
Astaxanthin and Docosahexaenoic Acid Reverse the Toxicity of the Maxi-K (BK) Channel Antagonist Mycotoxin Penitrem A




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Schwann Cells (RSC 96 ATCC CRL-2765) In Vitro Model
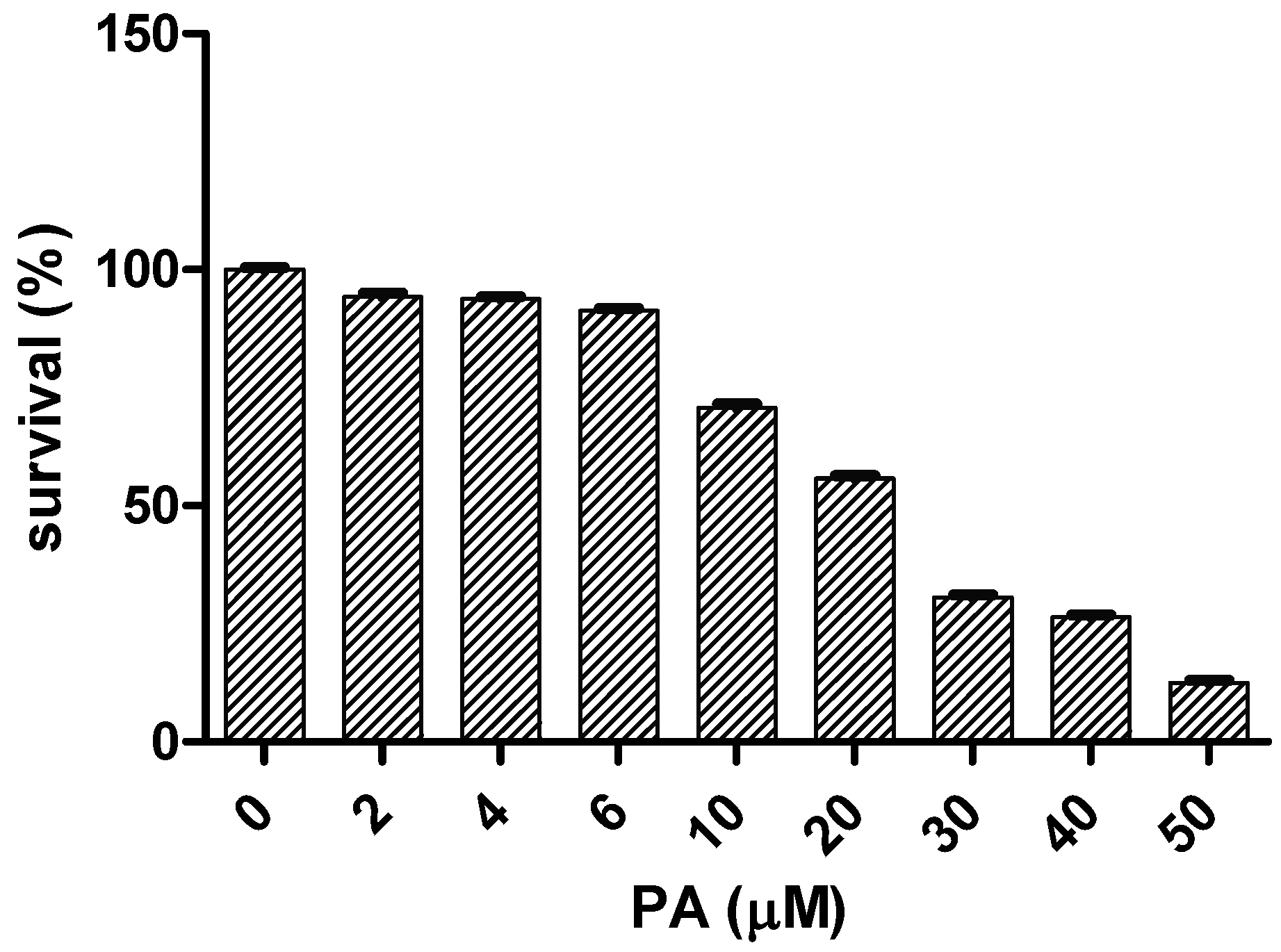
2.1.1. In Vitro Effects of PA on Neuronal Schwann Cell (RSC 96 ATCC CRL-2765) Proliferation and Survival
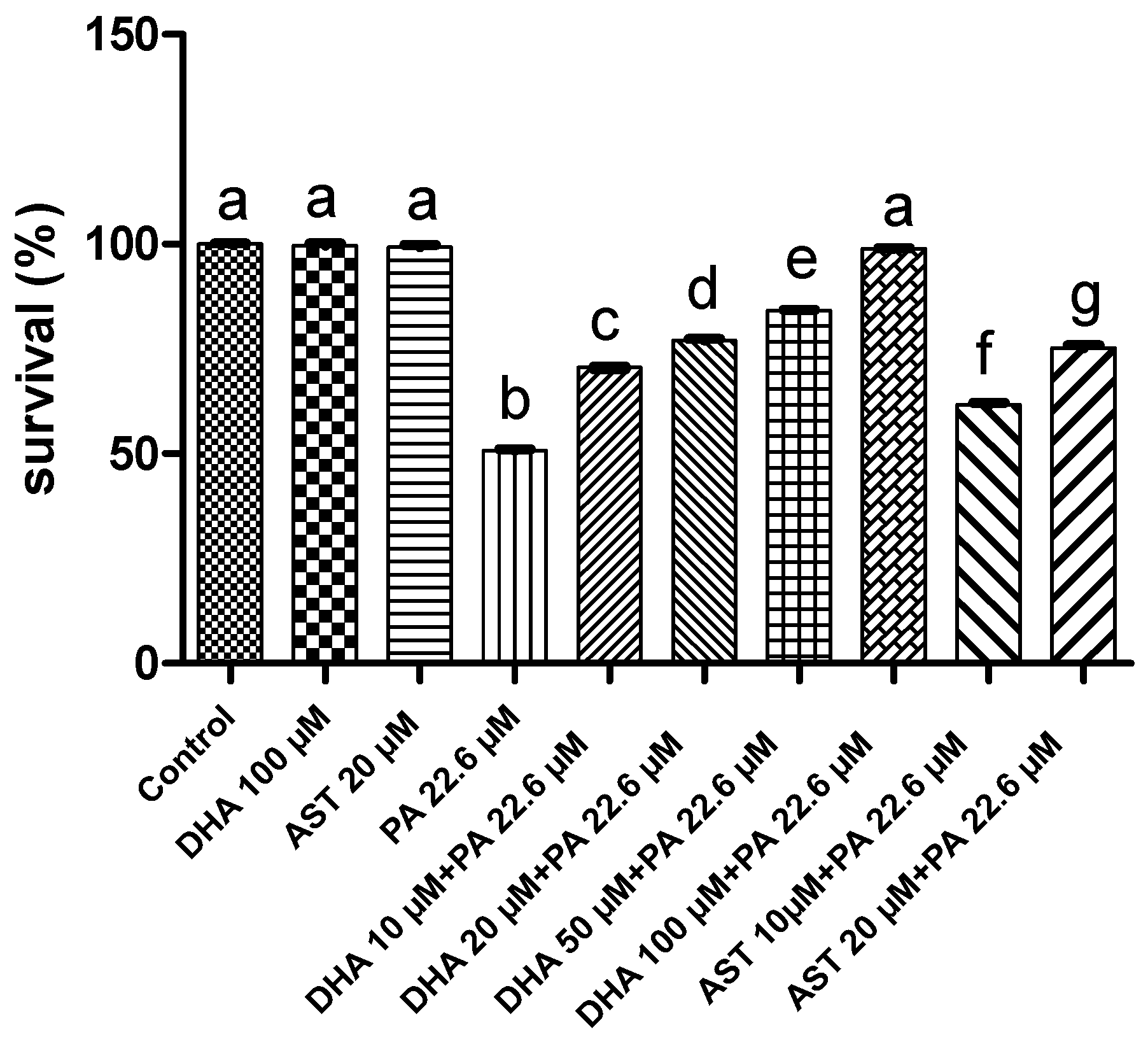
2.1.2. Protective Effects of AST and DHA against the PA-Induced Toxicity to Schwann Cells
2.2. Caenorhbditis elegans In Vivo Model
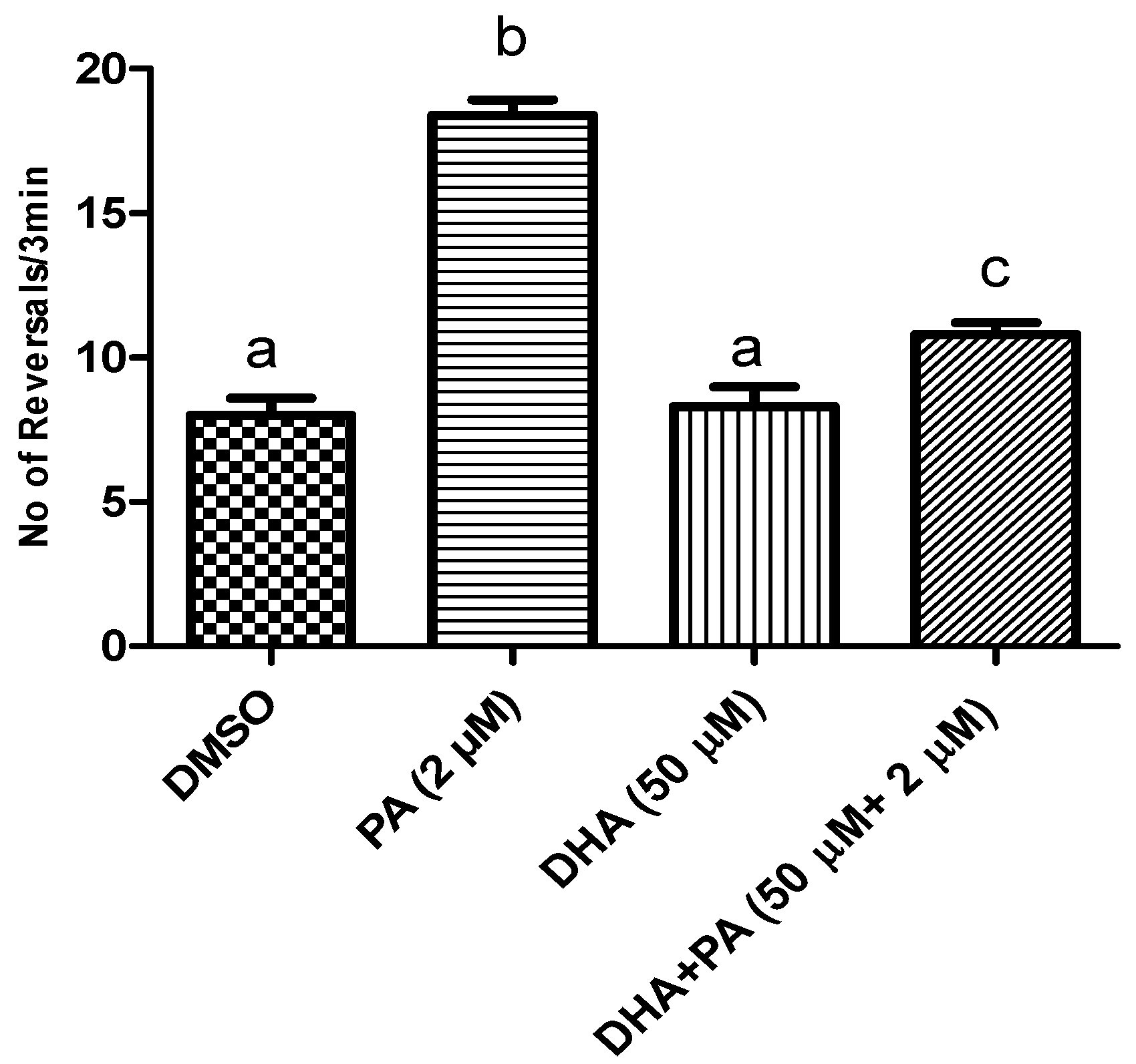
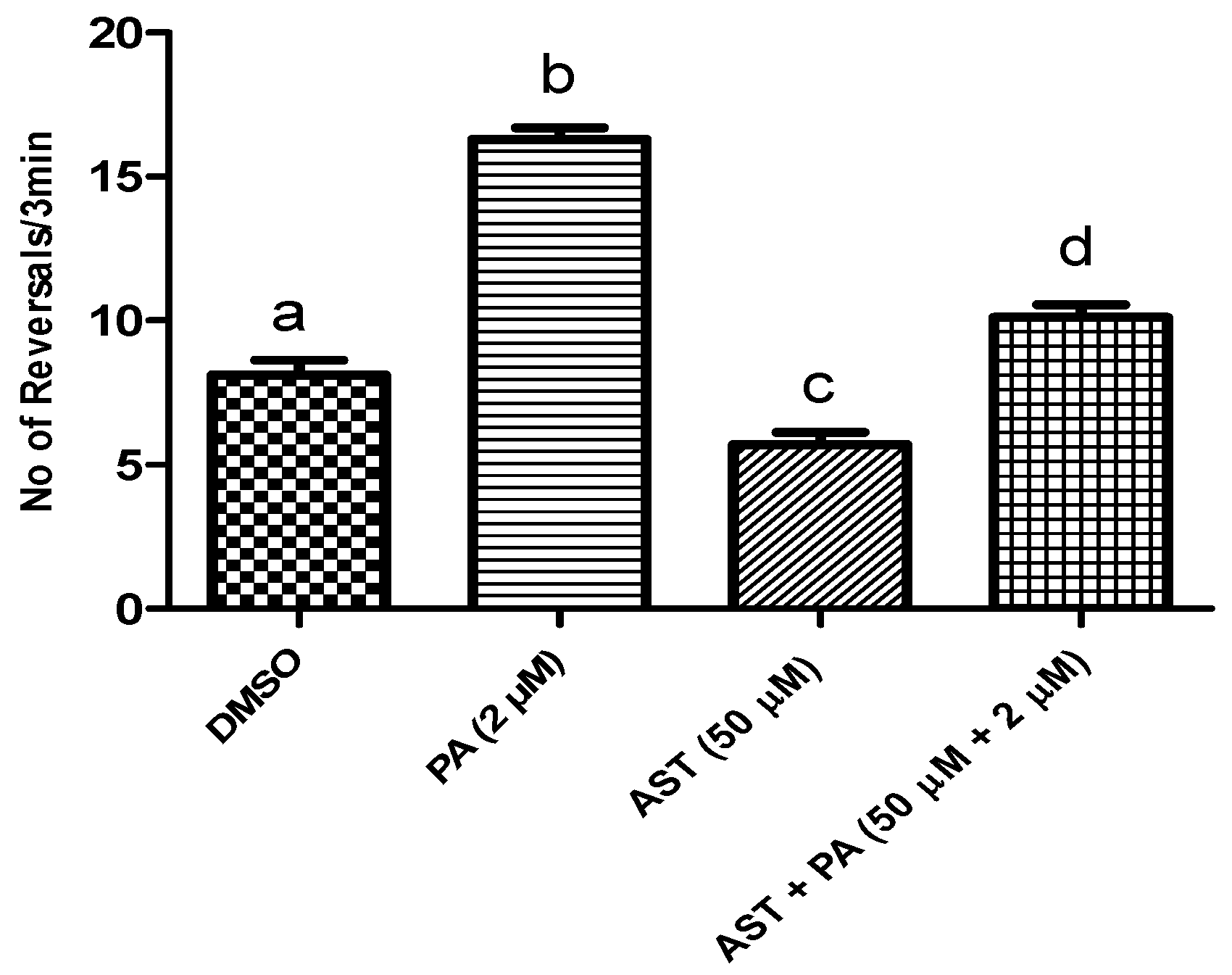
DHA and AST Counteracted PA’s BK Channel Antagonistic Activity in C. elegans Model
2.3. Sprague Dawley Rat In Vivo Model
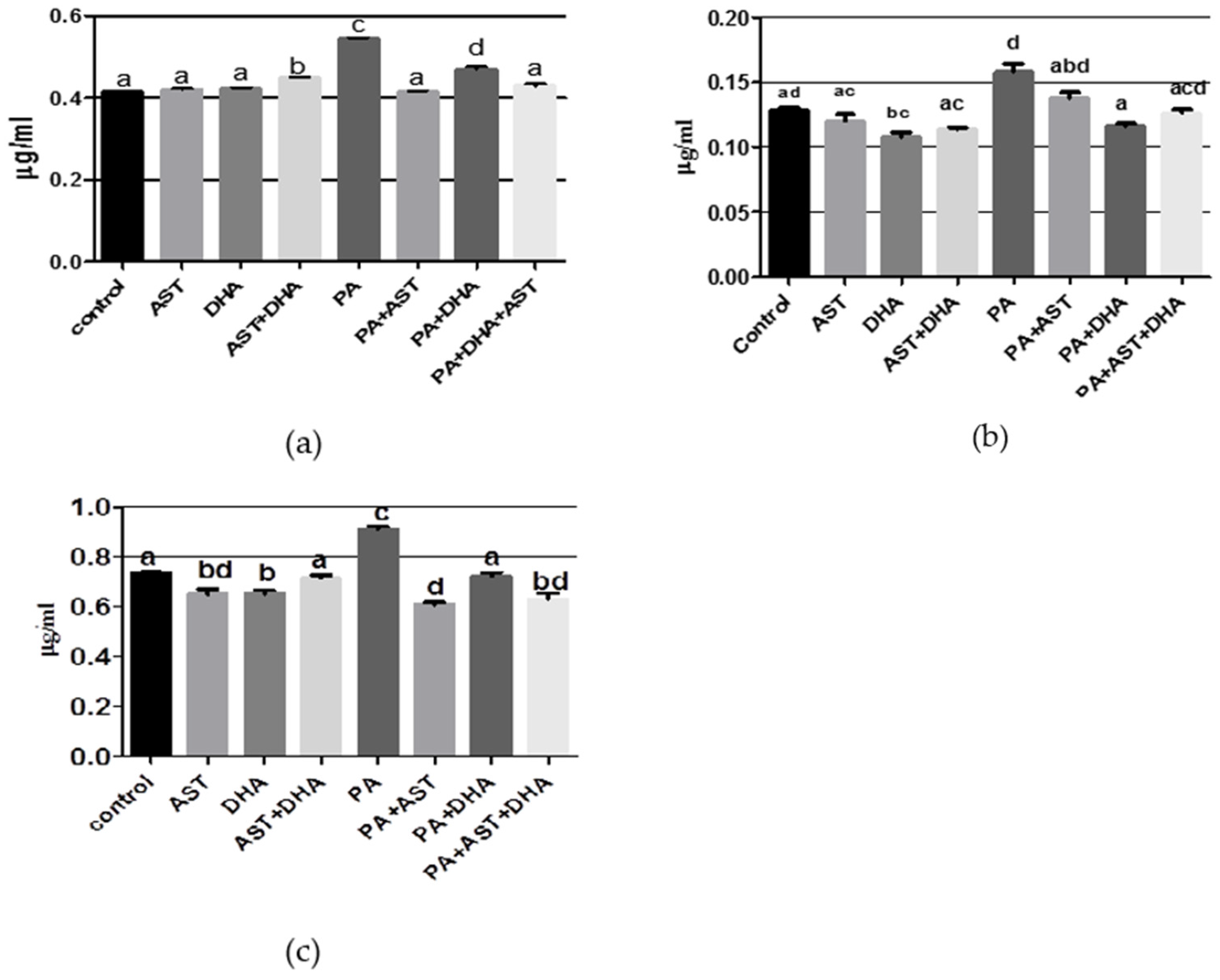
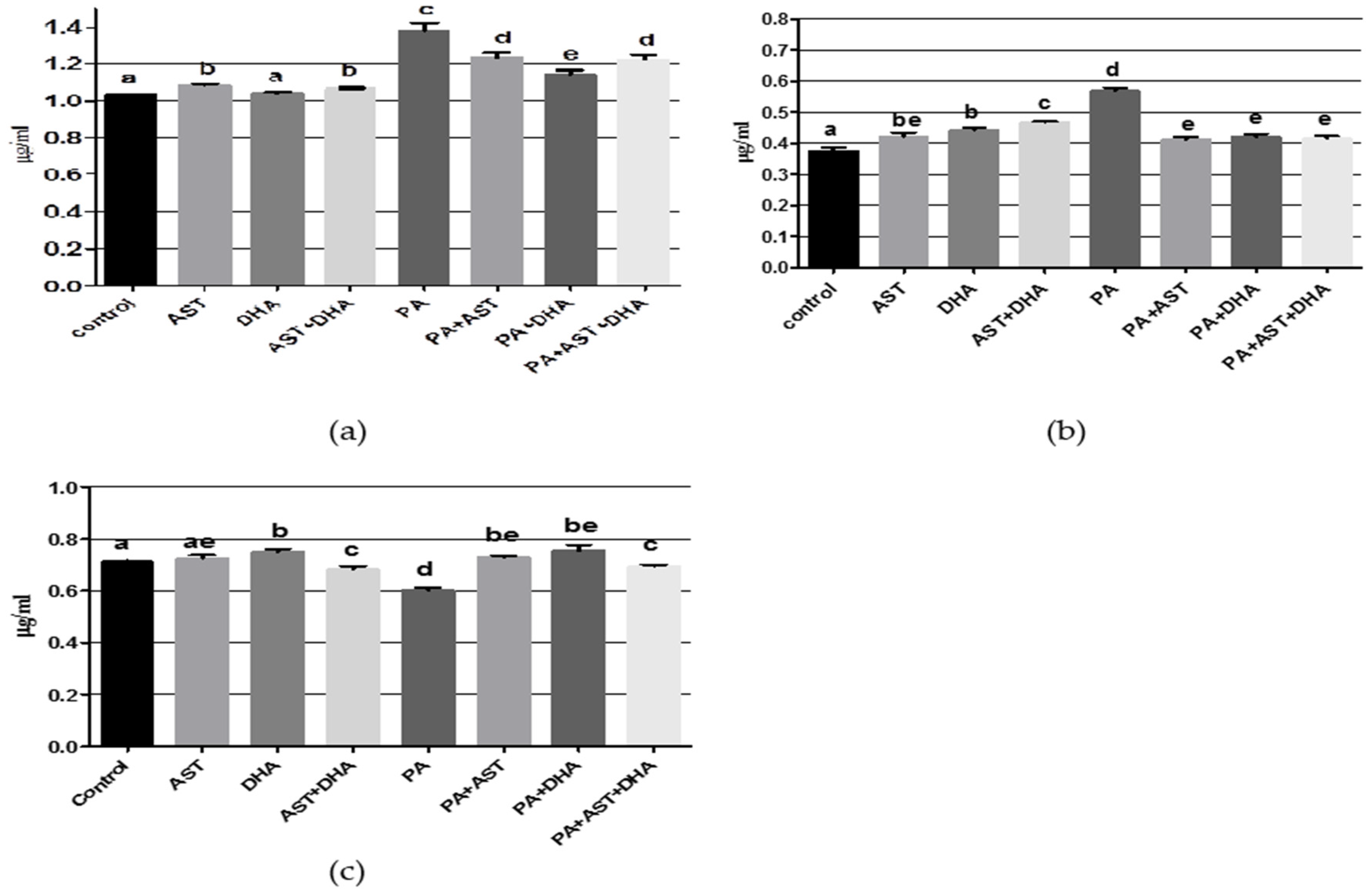
2.3.1. DHA and AST Counteract the Toxic Effects of PA on Amino Acid and Monoamine Neurotransmitters in Sprague Dawley Rats
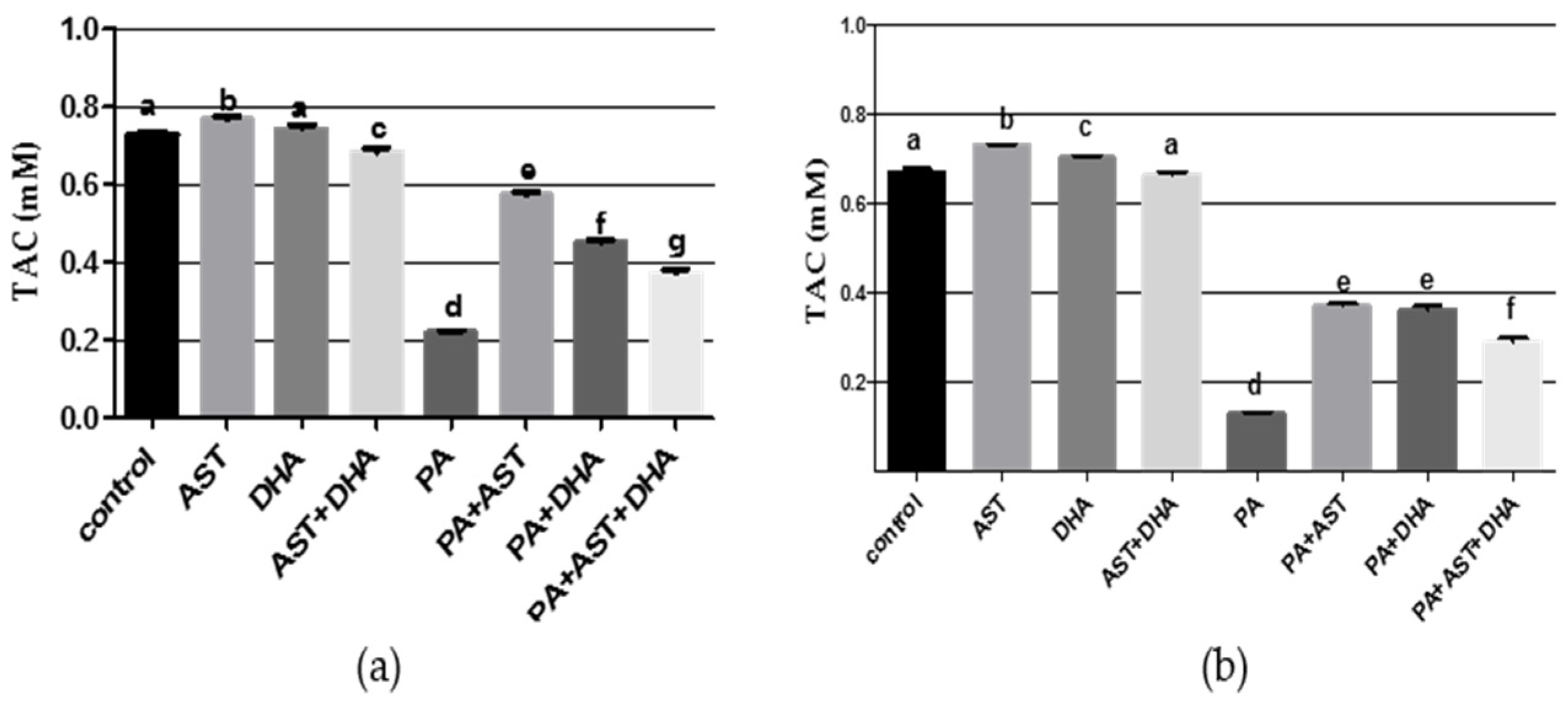
2.3.2. DHA and AST Significantly Increased TAC Value in PA-Treated Sprague Dawley Rat Serum and Tissue Homogenates
