
Aeroplysinin-1, a Sponge-Derived Multi-Targeted Bioactive Marine Drug



Abstract
:1. Introduction: Chemical Defense of Sponges
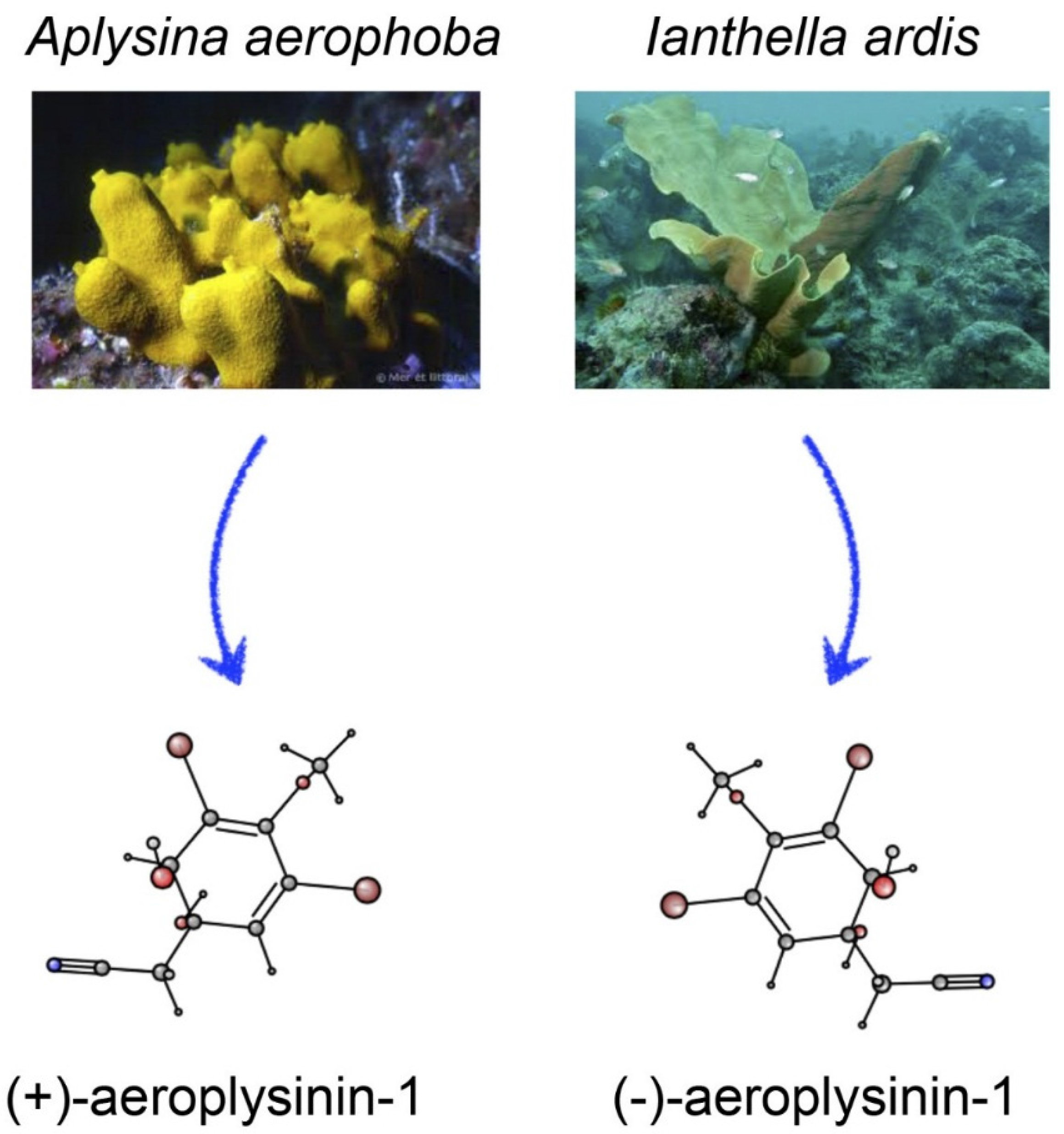
2. Order Verongida
3. Chemical Structure and Properties of Aeroplysinin-1

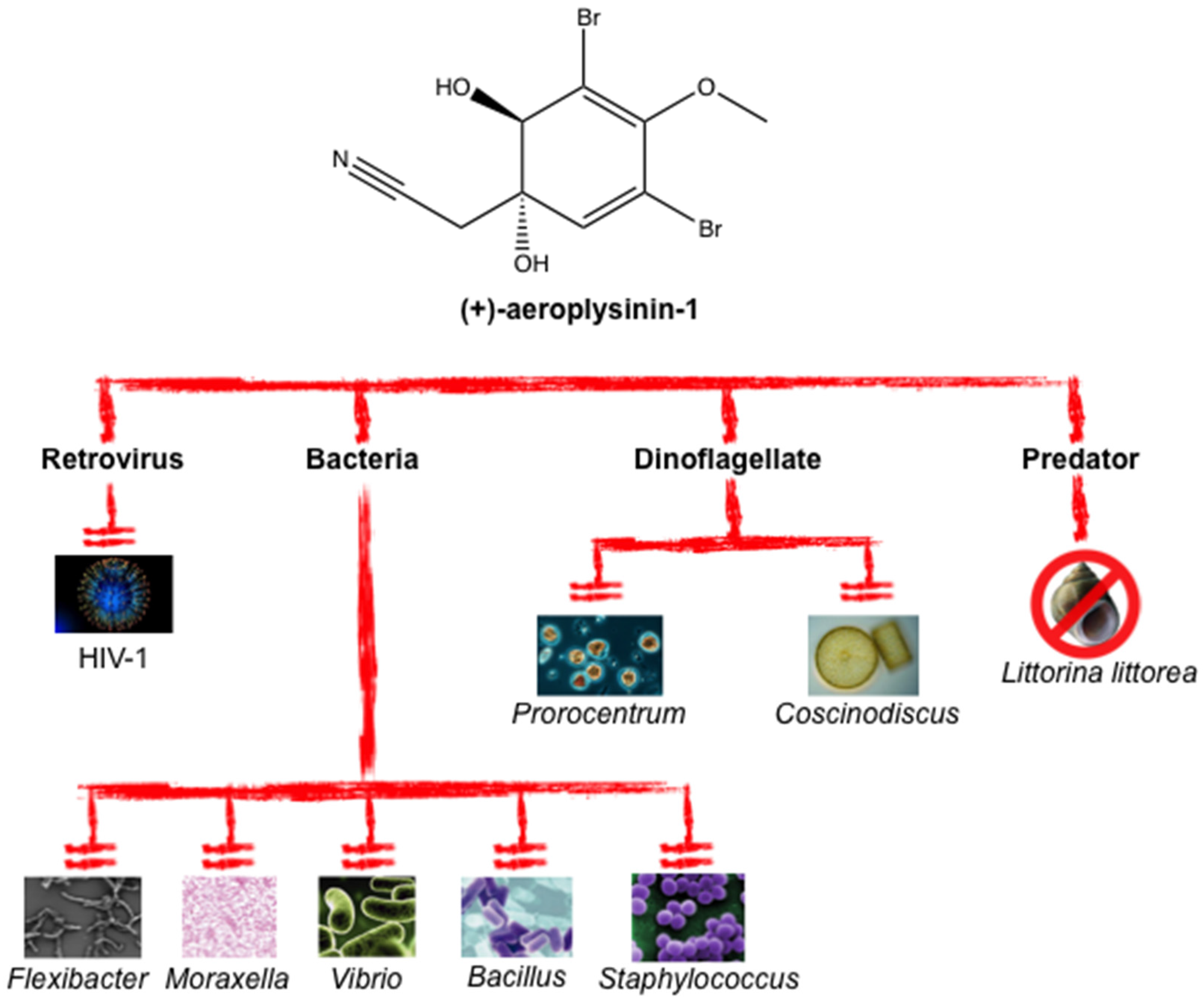
4. Reported Antibiotic, Antimicrobial and Antiviral Properties of Aeroplysinin-1 as a Defense Metabolite

{kind=link}
{kind=link}
{kind=link}
| Microorganism Species | IC50 (μM) | Reference |
|---|---|---|
| P. phosphoreum | 3.5 | [32] |
| C. wailesii | 5.6 | [32] |
| P. minimum | 7.0 | [32] |
| HIV-1 | 14.6 | [34] |

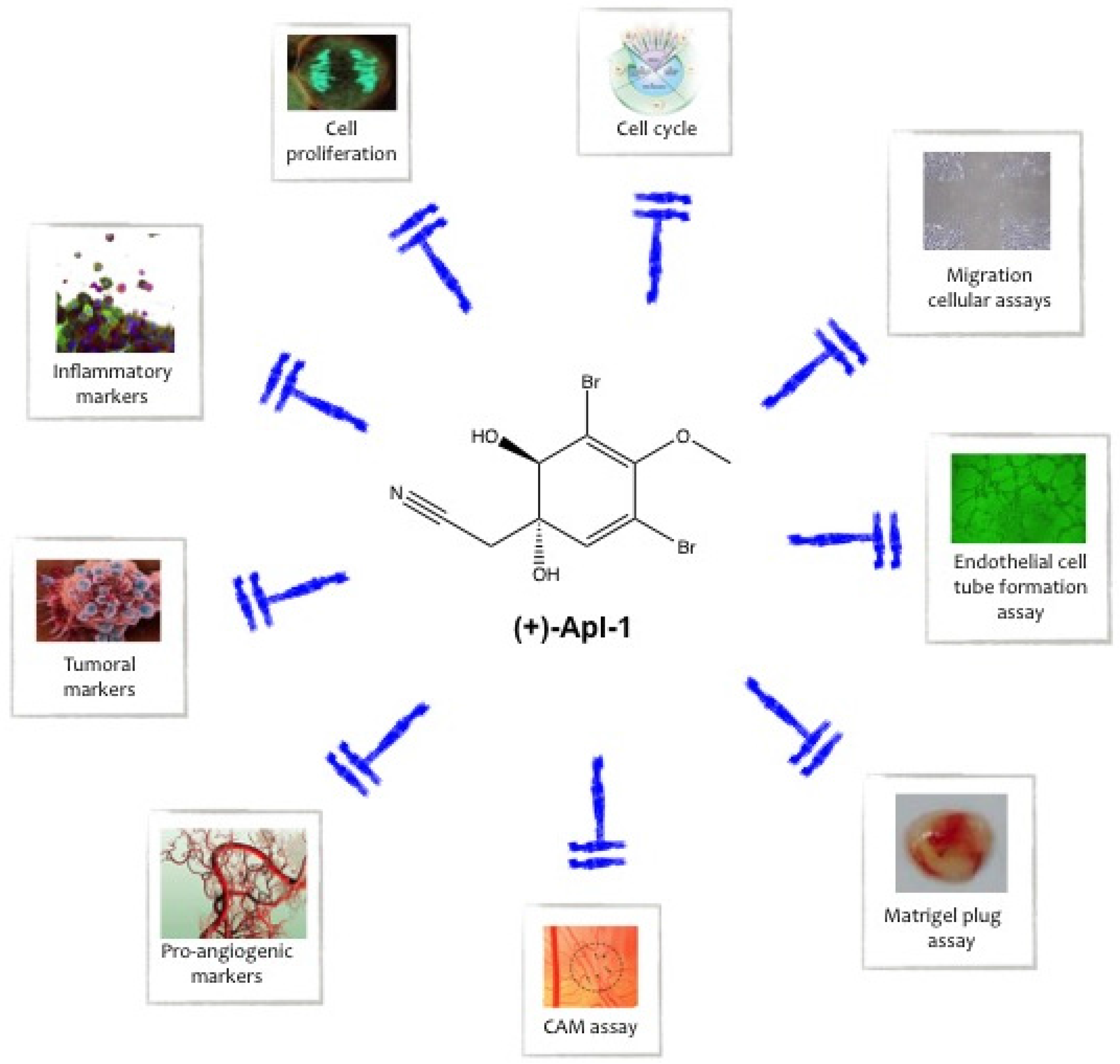
5. Aeroplysinin-1 as an Anti-Inflammatory Compound
| Cell Line | Treatment (μM) | Target | Effect | Activity | Reference |
|---|---|---|---|---|---|
| EVLC-2 | 2.5 | MMP-2 | Decrease | Anti-angiogenic | [38] |
| HUVEC | 2.5 | MMP-2 | Decrease | Anti-angiogenic | [38] |
| HUVEC | 10 | MCP-1 | Decrease | Anti-inflammatory | [38] |
| HUVEC | 10 | TSP-1 | Decrease | Anti-inflammatory | [38] |
| HUVEC | 10 | COX-2 | Decrease | Anti-inflammatory | [38] |
| HUVEC | 20 | Il-1α | Decrease | Anti-inflammatory | [38] |
| HUVEC | 20 | MMP-1 | Decrease | Anti-inflammatory | [38] |
| RF-24 | 2.5 | MMP-2 | Decrease | Anti-angiogenic | [38] |
| BAEC | 10 | Cleaved lamin-A | Increase | Apoptogenic | [39] |
| BAEC | 10 | Caspase-2, -3, -8, -9 | Increase | Apoptogenic | [39] |
| BAEC | 10 | Cytochrome C | Increase in cytoplasm | Apoptogenic | [39] |
| HUVEC | 10 | p-Bad | Increase | Apoptogenic | [39] |
| BAEC | 3 | MMP-2 | Decrease | Anti-angiogenic | [40] |
| BAEC | 3 | PA | Decrease | Anti-angiogenic | [40] |
| BAEC | 3 | PAI | Increase | Anti-angiogenic | [40] |
6. Aeroplysinin-1 as an Anti-Angiogenic Compound
| Endothelial Cell Line | IC50 (μM) | Reference |
|---|---|---|
| HUVEC | 4.7 | [38] |
| EVLC-2 | 3.0 | [38] |
| RF-24 | 2.8 | [38] |
| HMEC | 2.6 | [38] |
| BAEC | 2.1 | [40] |
