
The Oligo Fucoidan Inhibits Platelet-Derived Growth Factor-Stimulated Proliferation of Airway Smooth Muscle Cells


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
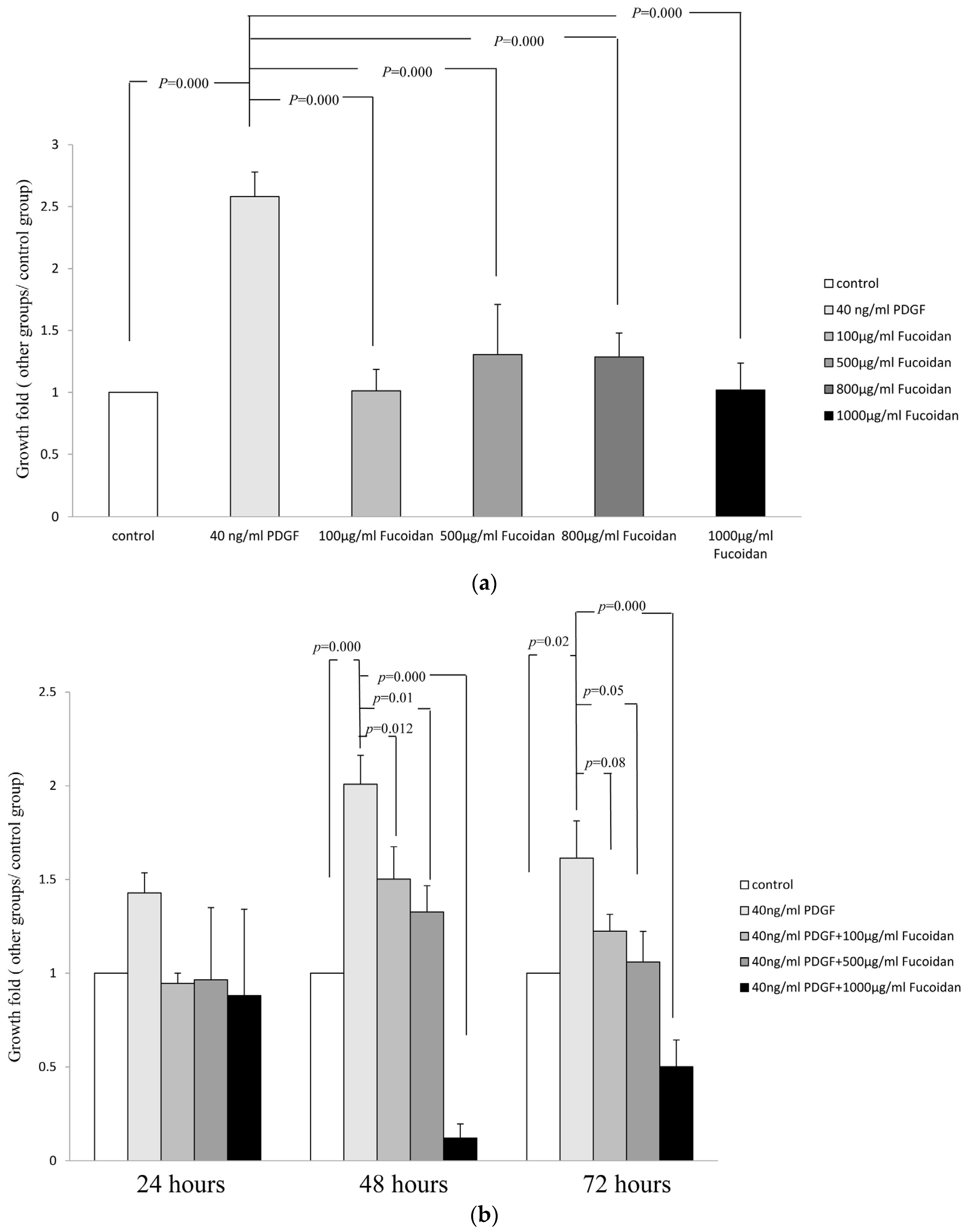
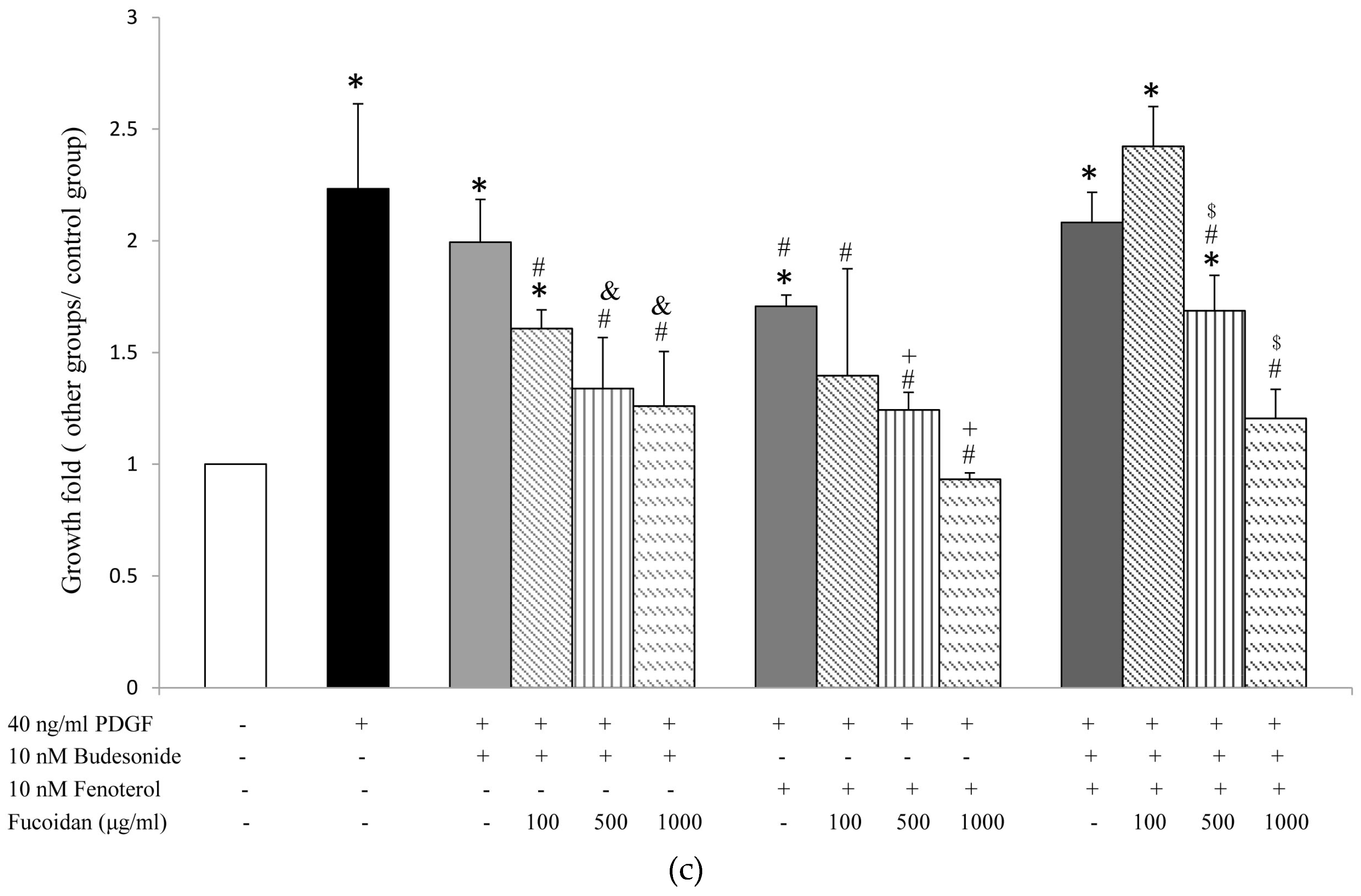
2.1. Oligo-Fucoidan Inhibited PDGF-Stimulated Proliferation of ASMCs


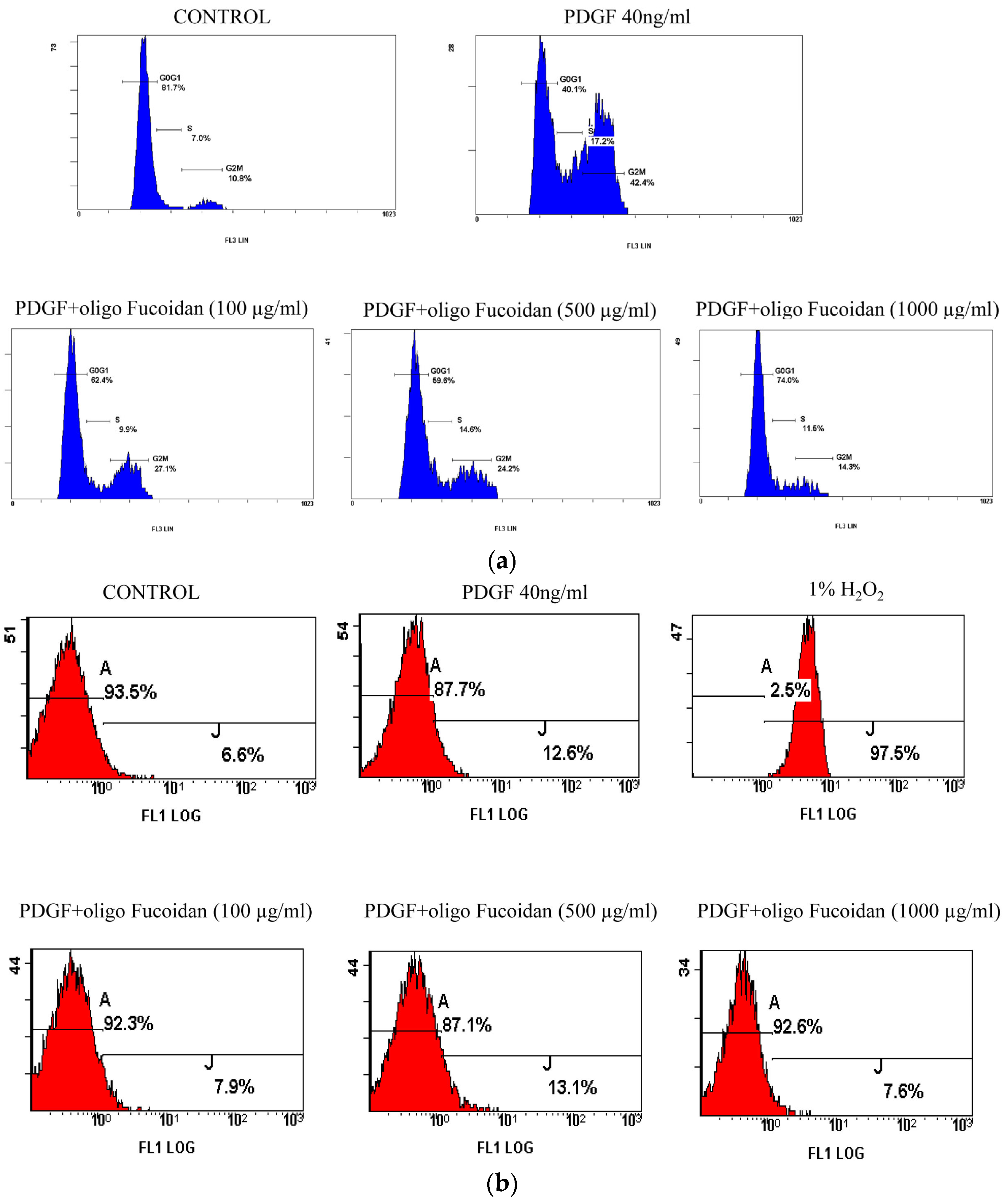
2.2. Oligo-Fucoidan Inhibited Cell Cycle Progression, but Did Not Induce Apoptosis in PGDF-Stimulated ASMCs

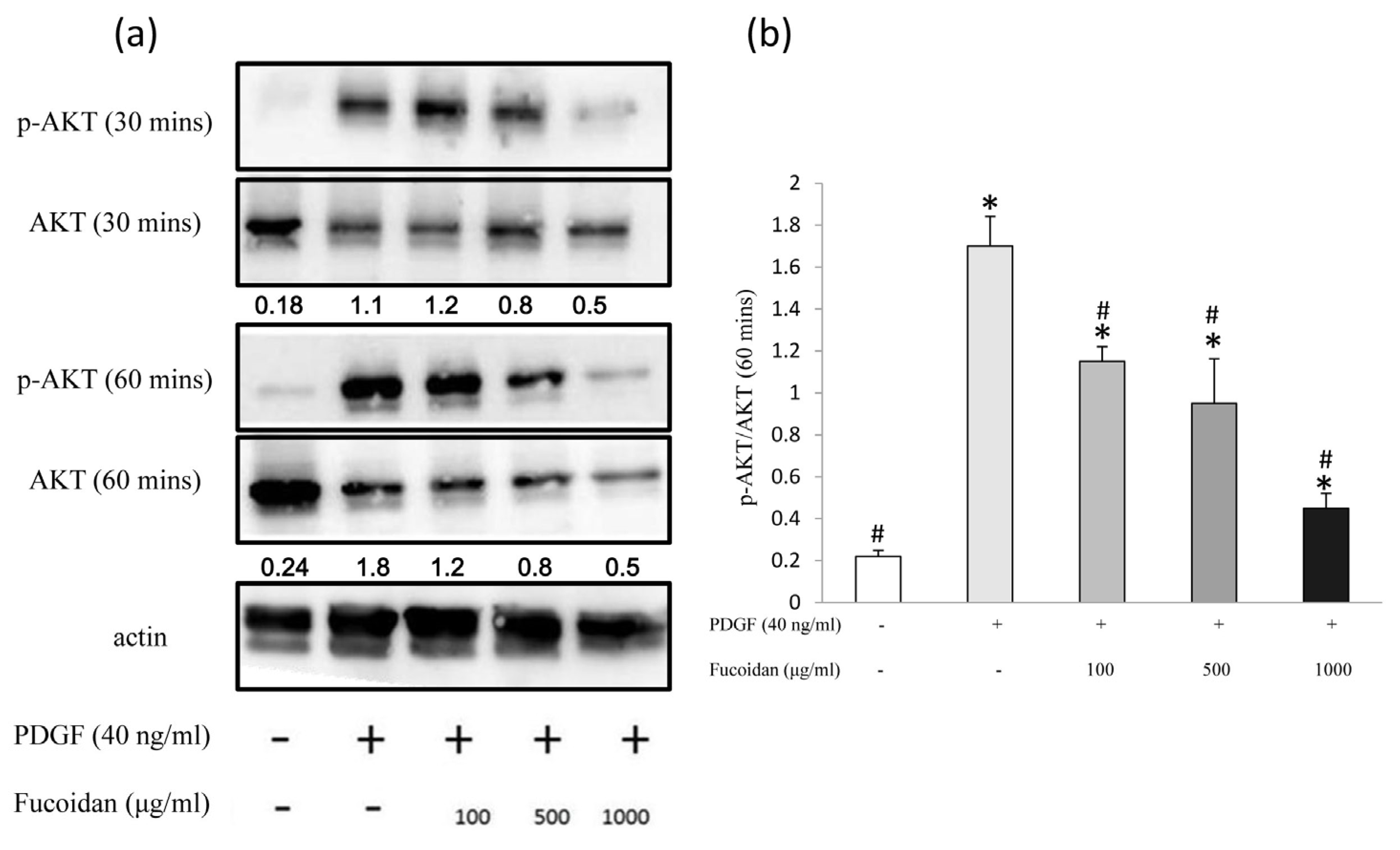
2.3. Oligo-Fucoidan Effectively Blocked the PDGF-Induced Akt and ERK1/2 Phosphorylation


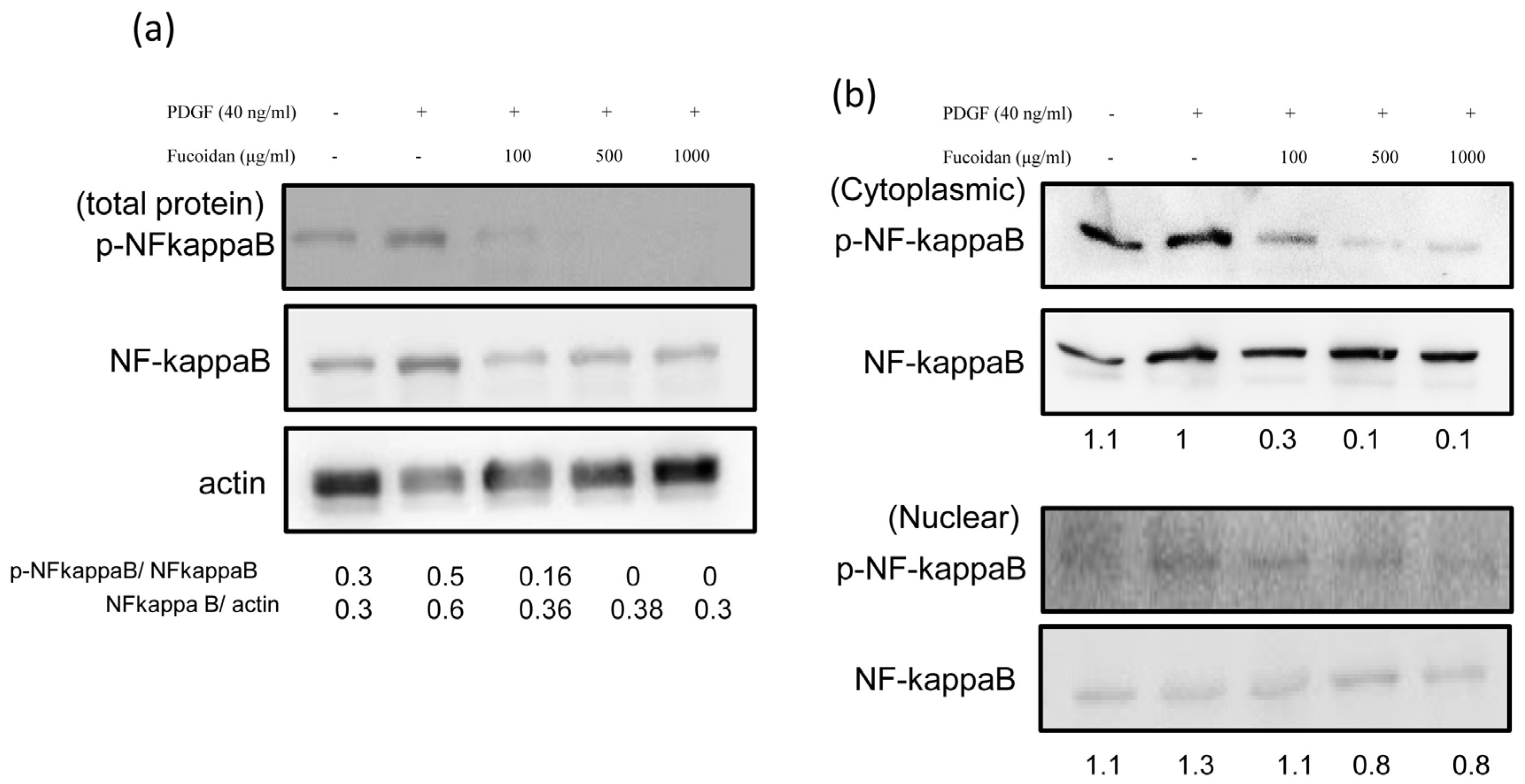
2.4. Oligo-Fucoidan Effectively Blocked the PDGF-Induced NF-κB Phosphorylation

3. Discussion
4. Materials and Methods
4.1. Culture of ASMCs
4.2. Evaluation of Cell Proliferation and Cell Cycle Progression
4.3. Apoptosis Assay
4.4. Extraction of Protein and Western Blot Analysis
4.5. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| The following abbreviations are used in this manuscript: |
| ASMCs: | Airway smooth muscle cells |
| AHR: | airway hyper-responsiveness |
| CAMs: | cell adhesion molecules |
| DMEM: | Dulbecco’s modified Eagle’s medium |
| ERK1/2: | extracellular signal-regulated kinase |
| FBS: | fetal bovine serum |
| GM-CSF: | granulocyte monocyte-colony stimulating factor |
