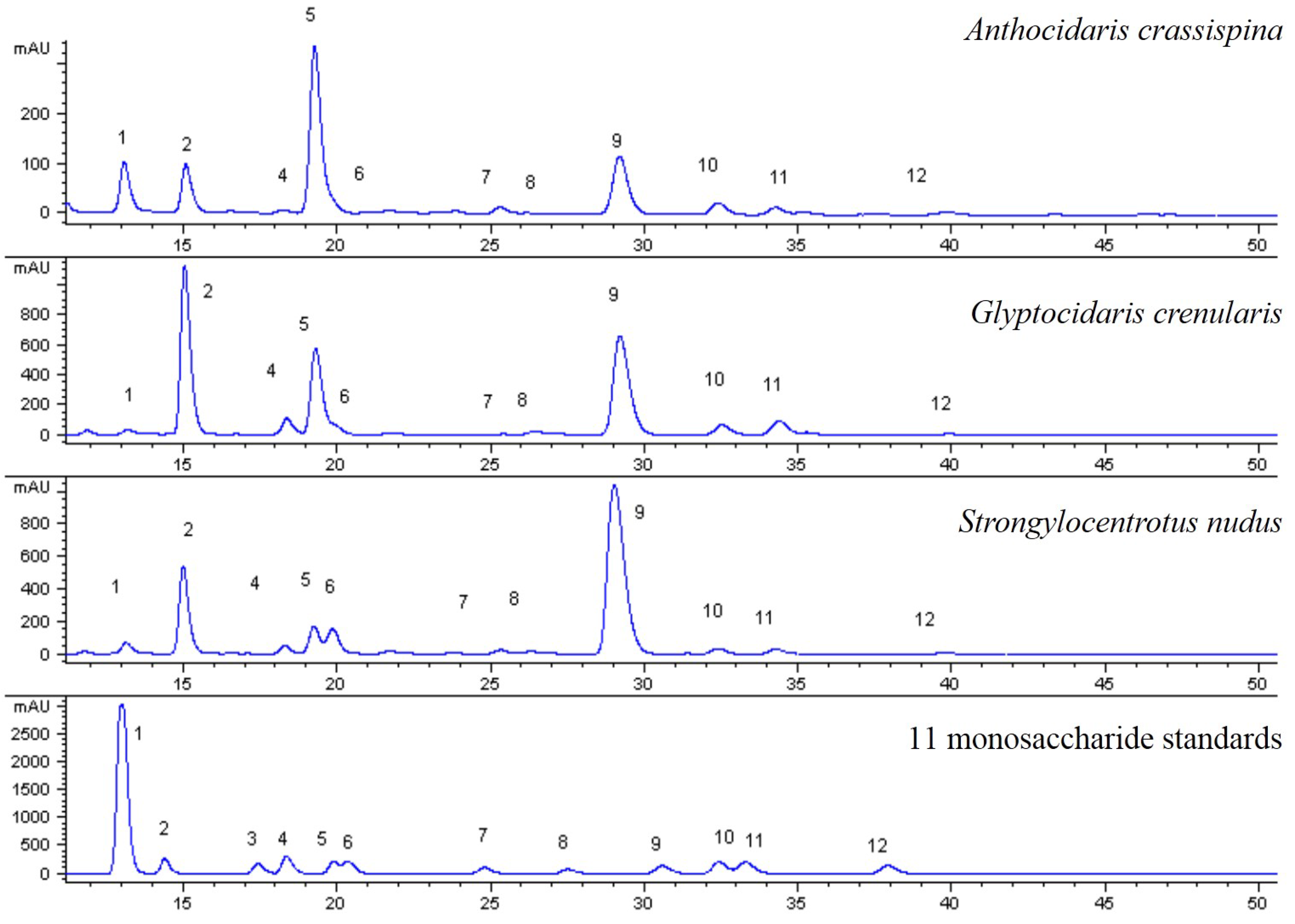
There are many types of sea urchins in the world, but how to distinguish them? This paper established the characteristic fingerprints of three species of sea urchin shell polysaccharide hydrolysate and determined the contents of 11 monosaccharide compositions in 14 batches of sea urchin shells. Then, it was shown that three species of sea urchin shells were consistent with the monosaccharide compositions, but exhibited differences in content. For example, the content of Fuc, Gal and Man of Strongylocentrotus nudus is higher than other compositions; purple sea urchin mainly consisted of Glc, Man and Fuc; the primary monosaccharide compositions of Glyptocidaris crenularis were Glc, Man and Gal. In Strongylocentrotus nudus, the content of fucose exceeded 20%, however, it was below 15% in Anthocidaris crassispina and Glyptocidaris crenularis. Thus, fucose can be used as an indicator for distinguishing among species of sea urchin. Furthermore, there was a gap in the content of glucose between Anthocidaris crassispina and Glyptocidaris crenularis, its content was mostly over 20% in purple sea urchin and below 20% in Glyptocidaris crenularis, so glucose was another index for the sea urchin. Besides, diversities of the structures of polysaccharides were also affected by specificity of sea urchin shells, environmental conditions and very low content of polysaccharides in samples.
4. Experimental Section
4.1. Samples
Fourteen batches of sea urchins were harvested from the north to the south (
Table 2). These samples were identified by Professor Xiaoqi Zeng, Ocean University of China. The collected sea urchins were eviscerated, washed and then air dried. Dried shells were packed and frozen at −20 °C. The voucher specimens (No. 130806, No. 121024, No. 131016) were deposited at the department of Biochemistry and Molecular Biology, Second Military Medical University, Shanghai, China.