7. Aeroplysinin-1 as an Anti-Tumor Compound
| Tumor cell line | IC50 (μM) | Reference |
|---|---|---|
| HT-1080 | 2.3 | 40 |
| HTC-116 | 4.7 | 40 |
| HeLa | 3.0 | 43 |
| THP-1 | 10.0 | 45 |
| NOMO-1 | 17.0 | 45 |
| HL-60 | 5.0 | 45 |
8. Is (+)-Aeroplysinin-1 a Receptor Tyrosine Kinase Inhibitor?
9. Conclusion: Potential and Perspectives of (+)-Aeroplysinin-1 as a Novel Multi-Targeted Drug

Acknowledgments
Author Contributions
Conflict of Interest
References
- Paul, V.J. Activation of chemical defenses in the tropical green algae Halimeda spp. J. Exp. Mar. Biol. Ecol. 1992, 160, 191–203. [Google Scholar] [CrossRef]
- Engel, S.; Jensen, P.R.; Fenical, W. Chemical ecology of marine microbial defense. J. Chem. Ecol. 2002, 28, 1971–1985. [Google Scholar] [CrossRef] [PubMed]
- Walters, K.D.; Pawlik, J.R. Is there a trade-off between wound-healing and chemical defenses among Caribbean reef sponges? Integr. Comp. Biol. 2005, 45, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Green, G. Ecology of toxicity in marine sponges. Mar. Biol. 1977, 40, 207–215. [Google Scholar] [CrossRef]
- Koopmans, M.; Martens, D.; Wijffels, R.H. Towards commercial production of sponge medicines. Mar. Drugs 2009, 7, 787–802. [Google Scholar] [CrossRef] [PubMed]
- Unson, M.D.; Holland, N.D.; Faulkner, D.J. A brominated secondary metabolite synthesized by the cyanobacterial symbiont of a marine sponge and accumulation of the crystalline metabolite in the sponge tissue. Mar. Biol. 1994, 119, 1–11. [Google Scholar] [CrossRef]
- Thoms, C.; Ebel, R.; Proksch, P. Activated chemical defense in Aplysina sponges revisited. J. Chem. Ecol. 2006, 32, 97–123. [Google Scholar] [CrossRef] [PubMed]