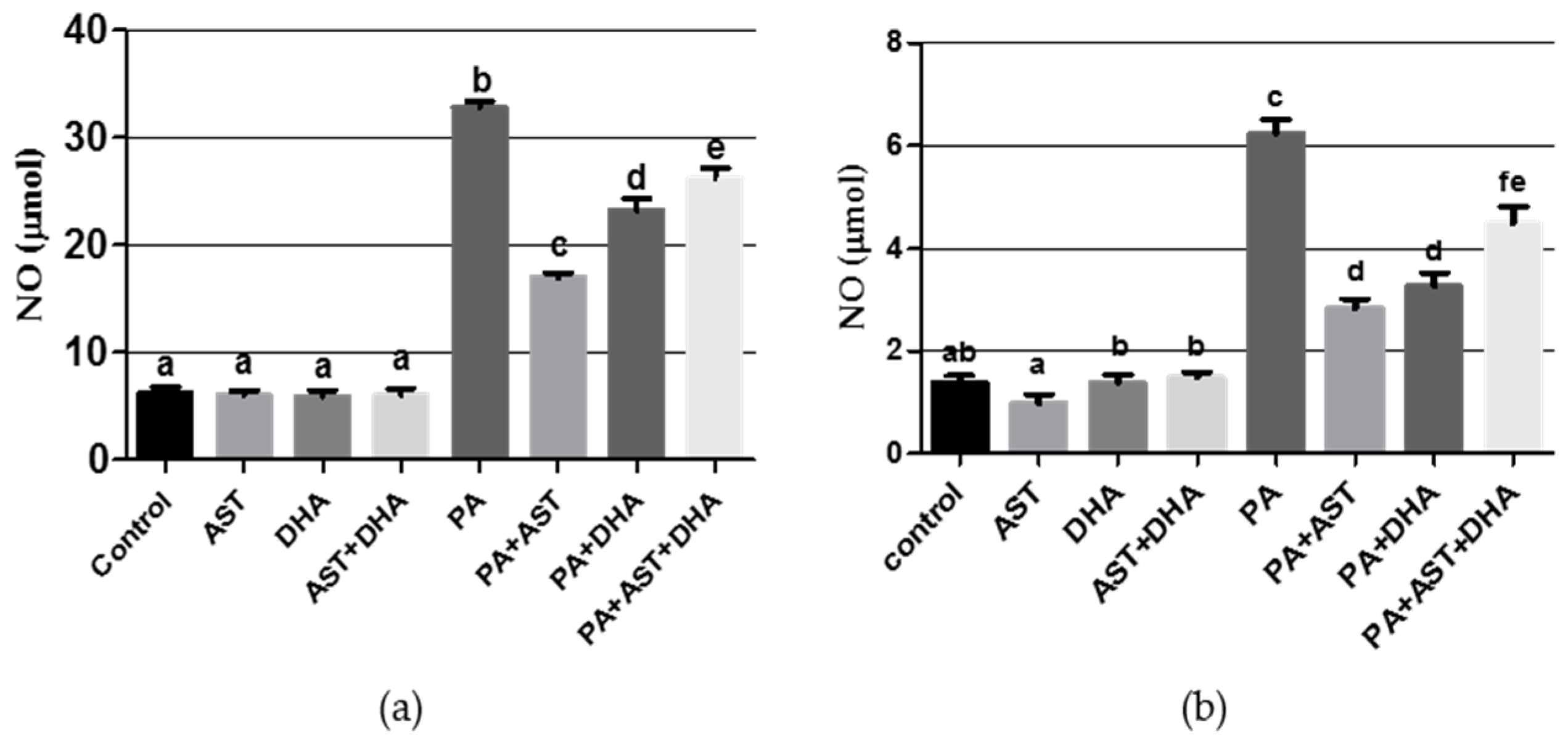
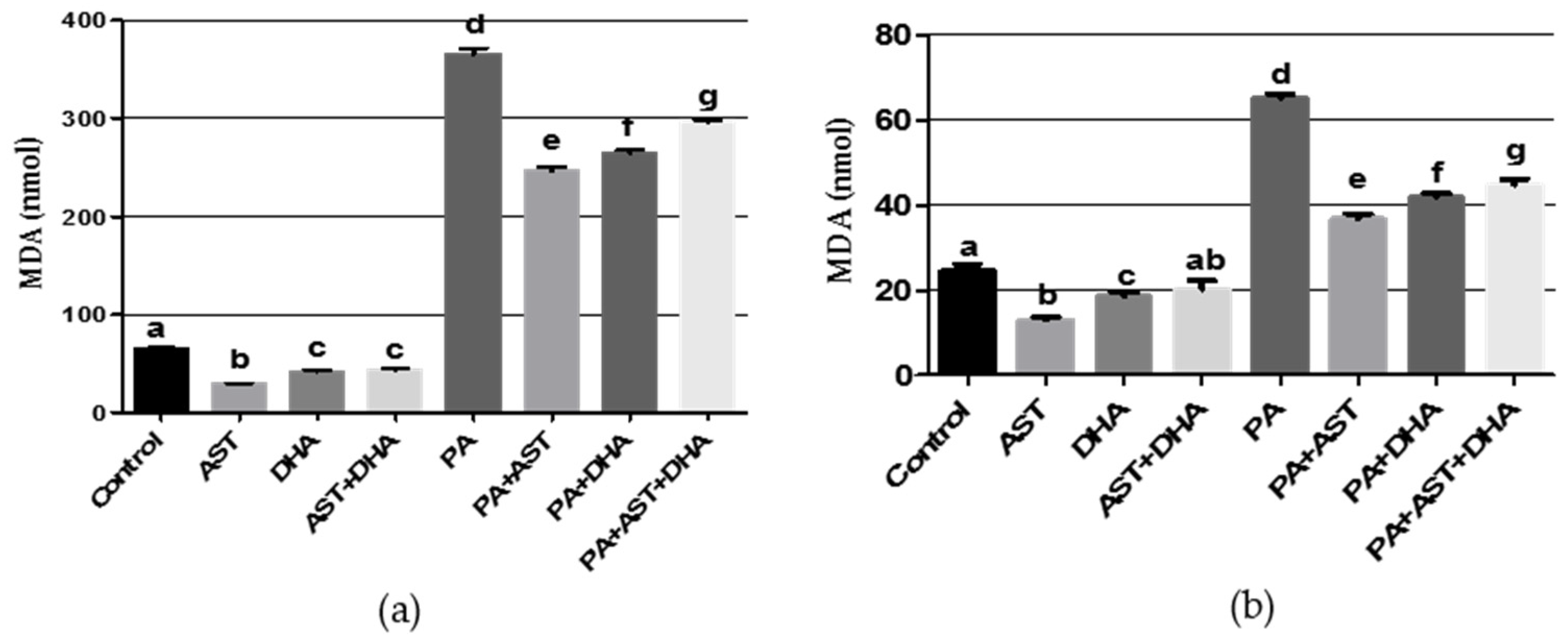
2.3.3. DHA and AST Significantly Decreased the Oxidative Stress Markers Nitric Oxide (NO) and Malondialdehyde (MDA) in PA-Treated Sprague Dawley Rat Serum or Tissue Homogenates
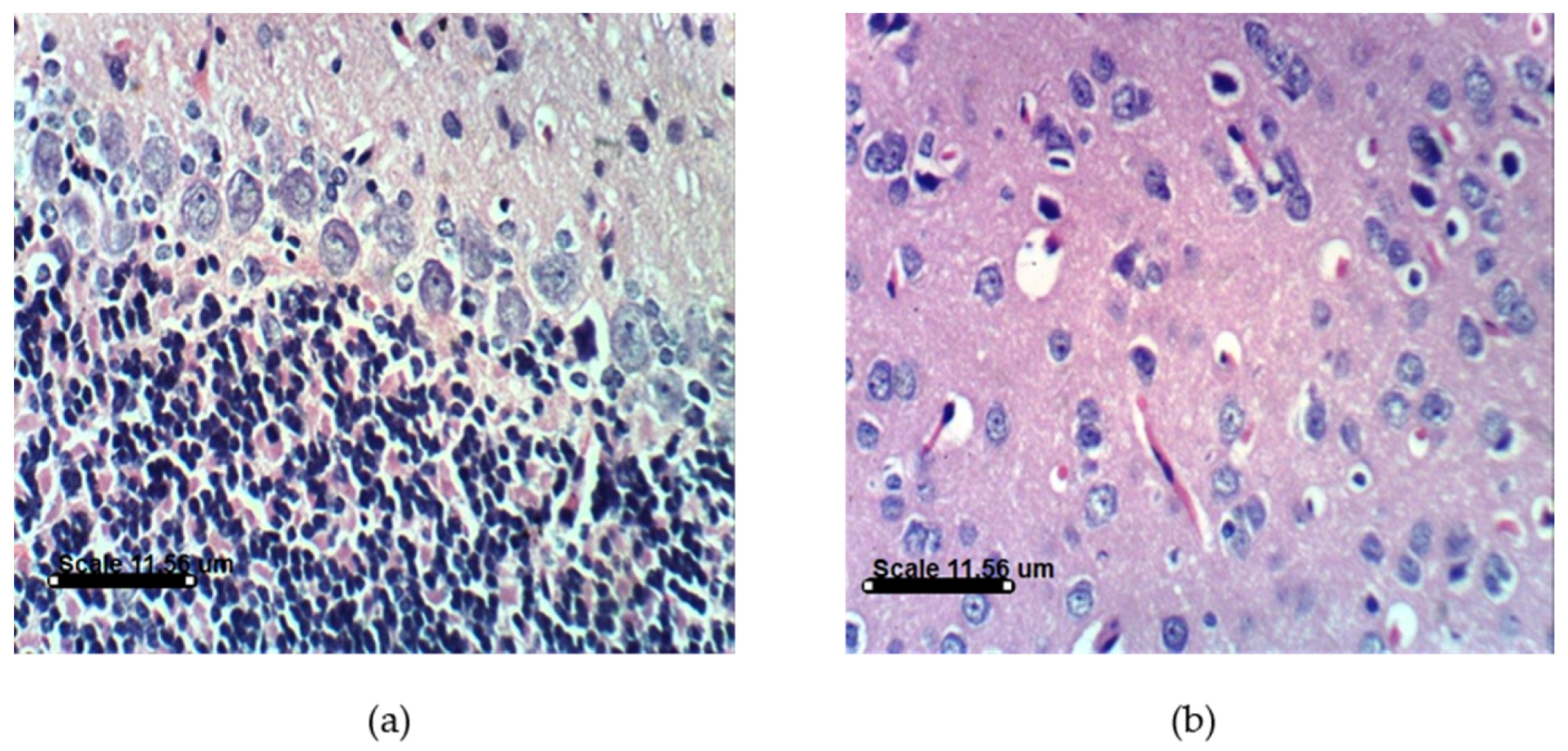
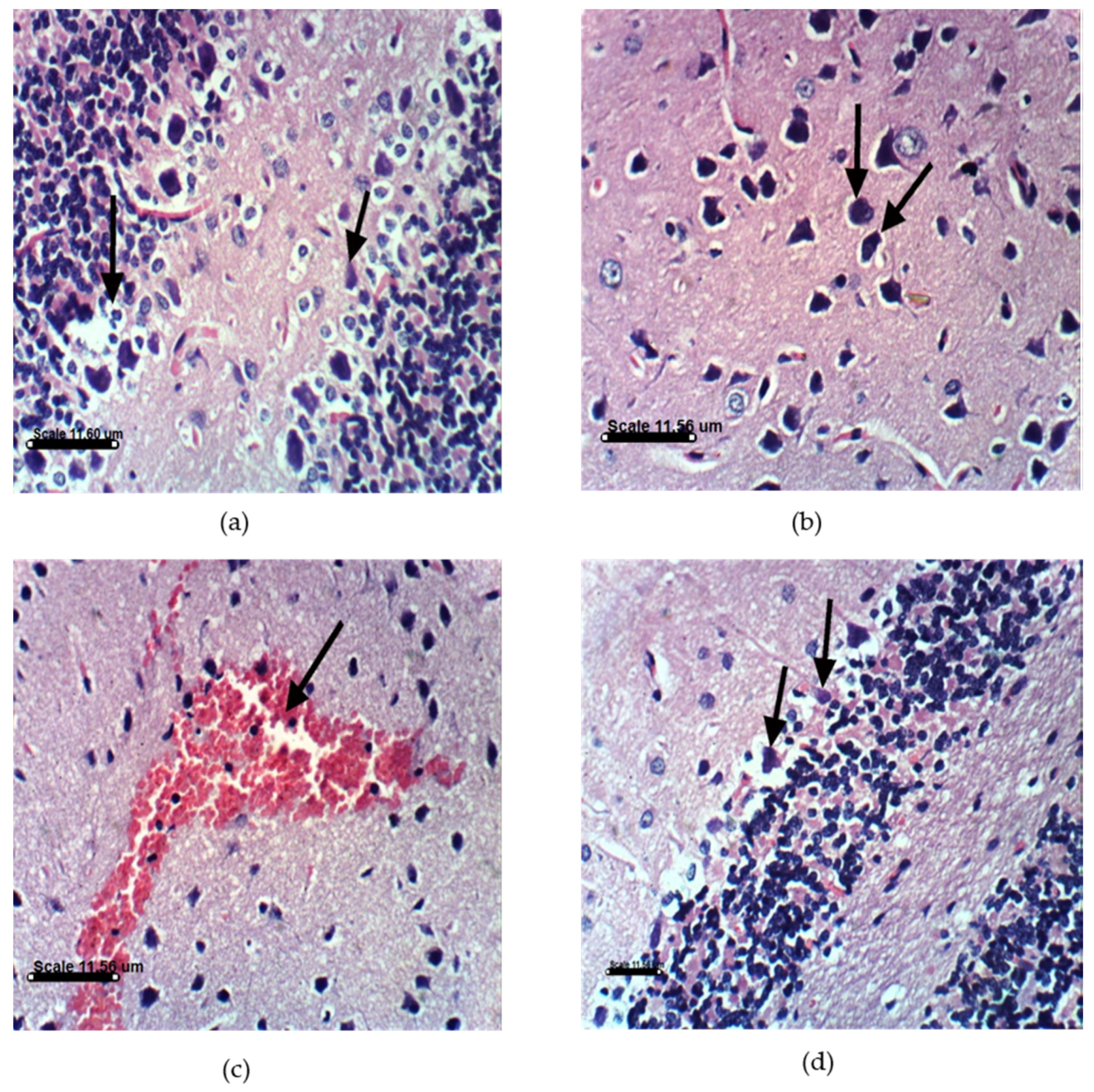
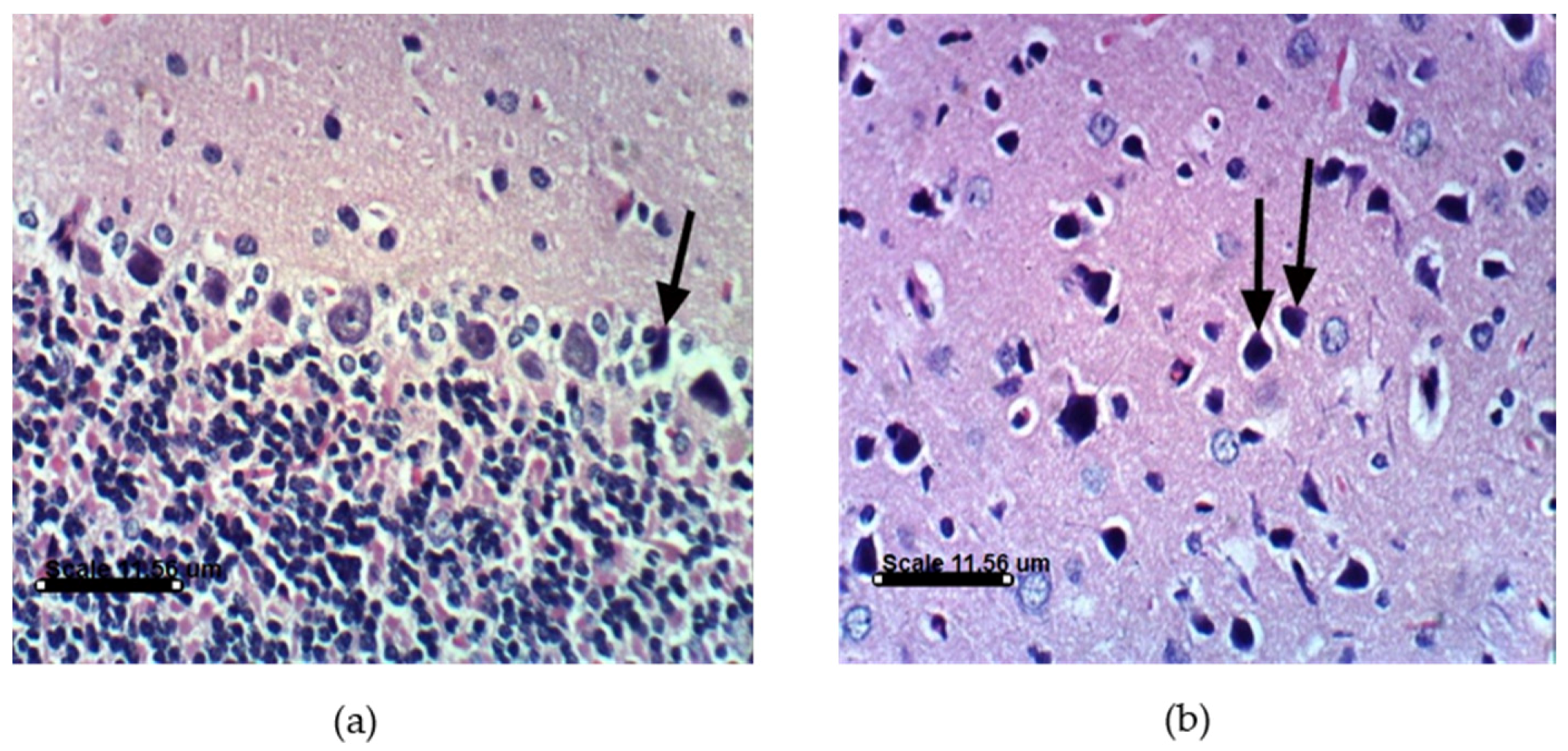
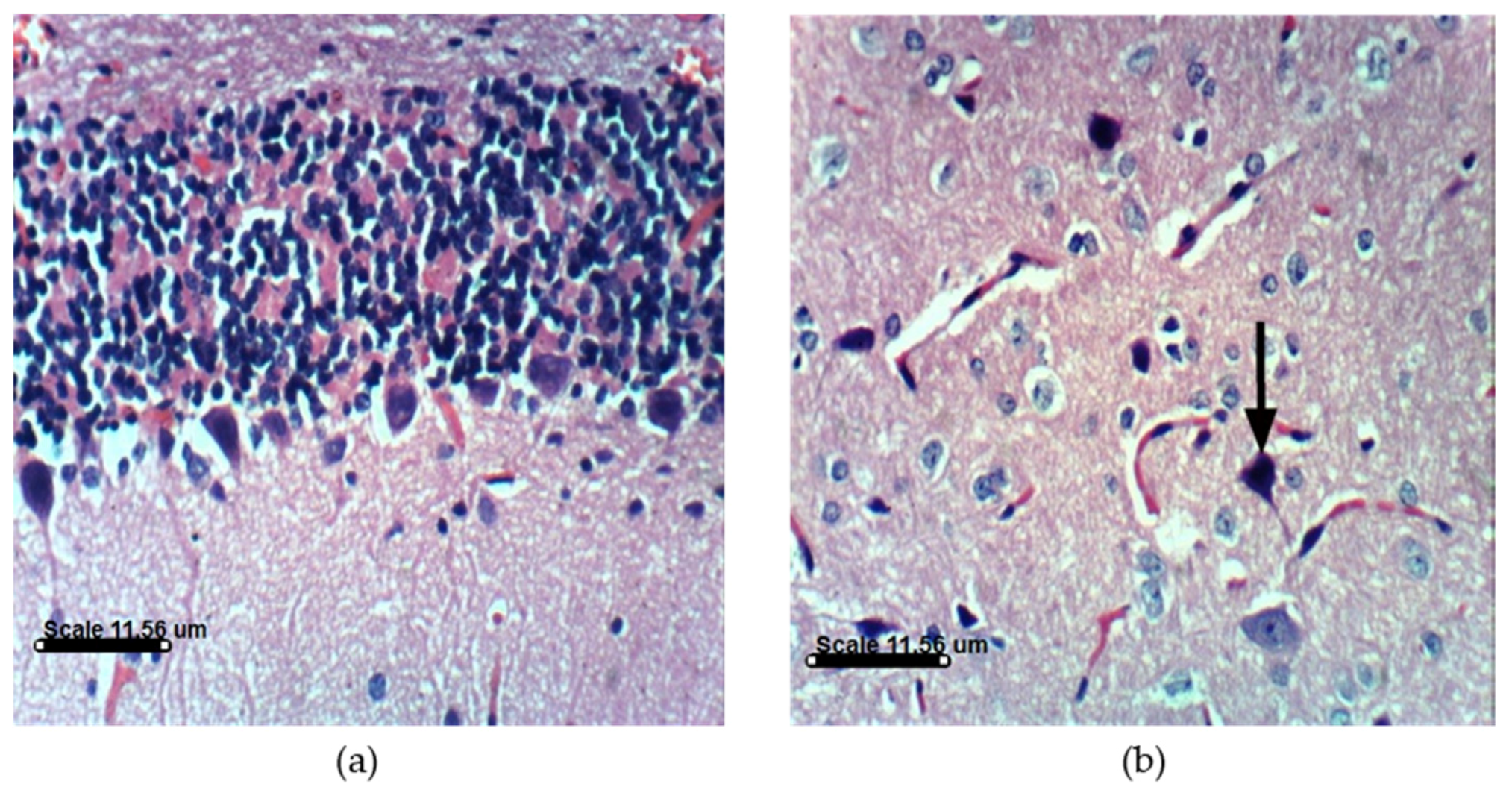
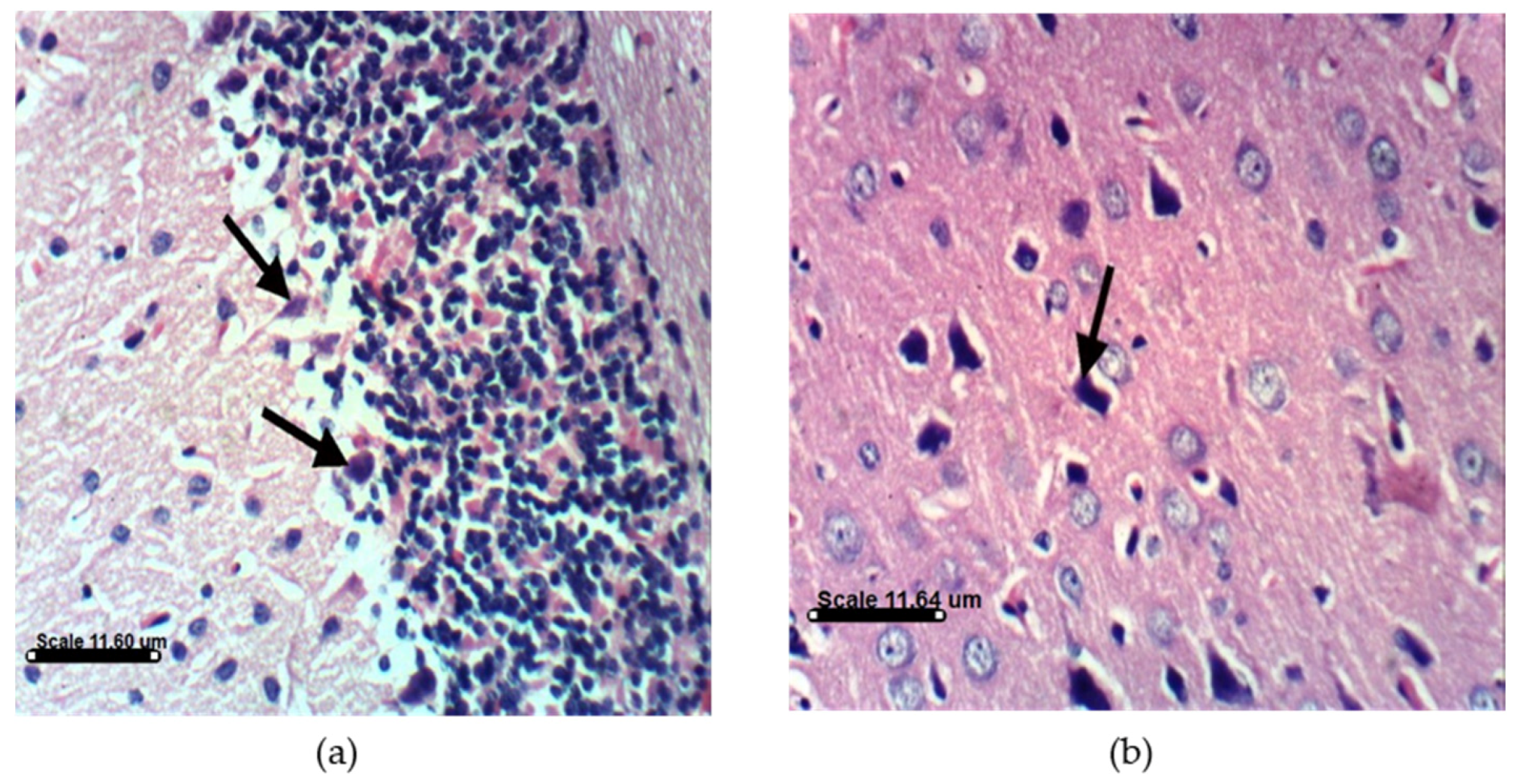
2.3.4. AST and DHA Prevented PA-Induced Histopathological Changes in Sprague Dawely Rat Brain (Cerebrum and Cerebellum)
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. PA Production
4.3. PA Isolation and HPLC Quantitation
4.4. Proliferation Assay
4.5. Caenorhabditis elegans Strains
4.5.1. Preparation of Assay Plates
4.5.2. Locomotion Assay: Number of Reversals Count
4.6. Sprague Dawley Rats
4.6.1. Histopathological Examination
4.6.2. Determination of Brain Amino Acids and Monoamines
4.6.3. Measurement of Total Antioxidant Capacity in Serum and Brain Homogenate
4.6.4. Determination of Nitric Oxide in Serum and Brain Homogenate