Table 2.
Summary of the tested samples.
Table 2.
Summary of the tested samples.
| Sample Number | Species (Family) | Popular Names | Province | Harvesting Time |
|---|
| 1 | Strongylocentrotus nudus | Dalian purple sea urchin or Blackthorn pot | Shandong | 6 August 2013 |
| 2 | Strongylocentrotus nudus | Shandong | 6 August 2013 |
| 3 | Strongylocentrotus nudus | Shandong | 6. August 2013 |
| 4 | Strongylocentrotus nudus | Shandong | 6. August 2013 |
| 5 | Glyptocidaris crenularis | / | Shandong | 25 March 2013 |
| 6 | Anthocidaris crassispina | Sea needle or Sea chestnuts | Zhejiang | 24 September 2012 |
| 7 | Glyptocidaris crenularis | / | Dalian | 24 October 2012 |
| 8 | Glyptocidaris crenularis | / | Dalian | 8 November 2012 |
| 9 | Glyptocidaris crenularis | / | Dalian | 23 November 2012 |
| 10 | Glyptocidaris crenularis | / | Dalian | 10 December 2012 |
| 11 | Glyptocidaris crenularis | / | Shandong | 8 August 2013 |
| 12 | Glyptocidaris crenularis | / | Shandong | 8 August 2013 |
| 13 | Anthocidaris crassispina | Sea needle or Sea chestnuts | Guangdong | 15 August 2013 |
| 14 | Anthocidaris crassispina | Sea needle or Sea chestnuts | Guangdong | 16 October 2013 |
4.2. Chemicals
Acetonitrile (HPLC grade), 3-Methyl-1-phenyl-2-pyrazolin-5-one (PMP, AR), trifluoroacetic acid (TFA, AR), trichloromethane (AR), HCl (AR), and NaOH (AR) were purchased from Sinopharm Chemical Reagent Co., Ltd. (Shanghai, China). Deionized water was obtained from a Hi-tech K-flow water purification system (Shanghai, China). Standard substances including Mannose (d-Man), Glucose (d-Glc), Galactose (d-Gal), Rhamnose (l-Rham), Xylose (d-Xly), Arabinose (l-Ara) were purchased from Sinopharm Chemical Reagent Co., Ltd. (Shanghai, China). Ribose (d-Rib), Fucose (l-Fuc) and Glucuronic acid (d-GlcUA) were purchased from Yuanye Biology Co., Ltd. (Shanghai, China). The purities of all the standards were above 98% as determined by HPLC.
