
Different Culture Metabolites of the Red Sea Fungus Fusarium equiseti Optimize the Inhibition of Hepatitis C Virus NS3/4A Protease (HCV PR)


Abstract
:1. Introduction
2. Results and Discussion
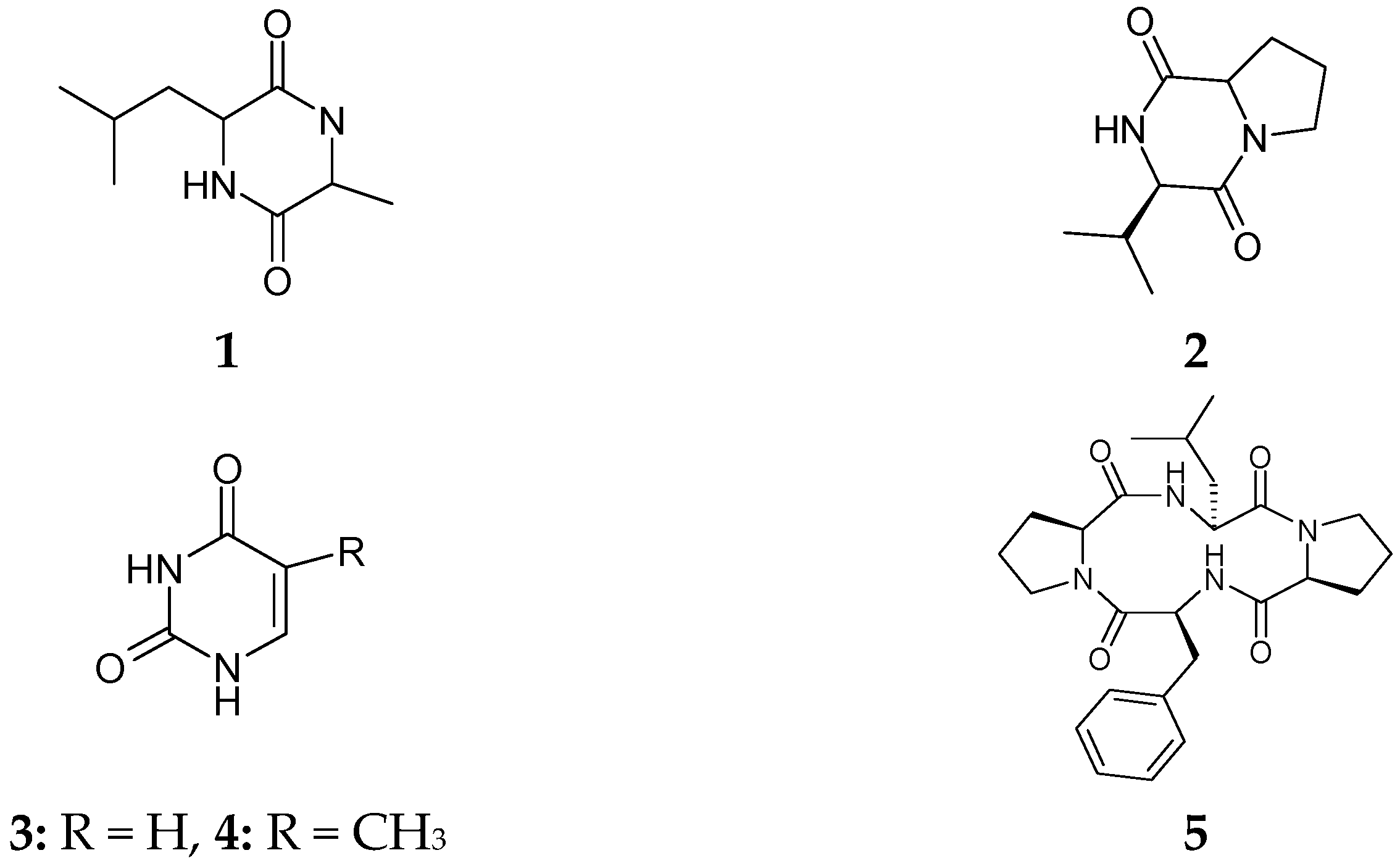
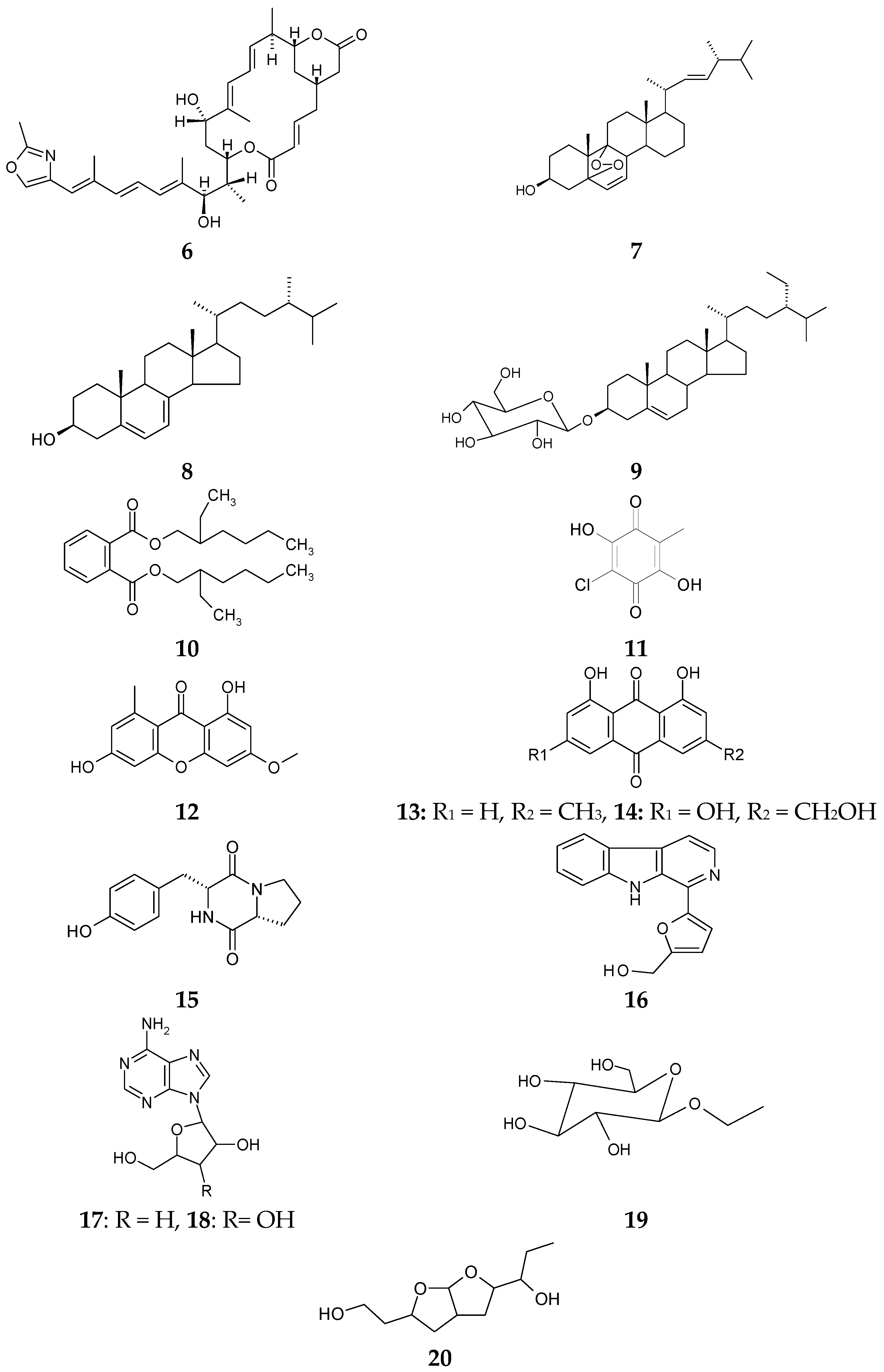
2.1. Characterization of Isolated Compounds
2.2. Antimicrobial Activity
2.3. HCV NS3-4A Protease and Trypsin Inhibition Activities
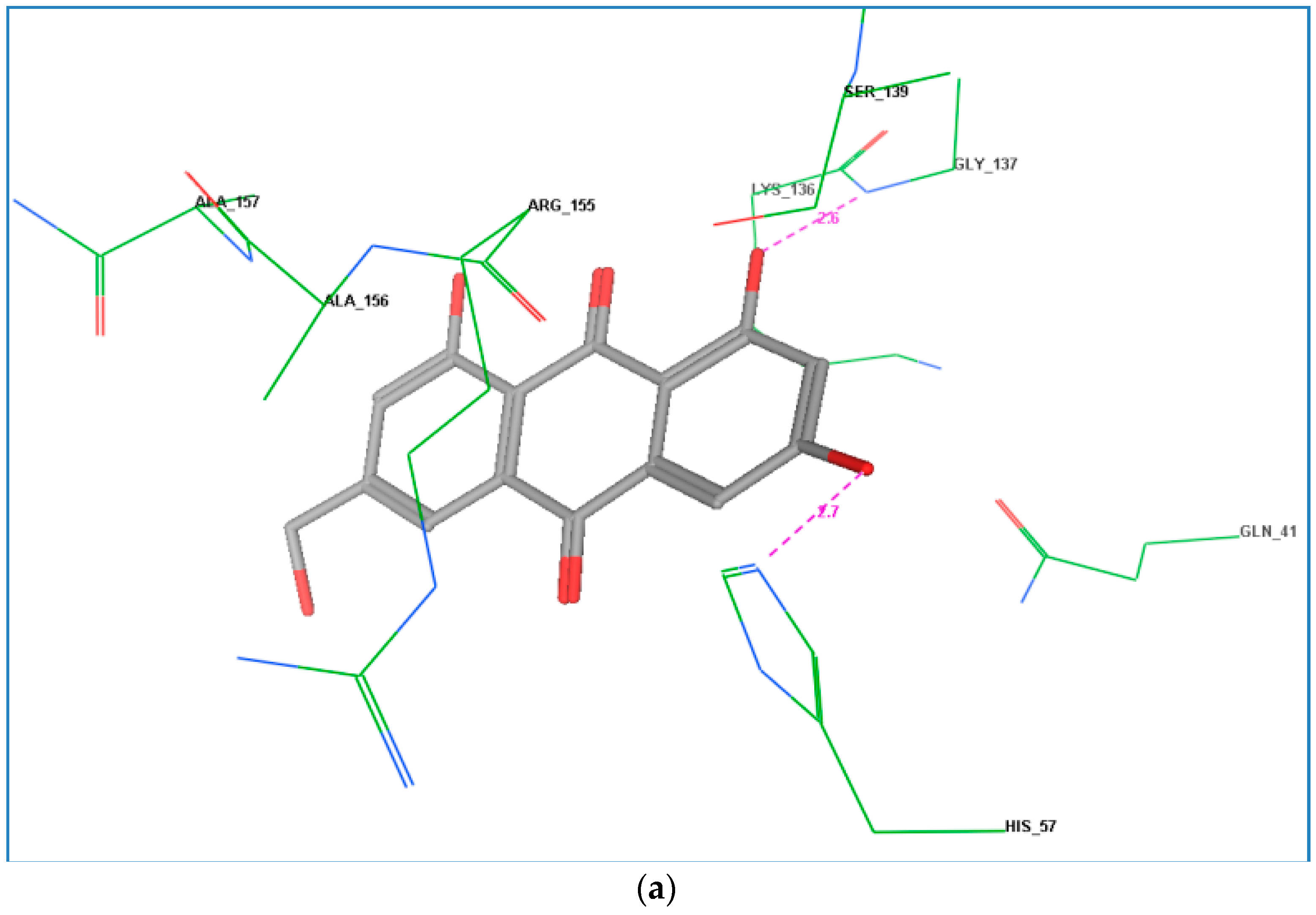
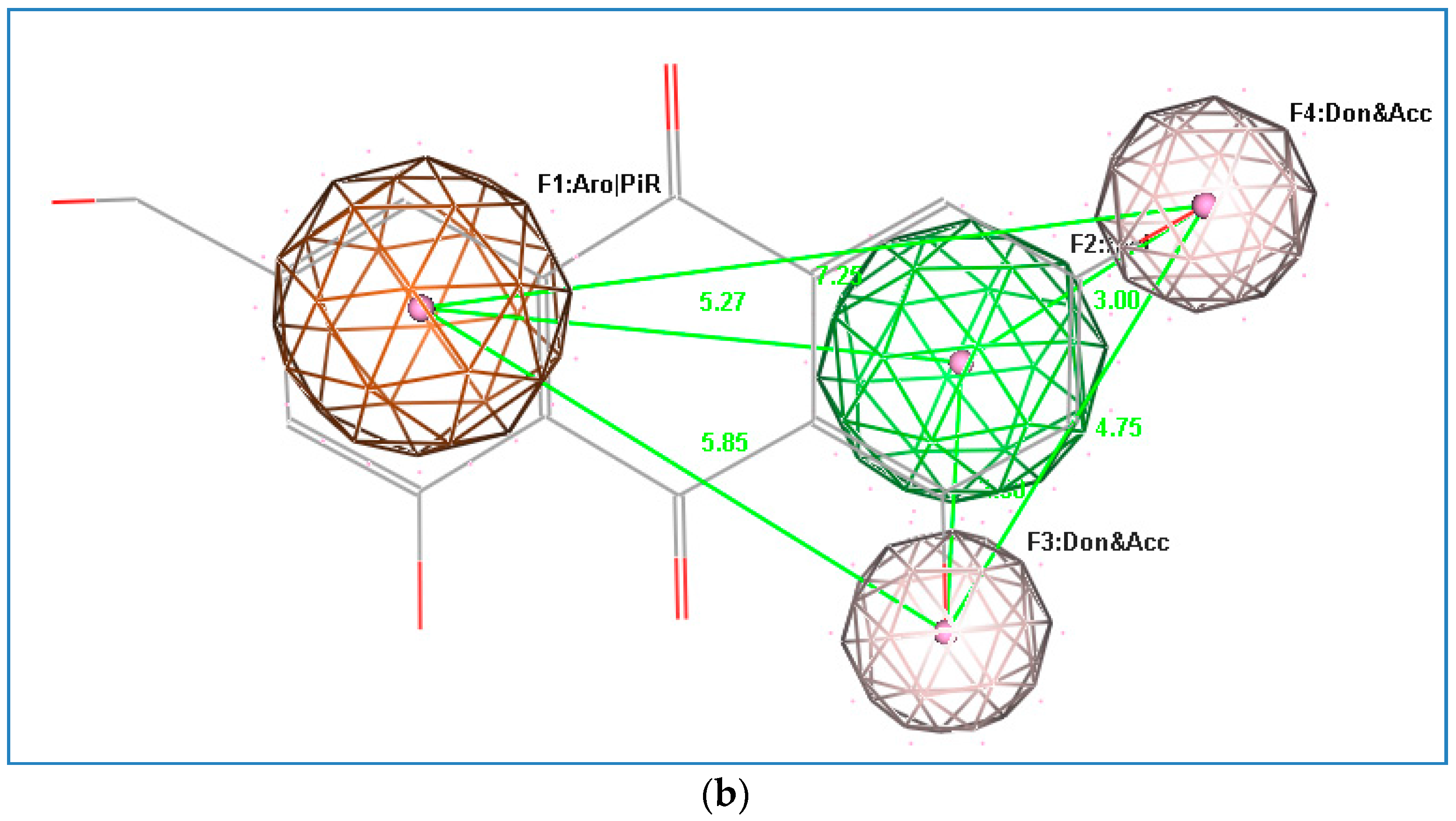
2.4. Docking Study
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Chemicals and Enzymes
3.3. Fungal Isolation and Culture Conditions
3.4. Identification of the Endophytic Isolates
3.5. Extraction and Isolation of Metabolites
3.6. Assay for Determination of HCV Protease Inhibitory Activity
3.7. Green Protease Assay
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yu, Y.; Jing, J.F.; Tong, X.K.; Pei, H.I.; Yuan, L.C.; You, H.H.; Wei, T.; Jian, Z.P. Discovering novel anti-HCV compounds with inhibitory activities toward HCV NS3/4A protease. Acta Pharmacol. Sin. 2014, 35, 1074–1081. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.L.; Morgenstern, K.A.; Lin, C.; Fox, T.; Dwyer, M.D.; Landro, J.A.; Chambers, S.P.; Markland, W.; Lepre, C.A.; O’Malley, E.T. Crystal structure of the hepatitis C virus NS3 protease domain complexed with a synthetic NS4A cofactor peptide. Cell 1996, 87, 343–355. [Google Scholar] [CrossRef]