4.6.5. Assessment of Lipid Peroxidation by Measurement of MDA in Brain Tissue
4.6.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AST | Astaxanthin |
| AUC | Area Under the Curve |
| BBB | Blood Brain Barrier |
| C. elegans | Caenorhbdities elegans |
| CNS | Central Nervous System |
| DA | Dopamine |
| DHA | Docosahexaenoic Acid |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| DMSO | Dimethyl Sulfoxide |
| ASP | Aspartate |
| GLU | Glutamate |
| GABA | Gamma-Amino Butyric Acid |
| 5-HT | 5-Hydroxytryptamine (Serotonin) |
| HPLC | High Performance Liquid Chromatography |
| iNOS | Inducible Nitric Oxide Synthase |
| NGM | Nematode Growth Medium |
| NE | Norepinephrine |
| NO | Nitric Oxide |
| MDA | Malondialdehyde |
| P. crustosum | Penicillium crustosum |
| PA | Penitrem A |
| TAC | Total Antioxidant Capacity |
| HPA | Hypothalamic–Pituitary–Adrenal Axis |
| PITC | Phenyl isothiocyanate |
References
- Netz, N.; Opatz, T. Marine indole alkaloids. Mar. Drugs 2015, 13, 4814–4914. [Google Scholar] [CrossRef] [PubMed]
- Cavanag, J.B.; Holton, J.L.; Noltan, C.C.; Ray, C.C.; Naik, D.E.; Mantle, P.G. The effects of tremorgenic mycotoxin penitrem A on the rat cerebellum. Vet. Pathol. 1998, 35, 53–63. [Google Scholar] [CrossRef]