| HBSS: | Hanks’ balanced salt solution |
| IL: | interleukin |
| ICAM: | intercellular CAM |
| ICS: | inhaled corticosteroids |
| IgE: | immunoglobulin E |
| LABA: | long-acting beta2-agonists |
| LIF: | leukemia inhibitory factor |
| MCP: | monocyte chemoattractant protein |
| MAPK: | mitogen activated protein kinase |
| (NF)-κB: | nuclear factor |
| PDGF: | Platelet-derived growth factor |
| PI3K: | phosphoinositide-3-kinase |
| TNF: | tumor necrosis factor |
| VCAM: | vascular CAM |
References
- Tzeng, Y.F.; Gau, B.S. Nursing care of a school-age child with asthma: An ecological system theory approach. Hu Li Za Zhi 2012, 59, 96–103. [Google Scholar] [PubMed]
- Kartha, S.; Naureckas, E.T.; Li, J.; Hershenson, M.B. Partial characterization of a novel mitogen-activated protein kinase/extracellular signal-regulated kinase activator in airway smooth-muscle cells. Am. J. Respir. Cell Mol. Biol. 1999, 20, 1041–1048. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.F.; Caramori, G.; Adcock, I.M. Inhaled corticosteroids as combination therapy with beta-adrenergic agonists in airways disease: Present and future. Eur. J. Clin. Pharmacol. 2009, 65, 853–871. [Google Scholar] [CrossRef] [PubMed]
- Stewart, A.G.; Grigoriadis, G.; Harris, T. Mitogenic actions of endothelin-1 and epidermal growth factor in cultured airway smooth muscle. Clin. Exp. Pharmacol. Physiol. 1994, 21, 277–285. [Google Scholar] [PubMed]