- Li, J.W.; Vederas, J.C. Drug discovery and natural products: End of an era or an endless frontier? Science 2009, 325, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A.L.; Clark, R.L.; Mackay, S.P.; Johnston, B.F. Current strategies for drug discovery through natural products. Exp. Opin. Drug Dis. 2010, 5, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Wohlleben, W.; Pelzer, S. New compounds by combining “Modern” genomics and “Old-Fashioned” mutasynthesis. Chem. Biol. 2002, 9, 1163–1164. [Google Scholar] [CrossRef]
- Datta, D.; Nath Talapatra, S.; Swarnakar, S. Bioactive compounds from marine invertebrates for potential medicines—An overview. Int. Lett. Nat. Sci. 2015, 7, 42–61. [Google Scholar] [CrossRef]
- Laport, M.S.; Santos, O.C.S.; Muricy, G. Marine sponges: Potential sources of new antimicrobial drugs. Curr. Pharm. Biotechnol. 2009, 10, 86–105. [Google Scholar] [CrossRef] [PubMed]
- Kunze, K.; Niemann, H.; Ueberlein, S.; Schulze, R.; Ehrlich, H.; Brunner, E.; Proksch, P.; Van Pée, K.H. Brominated skeletal components of the marine demosponges, Aplysina cavernicola and Ianthella basta: Analytical and biochemical investigations. Mar. Drugs 2013, 11, 1271–1287. [Google Scholar] [CrossRef] [PubMed]
- Diaz, M.C.; Thacker, R.W.; Redmond, N.E.; Matterson, K.O.; Collins, A.G. Phylogenetic novelties and geographic anomalies among tropical verongida. Integr. Comp. Biol. 2013, 53, 482–494. [Google Scholar] [CrossRef] [PubMed]
- Erwin, P.M.; Thacker, R.W. Phylogenetic analyses of marine sponges within the order Verongida: A comparison of morphological and molecular data. Invertebr. Biol. 2007, 126, 220–234. [Google Scholar] [CrossRef]
- Ivanova, E.P.; Kuznetsova, T.A.; Mikhailov, V.V. Antifungal compounds produced by bacterial populations of the Sea Sponge Verongia sp. Russ. J. Mar. Biol. 2000, 26, 118–119. [Google Scholar] [CrossRef]
- Muscholl-Silberhorn, A.; Thiel, V.; Imhoff, J.F. Abundance and bioactivity of cultured sponge-associated bacteria from the Mediterranean Sea. Microb. Ecol. 2008, 55, 94–106. [Google Scholar] [CrossRef] [PubMed]