- Angel, M.A.; Thomas, R.; Christiane, K.F.; Gunnar, S.; Eriksen, A.B. Neurotoxicity of Penicillium crustosum secondary metabolites. Toxicon 2012, 60, 1428–1435. [Google Scholar]
- Overy, D.P.; Seifert, K.A.; Savard, M.E.; Frisvad, J.C. Spoilage fungi and their mycotoxins in commercially marketed chestnuts. Int. J. Food Microb. 2003, 15, 69–77. [Google Scholar] [CrossRef]
- El-Banna, A.A.; Leistner, L. Production of penitrem A by Penicillium crustosum isolated from food stuffs. Int. J. Food Microb. 1988, 7, 9–17. [Google Scholar] [CrossRef]
- Lund, F.; Filtenborg, O.; Frisvad, J.C. Associated mycoflora of cheese. Food Microb. 1995, 12, 173–180. [Google Scholar] [CrossRef]
- Campbell, I.M. Secondary metabolism and microbial physiology. Adv. Microb. Physiol. 1984, 25, 1–60. [Google Scholar] [PubMed]
- Eriksen, G.S.; Hultin-Jaderlund, K.; Moldes-Anaya, A.; Schonheit, J.; Bemhoft, A.; Jaeger, G.; Rundberger, T.; Skaar, L. Poisoning of dogs with tremorgenic Penicillium Toxins. Med. Mycol. 