- Kelleher, M.D.; Abe, M.K.; Chao, T.S.; Jain, M.; Green, J.M.; Solway, J.; Rosner, M.R.; Hershenson, M.B. Role of map kinase activation in bovine tracheal smooth muscle mitogenesis. Am. J. Physiol. 1995, 268, L894–L901. [Google Scholar] [PubMed]
- Hirst, S.J.; Barnes, P.J.; Twort, C.H. PDGF isoform-induced proliferation and receptor expression in human cultured airway smooth muscle cells. Am. J. Physiol. 1996, 270, L415–L428. [Google Scholar] [PubMed]
- Goncharova, E.A.; Lim, P.N.; Chisolm, A.; Fogle, H.W., 3rd; Taylor, J.H.; Goncharov, D.A.; Eszterhas, A.; Panettieri, R.A., Jr.; Krymskaya, V.P. Interferons modulate mitogen-induced protein synthesis in airway smooth muscle. Am. J. Physiol. Lung Cell. Mol. Physiol. 2010, 299, L25–L35. [Google Scholar] [CrossRef] [PubMed]
- Yahiaoui, L.; Villeneuve, A.; Valderrama-Carvajal, H.; Burke, F.; Fixman, E.D. Endothelin-1 regulates proliferative responses, both alone and synergistically with PDGF, in rat tracheal smooth muscle cells. Cell. Physiol. Biochem. 2006, 17, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Bousquet, J.; Jeffery, P.K.; Busse, W.W.; Johnson, M.; Vignola, A.M. Asthma: From bronchoconstriction to airways inflammation and remodeling. Am. J. Respir. Crit. Care Med. 2000, 161, 1720–1745. [Google Scholar] [CrossRef] [PubMed]