- Dai, J.R.; Carte, B.K.; Sidebottom, P.J.; Yew, A.L.S.; Ng, S.; Huang, Y.; Butler, M.S. Circumdatin G, a new alkaloid from the fungus Aspergillus ochraceus. J. Nat. Prod. 2001, 64, 125–126. [Google Scholar] [CrossRef] [PubMed]
- Au, J.S.; Pockros, P.J. Novel therapeutic approaches for hepatitis C. Clin. Pharmacol. Ther. 2014, 95, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Schulz, B.; Boyle, C. What are endophytes. In Microbial Root Endophytes; Schulz, B., Boyle, C., Sieber, T.N., Eds.; Springer: Berlin, Germany, 2006; pp. 1–13. [Google Scholar]
- Zhang, H.W.; Song, Y.C.; Tan, R.X. Biology and chemistry of endophytes. Nat. Prod. Rep. 2006, 23, 753–771. [Google Scholar] [CrossRef] [PubMed]
- García-Caballero, M.; Marí-Beffa, M.; Cañedo, L.; Medina, M.A.; Quesada, A.R. Toluquinol, a marine fungus metabolite, is a new angiosuppresor that interferes with the Akt pathway. Biochem. Pharmacol. 2013, 85, 1727–1740. [Google Scholar] [CrossRef] [PubMed]
- Hansen, F.T.; Gardiner, D.M.; Lysøe, E.; Fuertes, P.R.; Tudzynsk, B.; Wiemann, P.; Sondergaard, T.E.; Giese, H.; Brodersen, D.E.; Sørensen, J.L. An update to polyketide synthase and non-ribosomal synthetase genes and nomenclature in Fusarium. Fungal Genet. Biol. 2015, 75, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Stępień, L.; Gromadzka, K.; Chełkowsk, J. Polymorphism of mycotoxin biosynthetic genes among Fusarium equiseti isolates from Italy and Poland. J. Appl. Genet. 2012, 53, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Mirocha, C.J.; Wen, Y. Formyl Fusarochromanone and Diacetyl Fusarochromanone, Two New Metabolites of Fusarium equiseti. J. Nat. Prod. 1991, 54, 1165–1167. [Google Scholar] [CrossRef]