2010, 48, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Lewis, P.R.; Donoghue, M.B.; Hocking, A.D.; Cook, L.; Granger, L.V. Tremor syndrome associated with fungal toxin: sequelae of food contamination. Med. J. Aust. 2005, 182, 582–584. [Google Scholar] [PubMed]
- Moldes-Anaya, A.; Fonnum, F.; Eriksen, G.S.; Rundberget, T.; Walaas, S.I.; Wigestrand, M.B. In vitro neuropharmacological evaluation of penitrem-induced tremorgenic syndromes: Importance of the GABAergic system. Neurochem. Int. 2011, 59, 1074–1081. [Google Scholar] [CrossRef] [PubMed]
- Hussein, G.; Sankawa, U.; Goto, H.; Matsumoto, K.; Watanabe, H. Astaxanthin, a carotenoid with potential in human health and nutrition. J. Nat. Prod. 2006, 69, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Shibato, T.; Hisaka, S.; Osawa, T. Astaxanthin inhibits reactive oxygen species-mediated cellular toxicity in dopaminergic SA-SY5Y cells via mitochondria-targeted protective mechanism. Brain Res. 2009, 1254, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, T.; Hosoda, K.; Hirano, R.; Kurata, H.; Matsumoto, A.; Miki, W.; Kamiyama, M.; Itakura, H.; Yamamoto, S.; Konda, K. Inhibition of low density lipoprotein oxidation by astaxanthin. J. Atheroscler. Thromb. 