- Fredriksson, L.; Li, H.; Eriksson, U. The PDGF family: Four gene products form five dimeric isoforms. Cytokine Growth Factor Rev. 2004, 15, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Halayko, A.J.; Tran, T.; Gosens, R. Phenotype and functional plasticity of airway smooth muscle: Role of caveolae and caveolins. Proc. Am. Thorac. Soc. 2008, 5, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Hirota, J.A.; Ask, K.; Farkas, L.; Smith, J.A.; Ellis, R.; Rodriguez-Lecompte, J.C.; Kolb, M.; Inman, M.D. In vivo role of platelet-derived growth factor-BB in airway smooth muscle proliferation in mouse lung. Am. J. Respir. Cell Mol. Biol. 2011, 45, 566–572. [Google Scholar] [CrossRef] [PubMed]
- Ito, I.; Fixman, E.D.; Asai, K.; Yoshida, M.; Gounni, A.S.; Martin, J.G.; Hamid, Q. Platelet-derived growth factor and transforming growth factor-beta modulate the expression of matrix metalloproteinases and migratory function of human airway smooth muscle cells. Clin. Exp. Allergy 2009, 39, 1370–1380. [Google Scholar] [CrossRef] [PubMed]
- Hirst, S.J.; Martin, J.G.; Bonacci, J.V.; Chan, V.; Fixman, E.D.; Hamid, Q.A.; Herszberg, B.; Lavoie, J.P.; McVicker, C.G.; Moir, L.M.; et al. Proliferative aspects of airway smooth muscle. J. Allergy Clin. Immunol. 2004, 114, S2–S17. [Google Scholar] [CrossRef] [PubMed]