- Xie, W.; Mirocha, C.J.; Wen, Y. Isolation and Structure Identification of Two New Derivatives of the Mycotoxin Fusarochromenone Produced by Fusarium equiseti. J. Nat. Prod. 1995, 58, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Dréau, D.; Foster, M.; Hogg, M.; Culberson, C.; Nunes, P.; Wuthier, R.E. Inhibitory effects of fusarochromanone on melanoma growth. Anticancer Drugs 2007, 18, 897–904. [Google Scholar] [PubMed]
- Kuiper, G.G.; Carlsson, B.; Grandien, K.; Enmark, E.; Haggblad, J.; Nilsson, S.; Gustafsson, J.A. Comparison of the ligand binding specificity and transcript tissue distribution of estrogen receptors alpha and beta. Endocrinology 1997, 138, 863–870. [Google Scholar] [PubMed]
- Gromadzka, K.; Waśkiewicz, A.; Chełkowski, J.; Goliński, P. Zearalenone and its metabolites: Occurrence, detection, toxicity and guidelines. World Mycotoxin J. 2008, 1, 209–220. [Google Scholar] [CrossRef]
- Caesar, F.; Jansson, K.K.; Mutschler, E.E. Nigragillin, a new alkaloid from the Aspergillus niger group. 1. Isolation and structure clarification of nigragillin and a dioxopiperazine. Pharm. Acta Helv. 1969, 44, 676–690. [Google Scholar] [PubMed]
- Shigemori, H.; Tenma, M.; Shimazaki, K.; Kobayashi, J. Three new metabolites from the marine yeast Aureobasidium pullulans. J. Nat. Prod. 1998, 61, 696–698. [Google Scholar] [CrossRef] [PubMed]