- Gerçe, B.; Schwartz, T.; Voigt, M.; Rühle, S.; Kirchen, S.; Putz, A.; Proksch, P.; Obst, U.; Syldatk, C.; Husamann, R. Morphological, bacterial, and secondary metabolite changes of Aplysina aerophoba upon long-term maintenance under artificial conditions. Microb. Ecol. 2009, 58, 865–878. [Google Scholar] [CrossRef] [PubMed]
- Turon, X.; Becerro, M.; Uriz, M.J. Distribution of brominated compounds within the sponge Aplysina aerophoba: Coupling of X-ray microanalysis with cryofixation techniques. Cell Tissue Res. 2000, 301, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, A. Temporal variation of the microbial community associated with the mediterranean sponge Aplysina aerophoba. FEMS Microbiol. Ecol. 2001, 38, 105–113. [Google Scholar] [CrossRef]
- Ciminiello, P.; Fattorusso, E.; Forino, M.; Magno, S.; Pansini, M. Chemistry of verongida sponges VIII—Bromocompounds from mediterranean sponges Aplysina aerophoba and Aplysina cavernicola. Tetrahedron 1997, 53, 6565–6572. [Google Scholar] [CrossRef]
- Kochanowska, A.J.; Rao, K.V.; Childress, S.; El-Alfy, A.; Matsumoto, R.R.; Kelly, M.; Stewart, G.S.; Sufka, K.J.; Hamann, M.T. Secondary metabolites from three Florida sponges with antidepressant activity. J. Nat. Prod. 2008, 71, 186–189. [Google Scholar] [CrossRef] [PubMed]
- Sacristán-Soriano, O.; Banaigs, B.; Becerro, M.A. Temporal trends in the secondary metabolite production of the sponge Aplysina aerophoba. Mar. Drugs 2012, 10, 677–693. [Google Scholar] [CrossRef] [PubMed]
- Ebel, R.; Brenzinger, M.; Kunze, A.; Gross, H.J.; Proksch, P. Wound activation of protoxins in marine sponge Aplysina aerophoba. J. Chem. Ecol. 1997, 23, 1451–1462. [Google Scholar] [CrossRef]
- Lira, N.S.; Carneiro-Montes, R.; Fechine-Tavares, J.; Sobral da Silva, M.; da Cunha, E.; Filgueiras, P.; Rodrigues, L.C.; da Silva Dias, C.; Barbosa-Fiho, J.M. Brominated compounds from marine sponges of the genus Aplysina and a compilation of their 13C NMR spectral data. Mar. Drugs 2011, 9, 2316–2368. [Google Scholar] [CrossRef] [PubMed]