- Krymskaya, V.P.; Penn, R.B.; Orsini, M.J.; Scott, P.H.; Plevin, R.J.; Walker, T.R.; Eszterhas, A.J.; Amrani, Y.; Chilvers, E.R.; Panettieri, R.A., Jr. Phosphatidylinositol 3-kinase mediates mitogen-induced human airway smooth muscle cell proliferation. Am. J. Physiol. 1999, 277, L65–L78. [Google Scholar] [PubMed]
- Zhou, L.; Hershenson, M.B. Mitogenic signaling pathways in airway smooth muscle. Respir. Physiol. Neurobiol. 2003, 137, 295–308. [Google Scholar] [CrossRef]
- Kusaykin, M.; Bakunina, I.; Sova, V.; Ermakova, S.; Kuznetsova, T.; Besednova, N.; Zaporozhets, T.; Zvyagintseva, T. Structure, biological activity, and enzymatic transformation of fucoidans from the brown seaweeds. Biotechnol. J. 2008, 3, 904–915. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Lu, F.; Wei, X.; Zhao, R. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, Q.; Zhang, Z.; Li, Z. Antioxidant activity of sulfated polysaccharide fractions extracted from laminaria japonica. Int. J. Biol. Macromol. 2008, 42, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Yanase, Y.; Hiragun, T.; Uchida, K.; Ishii, K.; Oomizu, S.; Suzuki, H.; Mihara, S.; Iwamoto, K.; Matsuo, H.; Onishi, N.; et al. Peritoneal injection of fucoidan suppresses the increase of plasma IgE induced by OVA-sensitization. Biochem. Biophys. Res. Commun. 