- Barrow, C.J.; Sun, H.H. Spiroquinazoline, a novel substance-p inhibitor with a new carbon skeleton, isolated from Aspergillus flavipes. J. Nat. Prod. 1994, 57, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Rungprom, W.; Siwu, E.R.O.; Lambert, L.K.; Dechsakulwatana, C.; Barden, M.C.; Kokpol, U.; Blanchfield, J.T.; Kita, M.; Garson, M.J. Cyclic tetrapeptides from marine bacteria associated with the seaweed Diginea sp. and the sponge Halisarca ectofibrosa. Tetrahedron 2008, 64, 3147–3152. [Google Scholar] [CrossRef]
- Shaaban, M.; Schröder, D.; Shaaban, K.A.; Helmke, E.; Grün-Wollny, I.; Wagner-Döbler, I.; Laatsch, H. Flazin, Perlolyrin, and other β-Carbolines from marine derived Bacteria. Rev. Latinoam. Quím. 2007, 35, 58–67. [Google Scholar]
- Kiyoto, S.; Kawai, Y.; Kawakita, T.; Kino, E.; Okuhara, M.; Uchida, I.; Tanaka, H.; Hashimoto, M.; Terano, H.; Kohsaka, M.; et al. A new antitumor complex, WF-1360, WF-1360A, B, C, D, E and F. J. Antibiot. 1986, 39, 762–772. [Google Scholar] [PubMed]
- Cimino, G.; de Rosa, S.; de Stefano, S.; Sodano, G. C-18 Hydroxy steroids from the Mediterranean gorgonian Leptogorgia sarmentosa. Cell. Mol. Life Sci. 1984, 40, 339–340. [Google Scholar] [CrossRef]
- Che, Y.; Gloer, J.B.; Scott, J.A.; Malloch, D. Communiols A–D: New mono- and bis-tetrahydrofuran derivatives from the coprophilous fungus Podospora communis. Tetrahedron Lett. 2004, 45, 6891–6894. [Google Scholar] [CrossRef]
- Cacho, R.A.; Chooi, Y.H.; Zhou, H.; Tang, Y. Complexity generation in fungal polyketide biosynthesis: A spirocyclic-forming P450 in the concise pathway to the antifungal drug griseofulvin. ACS Chem. Biol. 2013, 8, 2322–2330. [Google Scholar] [CrossRef] [PubMed]