LPS (Escherichia coli 055:B5) was purchased from Sigma. Fetal bovine serum (FBS), Trypsin-EDTA and DMEM with antibiotics (100 U/mL penicillin, 100 μg/mL streptomycin) were purchased from Beijing Solarbia Science and Technology Co., Ltd. (Beijing, China). A CCK-8 (Cell Counting Kit-8) was obtained from Obio Technology Co., Ltd. (Shanghai, China). A Nitric Oxide (NO) assay kit (Chemical NO-assay) was purchased from Nanjing Jiancheng Bioengineering Institute.
4.3. Preparation of Saccharide Derivatization Reagents
Dried sea urchin shells were crushed into powder. Fifty grams of powder was weighed and soaked in 750 mL water, extracted three times with a heating circumfluence method at 70 °C for 1 h each time, and filtered. The filtrate was combined and evaporated to 50 mL in a DK-8 Delectric water bath (Shanghai Jinghong Laboratory Instrument Co., Ltd., Shanghai, China), then 250 mL absolute ethyl alcohol was added for precipitation at room temperature and centrifuged at 5000 rpm for 10 min. The precipitate was dried and stored at 4 °C. Fifty milligrams of powder was accurately weighed and poured into the glass test tube, then added to 1 mL TFA. The mixture was boiled for 6 h with an oil-bath method and then evaporated to dryness by a rotary evaporator (Shanghai Ailang Instruments Co., Ltd., Shanghai, China).The acidolysis product was dissolved with 2 mL water and filtered through a 0.45 μm microporous membrane to remove impurities. Then, 100 μL polysaccharide hydrolysate, 50 μL NaOH (0.2 mol/L) and 40 μL PMP were reacted for 30 min by water bath at 70 °C. After returning to room temperature and neutralization for hydrochloric acid, the redundant derivatization reagent was extracted with the addition of 200 μL chloroform and centrifuged at 8000 rpm for 20 min, filtered through a 0.45 μm microporous membrane to yield the sample solution. Each sample was determined in triplicate.
4.4. Preparation of Standard Solutions
Eleven monosaccharide (Mannose, Glucose, Galactose, Rhamnose, Xylose, Arabinose, Ribose, Fucose, Glucuronic acid, Glucosamine and Galactosamine) standards were accurately weighed and dissolved in volumetric flasks to obtain appropriate stock solutions, respectively. All standard solutions were prepared in water and stored at 4 °C before use.
4.5. Analytical Method
A chromatographic separation was performed on an Agilent 1100 series HPLC-DAD system equipped with a vacuum degasser, a binary pump, a column compartment and a diode array detector (DAD). All compositions of the samples were injected on a CNW™ C18 column (250 mm × 4.6 mm, 5 μm) at 30 °C. The mobile phase was composed of solvent A (acetonitrile) and solvent B (PBS, pH 6.8) with a linear gradient: 0–25 min (A 16%–18%), 25–45 min (A 18%–19%), and 45–50 min (A 19%–19%) with a flow rate of 1 mL/min. The detection wavelength was set at 250 nm and the sample injection volume was 30 μL.
4.6. Method Validation
The performance of the quantitative method used to determine 11 monosaccharide components in sea urchin shells was established through a validation protocol that followed the appendix of China Pharmacopeia (2010 edition, Volume I). The parameters of linearity, precision, repeatability stability, and recovery accuracy were evaluated.
4.6.1. Linearity
These stock solutions were diluted to a series of mixed working solutions with different concentrations. Calibration curves were established based on six levels for each in triplicate. The curves’ regression equations were calculated in the form of y = ax + b. y represented the peak area and x represented the compound concentration.