2009, 387, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, H.; Tamauchi, H.; Hashimoto, M.; Nakano, T. Suppression of Th2 immune responses by mekabu fucoidan from undaria pinnatifida sporophylls. Int. Arch. Allergy Immunol. 2005, 137, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Chiou, Y.L.; Shieh, J.J.; Lin, C.Y. Blocking of Akt/NF-κB signaling by pentoxifylline inhibits platelet-derived growth factor-stimulated proliferation in brown norway rat airway smooth muscle cells. Pediatr. Res. 2006, 60, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Son, D.J.; Kang, J.; Kim, T.J.; Song, H.S.; Sung, K.J.; Yun-do, Y.; Hong, J.T. Melittin, a major bioactive component of bee venom toxin, inhibits PDGF receptor beta-tyrosine phosphorylation and downstream intracellular signal transduction in rat aortic vascular smooth muscle cells. J. Toxicol. Environ. Health A 2007, 70, 1350–1355. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.L.; Ling, D.Y.; Zhu, Q.Y.; Fan, W.J.; Zhang, W. The effect of 2,3,4′,5-tetrahydroxystilbene-2-O-beta-d glucoside on neointima formation in a rat artery balloon injury model and its possible mechanisms. Eur. J. Pharmacol. 2013, 698, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, S.E. Asthma: Defining of the persistent adult phenotypes. Lancet 2006, 368, 804–813. [Google Scholar] [CrossRef]