2000, 7, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.K.; Park, Y.S.; Choi, D.K.; Chang, H.I. Effect of astaxanthin on the production of NO and the expression of Cox-2 and i NOS in LPS-stimulated BV2 microglial cells. J. Microb. Biotechnol. 2008, 18, 1990–1996. [Google Scholar]
- Khan, S.K.; Malinski, T.; Mason, R.P.; Kubant, R.; Jacob, R.F.; Fujioka, K.; Denstaedts, J.; King, T.J.; Jackson, H.L.; Hieber, A.D.; et al. Novel astaxanthin prodrug (CDX-085) attenuates thrombosis in a mouse model. Thromb. Res. 2010, 126, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Guertin, M.; Huntley, M.E.; Olaizola, M. Haematococcus astaxanthin application for human health and nutrition. Trends Biotechnol. 2003, 21, 210–216. [Google Scholar] [CrossRef]
- Hammond, H.G.; Mayhew, D.A.; Naylor, M.W.; Ruecker, F.A.; Mast, R.W.; Sander, W.J. Safety assessment of DHA-rich microalgae from Schizochytrium Sp.: I. Subchronic rat feeding study. Regul. Toxicol. Pharmacol. 2001, 33, 192–204. [Google Scholar] [CrossRef] [PubMed]
- Caviello, G.; Palozza, P.; Piccioni, E.; Maggiano, N.; Frattuci, A.; France-Schelli, P.; Bartoli, G.M. Dietary supplementation with eicosapentaenoic and docosahexaenoic acid inhibit growth of Morris hepatocarcinoma 3924A in rats: Effects on proliferation and apoptosis. Int. J. Cancer 1998, 75, 699–705. [Google Scholar] [CrossRef]
- Amminger, G.P.; Schafer, M.R.; Papageorgiou, K.; Klier, C.M.; Cotton, S.M.; Mackinnon, A.; MCGorry, P.D.; Berger, G.E. Long-omega-3 fatty acids for indicated prevention of psychotic disorders: A randomized, placebo-controlled trial. Arch. Gen. Psychiatry 2010, 67, 146–154. [Google Scholar] [CrossRef] [PubMed]
- McNamara, R.K. Evaluation of docosahexaenoic acid deficiency as a preventable risk factor for recurrent affective disorders: Current status, future directions and dietary recommendations. Prostaglandins Leukot. Essent. Fat. Acids 2009, 81, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Sanford, M.T.; Xin, Z.; Lin, G.; Lue, T.F. Role of Schwann cells in the regeneration of penile and peripheral nerves. Asian J. Androl. 2015, 17, 776–782. [Google Scholar] [PubMed]