4.6.2. Precision, Repeatability and Stability
The precision was examined by performing assays with six replicate injections of sample 1. Six replicate sample solutions were independently prepared and analyzed to confirm the repeatability of the method. The stability was assessed by analyzing the same sample solution at 0, 2, 4, 8, 12 and 24 h.
4.6.3. Recovery
The recovery test was investigated with a standard addition method. Three different concentration levels (approximately equivalent to 0.8, 1.0 and 1.2 times the concentration of sample 1) of the mixed standard solution were added into the sample 1 solution.
4.7. Quantitative Determination of 11 Monosaccharide Components in Sea Urchin Shells
Eleven known compositions were determined in 14 batches of the sea urchin shells. The peak area of each component was recorded and the corresponding concentration calculated according to the calibration curve. Each sample was evaluated in triplicate. The content percentages of each component of three species of the sea urchin shell polysaccharide were written in
Table 1.
4.8. Establishment of the HPLC Fingerprints of 14 Batches of the Sea Urchin Shells
The HPLC fingerprints of 14 batches of samples after peak alignment are established by Chromatographic Fingerprint of Traditional Chinese Medicine. The reference fingerprint of different batches of samples was generated by similarity analysis software for Chromatographic Fingerprint of Traditional Chinese Medicine, which was recommended by State Food and Drug Administration of China (SFDA).
4.9. Anti-Inflammatory Activity
4.9.1. Cell Culture
The murine macrophage cell line RAW264.7 was cultured in the flask in Dulbecco’s modified Eagle’s medium (DMEM) with 10% fetal bovine serum (FBS). The cells were incubated at 37 °C under 5% CO2 humidified air.
4.9.2. Cell Viability Assay
The cells were seeded in 96-well plates for 24 h and the old medium changed with various concentrations of the sea urchin shell and gonads polysaccharides (100 μg/mL, 50 μg/mL) for 48 h at 37 °C. Using the cell counting kit-8 (CCK-8), CCK-8 solution was added to 96 wells and the cells were incubated for half an hour. The optical density was read at 450 nm with an ELISA reader (550 ELISA Reader, Bio-Rad Co., Ltd., Hercules, CA, USA).
4.9.3. Cell Viability Assay after LPS-Induced Raw264.7
The cells were plated in 96-well plates for 24 h. The medium was discarded and LPS (1 μg/mL) was added to every well except for the vehicle group. Next, the cells were incubated for 48 h at 37 °C. The optical density was read at 450 nm with an ELISA reader.
4.9.4. Determination of NO Production
The cells were plated in 24-well plates for 24 h. The medium was discarded. Cells were incubated with LPS (1 μg/mL) and the varying concentrations of the sea urchin shell polysaccharide (100 μg/mL, 50 μg/mL) solutions for 36 h. The supernatant was treated as the Nitric Oxide (NO) assay kit. Then the absorbance was measured at 570 nm with an ELISA reader. The assays were performed in triple and the mean and standard deviation (SD) were calculated. The amount of NO production was calculated by the following formula: NO (μmol/L) = (Ai − Ao)/(As − Ao) × 20 (μmol/L) × d, where Ao is the absorbance with the vehicle group, Ai is absorbance with the sample group, As is the absorbance with the NO standard solution (20 μmol/L), and d is the diluted multiples. The percentage of NO production was calculated by the following formula: NO% = (Ai − Ao)/(AL − Ao) × 100%, where Ao is the absorbance with the vehicle group, and Ai is absorbance with the sample group, AL is the absorbance with the LPS group.