- Howarth, P.H.; Knox, A.J.; Amrani, Y.; Tliba, O.; Panettieri, R.A., Jr.; Johnson, M. Synthetic responses in airway smooth muscle. J. Allergy Clin. Immunol. 2004, 114, S32–S50. [Google Scholar] [CrossRef] [PubMed]
- Hirst, S.J. Airway smooth muscle as a target in asthma. Clin. Exp. Allergy 2000, 30 (Suppl. 1), 54–59. [Google Scholar] [CrossRef] [PubMed]
- Hirst, S.J. Airway smooth muscle cell culture: Application to studies of airway wall remodeling and phenotype plasticity in asthma. Eur. Respir. J. 1996, 9, 808–820. [Google Scholar] [CrossRef] [PubMed]
- Dekkers, B.G.; Pehlic, A.; Mariani, R.; Bos, I.S.; Meurs, H.; Zaagsma, J. Glucocorticosteroids and beta2-adrenoceptor agonists synergize to inhibit airway smooth muscle remodeling. J. Pharmacol. Exp. Ther. 2012, 342, 780–787. [Google Scholar] [CrossRef] [PubMed]
- Vo, T.S.; Kim, S.K. Marine-derived polysaccharides for regulation of allergic responses. Adv. Food Nutr. Res. 2014, 73, 1–13. [Google Scholar] [PubMed]
- Kanabar, V.; Hirst, S.J.; O’Connor, B.J.; Page, C.P. Some structural determinants of the antiproliferative effect of heparin-like molecules on human airway smooth muscle. Br. J. Pharmacol. 2005, 146, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Kilfeather, S.A.; Tagoe, S.; Perez, A.C.; Okona-Mensa, K.; Matin, R.; Page, C.P. Inhibition of serum-induced proliferation of bovine tracheal smooth muscle cells in culture by heparin and related glycosaminoglycans. Br. J. Pharmacol. 1995, 114, 1442–1446. [Google Scholar] [CrossRef] [PubMed]