- Teeyapant, R.; Proksch, P. Biotransformation of brominated compounds in the marine sponge Verongia aerophoba—Evidence for an induced chemical defense? Naturwissenschaften 1993, 80, 369–370. [Google Scholar] [CrossRef]
- Lipowicz, B.; Hanekop, N.; Schmitt, L.; Proksch, P. An aeroplysinin-1 specific nitrile hydratase isolated from the marine sponge Aplysina cavernicola. Mar. Drugs 2013, 11, 3046–3067. [Google Scholar] [CrossRef] [PubMed]
- Fulmor, W.; Van Lear, G.E.; Morton, G.O.; Mills, R.D. Isolation and absolute configuration of the aeroplysinin I enantiomorphic pais from Ianthella ardis. Tetrahedron. Lett. 1970, 52, 4551–4552. [Google Scholar] [CrossRef]
- Fattorusso, E.; Minale, L.S.G. Aeroplysinin-1, an antibacterial bromo-compound from the sponge Verongia aerophoba. J. Chem. Soc. 1972, 1, 16–18. [Google Scholar] [CrossRef]
- Andersen, R.J.; Faulkner, D.J. Synthesis of aeroplysinin-1 and related compounds. J. Am. Chem. Soc. 1975, 97, 936–937. [Google Scholar] [CrossRef]
- Nieto-Ortega, B.; Casado, J.; Blanck, E.W.; López-Navarrete, J.; Quesada, A.R.; Ramírez, F.J. Raman optical activity spectra and conformational elucidation of chiral drugs. The case of the antiangiogenic aeroplysinin-1. J. Phys. Chem. A 2011, 115, 2752–2755. [Google Scholar] [CrossRef] [PubMed]
- Weiss, B.; Ebel, R.; Elbrächter, M.; Kirchner, M.; Proksch, P. Defense metabolites from the marine sponge Verongia aerophoba. Biochem. Syst. Ecol. 1996, 24, 1–12. [Google Scholar] [CrossRef]
- Teeyapant, R.; Woerdenbag, H.J.; Kreis, P.; Hacker, J.; Wray, V.; Witte, L.; Proksch, P. Antibiotic and cytotoxic activity of brominated compounds from the marine sponge Verongia aerophoba. Z. Naturforsch. C 1993, 48, 939–945. [Google Scholar] [PubMed]
- Gómez-Archila, L.; Zapata, W.; Galeano, E.; Martínez, A.; Díaz, F.J.; Rugeles, M.T. Bromotyrosine derivatives from marine sponges inhibit the HIV-1 replication in vitro. VITAE. Rev. Fac. Química Farm. 2014, 21, 114–125. [Google Scholar]