- Hawas, U.W.; El-Halawany, A.M.; Ahmed, E.F. Hepatitis C virus NS3-NS4A protease inhibitors from the endophytic Penicillium chrysogenum isolated from the red alga Liagora viscida. Z. Naturforsch. C 2013, 68, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.M.M.U.; Hossain, M.S. Scopoletin and β-sitosterol glucoside from roots of Ipomoea digitata. J. Pharmacogn. Phytochem. 2015, 4, 5–7. [Google Scholar]
- Page, M.J.; di Cera, E. Serine peptidases: Classification, structure and function. Cell. Mol. Life Sci. 2008, 65, 1220–1236. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; di Cera, E. Evolution of peptidase diversity. J. Biol. Chem. 2008, 283, 30010–30014. [Google Scholar] [CrossRef] [PubMed]
- Njoroge, F.G.; Chen, K.X.; Shih, N.Y.; Piwinski, J.J. Challenges in modern drug discovery: A case study of boceprevir, an HCV protease inhibitor for the treatment of hepatitis C virus infection. Acc. Chem. Res. 2008, 41, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.X.; Njoroge, F.G.; Prongay, A.; Pichardo, J.; Madison, V.; Girijavallabhan, N. Synthesis and biological activity of macrocyclic inhibitors of hepatitis C virus (HCV) NS3 protease. Bioorg. Med. Chem. Lett. 2005, 15, 4475–4478. [Google Scholar] [CrossRef] [PubMed]
- Chomczynski, P. A reagent for the single-step simultaneous isolation of RNA, DNA and proteins from cell and tissue samples. Biotechniques 1993, 15, 532–534. [Google Scholar] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Gram-Positive Bacteria | Fungi | ||