- Na, H.J.; Moon, P.D.; Ko, S.G.; Lee, H.J.; Jung, H.A.; Hong, S.H.; Seo, Y.; Oh, J.M.; Lee, B.H.; Choi, B.W.; et al. Sargassum hemiphyllum inhibits atopic allergic reaction via the regulation of inflammatory mediators. J. Pharmacol. Sci. 2005, 97, 219–226. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C.-H.; Tsao, C.-F.; Ko, W.-S.; Chiou, Y.-L. The Oligo Fucoidan Inhibits Platelet-Derived Growth Factor-Stimulated Proliferation of Airway Smooth Muscle Cells. Mar. Drugs 2016, 14, 15. https://doi.org/10.3390/md14010015
Yang C-H, Tsao C-F, Ko W-S, Chiou Y-L. The Oligo Fucoidan Inhibits Platelet-Derived Growth Factor-Stimulated Proliferation of Airway Smooth Muscle Cells. Marine Drugs. 2016; 14(1):15. https://doi.org/10.3390/md14010015
Chicago/Turabian StyleYang, Chao-Huei, Chiung-Fang Tsao, Wang-Sheng Ko, and Ya-Ling Chiou. 2016. "The Oligo Fucoidan Inhibits Platelet-Derived Growth Factor-Stimulated Proliferation of Airway Smooth Muscle Cells" Marine Drugs 14, no. 1: 15. https://doi.org/10.3390/md14010015
APA StyleYang, C. -H., Tsao, C. -F., Ko, W. -S., & Chiou, Y. -L. (2016). The Oligo Fucoidan Inhibits Platelet-Derived Growth Factor-Stimulated Proliferation of Airway Smooth Muscle Cells. Marine Drugs, 14(1), 15. https://doi.org/10.3390/md14010015