- Aderem, A.; Ulevitch, R.J. Toll-like receptors in the induction of the innate immune response. Nature 2000, 406, 782–787. [Google Scholar] [CrossRef] [PubMed]
- Fulkerson, P.C.; Rothenberg, M.E. Targeting eosinophils in allergy, inflammation and beyond. Nat. Rev. Drug Discov. 2013, 12, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Poveda, B.; García-Vilas, J.A.; Cárdenas, C.; Melgarejo, E.; Quesada, A.R.; Medina, M.A. The brominated compound aeroplysinin-1 inhibits proliferation and the expression of key pro-inflammatory molecules in human endothelial and monocyte cells. PLoS ONE 2013, 8, e55203. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Poveda, B.; Rodríguez-Nieto, S.; García-Caballero, M.; Medina, M.A.; Quesada, A.R. The antiangiogenic compound aeroplysinin-1 induces apoptosis in endothelial cells by activating the mitochondrial pathway. Mar. Drugs 2012, 10, 2033–2046. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Nieto, S.; González-Iriarte, M.; Carmona, R.; Muñoz-Chápuli, R.; Medina, M.A.; Quesada, A.R. Antiangiogenic activity of aeroplysinin-1, a brominated compound isolated from a marine sponge. FASEB J. 2001, 16. [Google Scholar] [CrossRef]
- Rodríguez-Caso, L.; Reyes-Palomares, A.; Sánchez-Jiménez, F.; Quesada, A.R.; Medina, M.A. What is knwon on angiogenesis-related rare diseases? A systematic review of literature. J. Cell. Mol. Med. 2012, 16, 2872–2893. [Google Scholar] [CrossRef] [PubMed]
- Córdoba, R.; Salvaro-Tormo, N.; Fernñandez-Medarde, A.; Plumet, J. Angiogenic versus cytotoxic activity in analogues of aeroplysinin-1. Bioorg. Med. Chem. 2007, 15, 5300–5315. [Google Scholar] [CrossRef] [PubMed]
- Kreuter, M.H.; Bernd, A.; Holzmann, H.; Müller.Klieser, W.; Maidhof, A.; Weissmann, N.; Kljajic, Z.; Batel, R.; Schröder, H.C.; Müller, W.E.G. Cytostatic activity of aeroplysinin-1 against lymphoma and epithelioma cells. Z. Naturforsch. C 1989, 44, 680–688. [Google Scholar] [PubMed]