|---|---|---|---|---|
| Staphylococcus aureus | Bacillus megaterium | Bacillus subtilis | Candida albicans | |
| Ethyl acetate extract (bi-malt culture) | 10 * | 11 | 18 | 13 |
| Cyclo-l-Ala-l-Leu (1) | - | 12 | - | 13 |
| Cyclo(l-Pro-l-Val) (2) | - | 11 | 18 | 9 |
| Cyclo-(Phenylalanyl-Pro-Leu-Pro) (5) | 9 | 10 | - | 12 |
| 17-Demethyl-2,11-dideoxy-rhizoxin (6) | 12 | 8 | - | 19 |
| 3-O-β-Glucosylsitosterol (9) | 9 | 14 | 11 | 10 |
| Griseoxanthone C (12) | - | 10 | 13 | - |
| Ethyl acetate extract (Czapek culture) | 15 | 17 | 9 | 19 |
| Chrysophanol (13) | 15 | 10 | - | 11 |
| ω-Hydroxyemodin (14) | 12 | 17 | 9 | 18 |
| Cyclo(l-Tyr-l-Pro) (15) | 11 | 8 | - | 13 |
| Perlolyrine (16) | 14 | 9 | 8 | 10 |
| Cordycepin (17) | 16 | 11 | 9 | 14 |
| Ara-A (18) | 10 | 12 | 12 | - |
| Oxytetracycline (30 μg) | 17 | 20 | 18 | - |
| Biomalt-Peptone Culture Medium | Czapek’s (Cz)-Peptone Culture Medium | ||||
|---|---|---|---|---|---|
| Sample | IC50 [μM] | ||||
| HCV Protease Inhibitory Activity | Trypsin Inhibitory Activity | Sample | HCV Protease Inhibitory Activity | Trypsin Inhibitory Activity | |
| EtOAc extract (Biomalt) | 56.0 ± 18 μg/mL | >100 | EtOAc extract (Cz) | 27.65 ± 2.2 μg/mL | 88.89 ± 3.39 μg/mL |
| Cyclo-l-Ala-l-Leu (1) | 58.33 ± 3.51 | Nt | Chrysophanol (13) | >1000 | Nt |
| Cyclo(l-Pro-l-Val) (2) | 23.29 ± 1.23 | 54.21 ± 1.88 | ω-Hydroxyemodin (14) | 10.71 ± 2.3 | 48.52 ± 1.9 |
| Thymine (4) | 51.82 ± 2.49 | Nt | Cyclo(l-Tyr-l-Pro) (15) | 18.20 ± 1.7 | 37.91 ± 3.8 |