4.10. Data Statistics
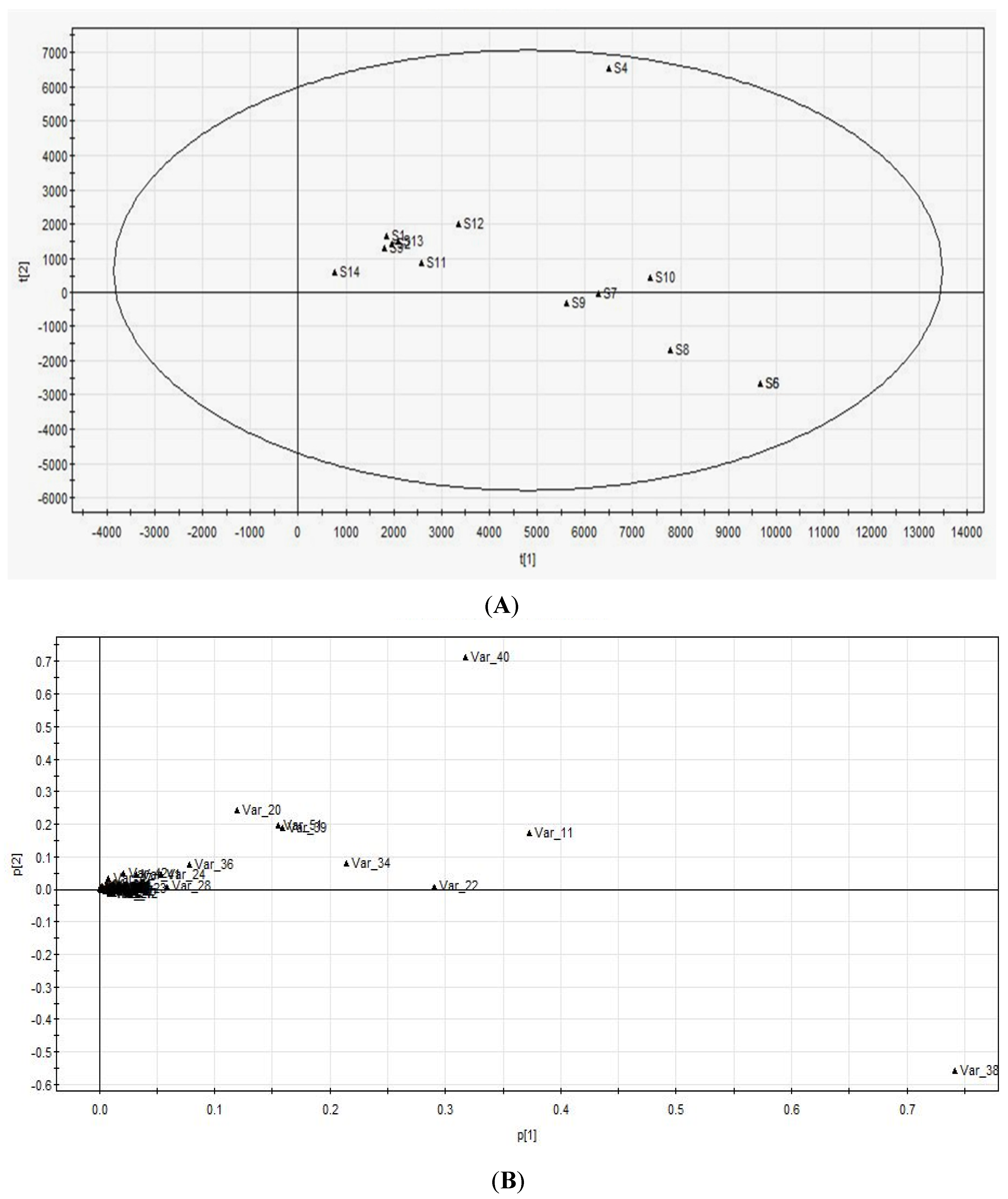
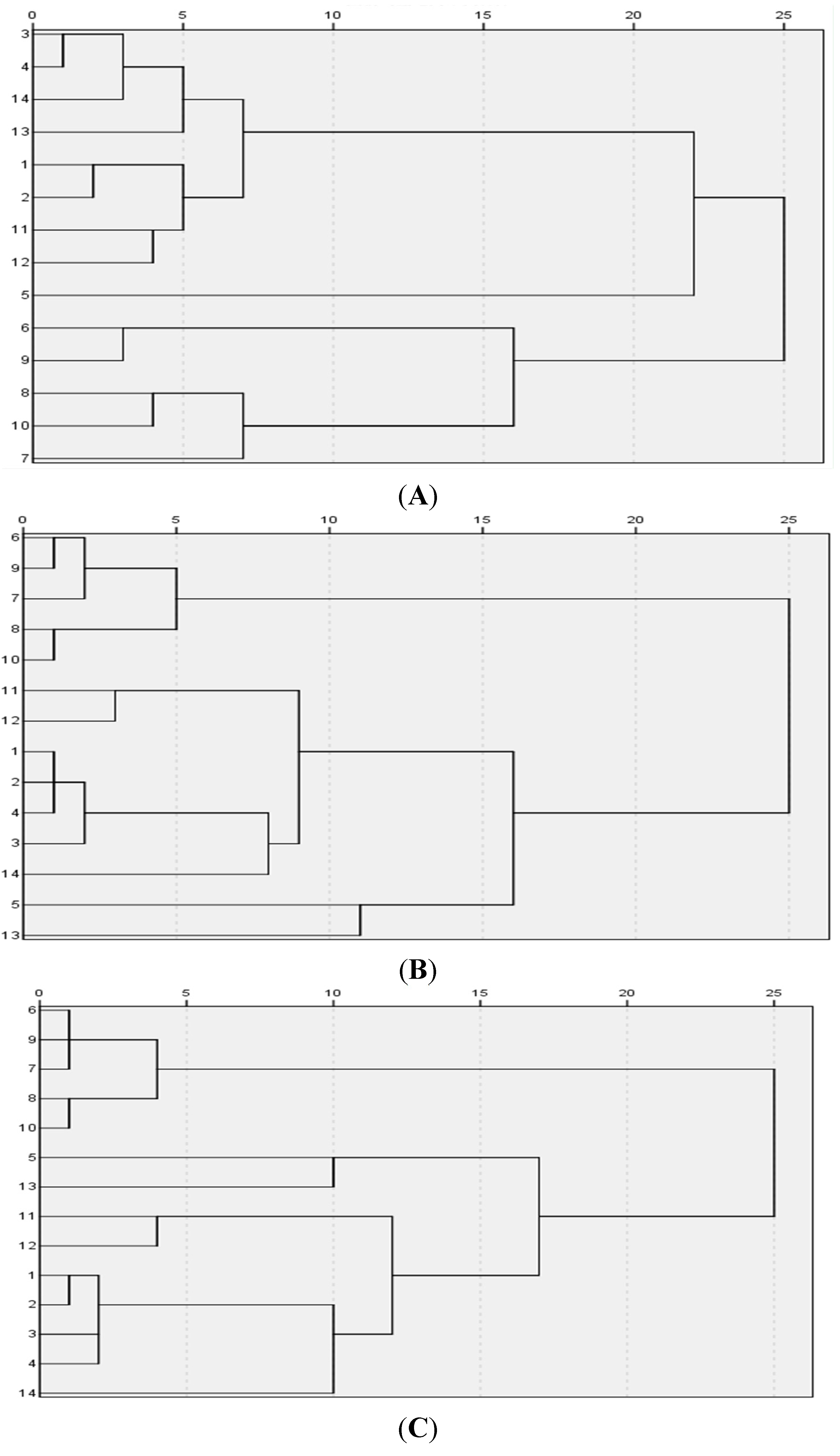
The relative standard deviations (RSD) of eleven components were calculated in Excel. Boxplot and Hierarchical cluster analyses were performed by IBM SPSS Statistics. PCA was performed by SIMCA-P12.0.


 and
and 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