- Kreuter, M.H.; Leake, R.E.; Rinaldi, F.; Müller-Klieser, W.; Maidhof, A.; Müller, W.E.; Schröder, H.C. Inhibition of intrinsic protein tyrosine kinase activity of EGF-receptor kinase complex from human breast cancer cells by the marine sponge metabolite (+)-aeroplysinin-1. Comp. Biochem. Physiol. B 1990, 97, 151–158. [Google Scholar] [CrossRef]
- Stuhldreier, F.; Kassel, S.; Schumacher, L.; Wesselborg, S.; Proksch, P.; Fritz, G. Pleiotropic effects of spongean alkaloids on mechanisms of cell death, cell cycle progression and DNA damage response (DDR) of acute myeloid leukemia (AML) cells. Cancer Lett. 2015, 361, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, K.; Ito, F. Target Therapy for Cancer: Anti-cancer drugs targeting growth-factor signaling molecules receptor tyrosine kinases and targeted cancer therapeutics. Biopharm. Bull. 2011, 34, 1774–1780. [Google Scholar] [CrossRef]
- Hinterding, K.; Knebel, A.; Herrlich, P.; Waldmann, H. Synthesis and biological evaluation of aeroplysinin analogues: A new class of receptor tyrosine kinase inhibitors. Bioorg. Med. Chem. 1998, 6, 1153–1162. [Google Scholar] [CrossRef]
- Koulman, A.; Proksch, P.; Rainer, E.; Beekman, A.C.; van Uden, W.; Konings, A.W.T.; Pedersen, J.A.; Pras, N.; Woerdenbag, J. Cytotoxicity and mode of action aeroplysinin-1 and a related dienone from the sponge Aplysina aerophoba. J. Nat. Prod. 1996, 59, 591–594. [Google Scholar] [CrossRef] [PubMed]
- Perović-Ottstadt, S.; Adell, T.; Proksch, P.; Wiens, M.; Korzhev, M.; Gamulin, V.; Müller, I.M.; Müller, W.E.G. A (1→3)-β-d-glucan recognition protein from the sponge Suberites domuncula. Mediated activation of fibrinogen-like protein and epidermal growth factor gene expression. Eur. J. Biochem. 2004, 271, 1924–1937. [Google Scholar]
- (+)-Aeroplysinin-1 (CAS 28656-91-9). Available online: http://www.scbt.com/es/datasheet-202445-aeroplysinin-1.html (accessed on 11 October 2015).