| Cyclo-(Phenylalanyl-Pro-Leu-Pro) (5) | 29.45 ± 1.98 | 226.21 ± 4.26 | Perlolyrine (16) | 37.89 ± 2.11 | >1000 |
| 17-Demethyl-2,11-dideoxy-rhizoxin (6) | 34.42 ± 1.44 | 144.67 ± 3.66 | Cordycepin (17) | 22.35 ± 3.12 | >1000 |
| Ergostra-5,7-dien-3β-ol (8) | 77.14 ± 4.55 | Nt | Ara-A (18) | 24.53 ± 2.3 | 137.58 ± 2.49 |
| 3-O-β-Glucosylsitosterol (9) | 76.56 ± 3.78 | Nt | Communiol D (19) | >1000 | Nt |
| 5-Chloro-3,6-dihydroxy-2-methyl-1,4-benzoquinone (11) | 35.15 ± 3.92 | 294.82 ± 2.87 | Ethyl-O-β-glucoside (20) | >1000 | Nt |
| Griseoxanthone C (12) | 19.88 ± 1.35 | 51.33 ± 2.34 | |||
| HCV-I2 | 1.64 ± 0.5 | HCV-I2 | 1.64 ± 0.5 | ||
| T-I | 0.5 | T-I | 0.5 | ||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hawas, U.W.; Al-Farawati, R.; Abou El-Kassem, L.T.; Turki, A.J. Different Culture Metabolites of the Red Sea Fungus Fusarium equiseti Optimize the Inhibition of Hepatitis C Virus NS3/4A Protease (HCV PR). Mar. Drugs 2016, 14, 190. https://doi.org/10.3390/md14100190
Hawas UW, Al-Farawati R, Abou El-Kassem LT, Turki AJ. Different Culture Metabolites of the Red Sea Fungus Fusarium equiseti Optimize the Inhibition of Hepatitis C Virus NS3/4A Protease (HCV PR). Marine Drugs. 2016; 14(10):190. https://doi.org/10.3390/md14100190
Chicago/Turabian StyleHawas, Usama W., Radwan Al-Farawati, Lamia T. Abou El-Kassem, and Adnan J. Turki. 2016. "Different Culture Metabolites of the Red Sea Fungus Fusarium equiseti Optimize the Inhibition of Hepatitis C Virus NS3/4A Protease (HCV PR)" Marine Drugs 14, no. 10: 190. https://doi.org/10.3390/md14100190