- Martínez-Poveda, B.; Quesada, A.R.; Medina, M.A. (+)-Aeroplysinin-1 inhibits Akt and ERK pathway signaling selectively in endothelial cells. Integr. Cancer Sci. Ther. 2015, 2, 181–184. [Google Scholar]
- Muñoz-Chápuli, R.; Quesada, A.R.; Medina, M.A. Angiogenesis and signal transduction in endothelial cells. Cell. Mol. Life Sci. 2004, 61, 2224–2243. [Google Scholar] [CrossRef] [PubMed]
- García-Godoy, M.J.; López-Camacho, E.; García-Nieto, J.; Aldana-Montes, A.J. Solving molecular docking problems with multi-objective metaheuristics. Molecules 2015, 20, 10124–10183. [Google Scholar] [CrossRef] [PubMed]
- Bhatnagar, I.; Kim, S.K. Immense essence of excellence: Marine microbial bioactive compounds. Mar. Drugs 2010, 8, 2673–2701. [Google Scholar] [CrossRef] [PubMed]
- Imhoff, J.F.; Labes, A.; Wiese, J. Bio-mining the microbial treasures of the ocean: New natural products. Biotechnol. Adv. 2011, 29, 468–482. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.E.; González-Iriarte, M.; Barrero, A.F.; Salvador-Tormo, N.; Muñoz-Chápuli, R.; Medina, M.A.; Quesada, A.R. Study of puupehenone and related compounds as inhibitors of angiogenesis. Int. J. Cancer 2004, 110, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Poveda, B.; Quesada, A.R.; Medina, M.A. The anti-angiogenic 8-epipuupehedione behaves as a potential anti-leukemic compound against HL-60 cells. J. Cell. Mol. Med. 2008, 12, 701–706. [Google Scholar] [CrossRef] [PubMed]
- García-Caballero, M.; Marí-Beffa, M.; Cañedo, L.; Medina, M.A.; Quesada, A.R. Toluquinol, a marine fungus metabolite, is a new angiosuppresor that interferes the Akt pathway. Biochem. Pharmacol. 2013, 85, 1727–1740. [Google Scholar] [CrossRef] [PubMed]
- García-Caballero, M.; Cañedo, L.; Fernández-Medarde, A.; Medina, M.A.; Quesada, A.R. The marine fungal metabolite, AD0157, inhibits angiogenesis by targeting the Akt signaling pathway. Mar. Drugs 2014, 12, 279–299. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Vilas, J.A.; Martínez-Poveda, B.; Quesada, A.R.; Medina, M.Á. Aeroplysinin-1, a Sponge-Derived Multi-Targeted Bioactive Marine Drug. Mar. Drugs 2016, 14, 1. https://doi.org/10.3390/md14010001
García-Vilas JA, Martínez-Poveda B, Quesada AR, Medina MÁ. Aeroplysinin-1, a Sponge-Derived Multi-Targeted Bioactive Marine Drug. Marine Drugs. 2016; 14(1):1. https://doi.org/10.3390/md14010001
Chicago/Turabian StyleGarcía-Vilas, Javier A., Beatriz Martínez-Poveda, Ana R. Quesada, and Miguel Ángel Medina. 2016. "Aeroplysinin-1, a Sponge-Derived Multi-Targeted Bioactive Marine Drug" Marine Drugs 14, no. 1: 1. https://doi.org/10.3390/md14010001
APA StyleGarcía-Vilas, J. A., Martínez-Poveda, B., Quesada, A. R., & Medina, M. Á. (2016). Aeroplysinin-1, a Sponge-Derived Multi-Targeted Bioactive Marine Drug. Marine Drugs, 14(1), 1. https://doi.org/10.3390/md14010001