- Sallam, A.; Ayoub, N.; Foudah, A.; Gissendanner, C.; Meyer, S.; El Sayed, K. Indole diterpenoids as novel inhibitors of the wnt/β-catenin pathway in breast cancer cells. Eur. J. Med. Chem. 2013, 70, 594–606. [Google Scholar] [CrossRef] [PubMed]
- Sallam, A.A.; Houssen, W.E.; Gissendanner, C.R.; Orabi, K.Y.; Foudah, A.I.; El Sayed, K.A. Bioguided discovery and pharmacophore modeling of the mycotoxic indole diterpene alkaloids penitrems as breast cancer proliferation, migration, and invasion inhibitors. Med. Chem. Commun. 2013, 4, 1360–1369. [Google Scholar] [CrossRef] [PubMed]
- Welz, C.; Krueger, N.; Schniederjans, M.; Miltsch, S.M.; Kruecken, J.; Guest, M.; Holden-Dye, L.; Harder, A.; von Samson-Himmelstjerna, G. SLO-1 channels of parasitic nematodes reconstitute locomotor behavior and emodepside sensitivity in Caenorhabditis elegans slo-1 loss of function mutants. PLoS Pathog. 2011, 7, e1001330. [Google Scholar] [CrossRef] [PubMed]
- Hoshi, T.; Xu, R.; Hou, S.; Heinemann, S.H.; Tian, Y. A point mutation in the human Slo-1 channel that impairs its sensitivity to omega-3-docosahexaenoic acid. J. Gen. Physiol. 2013, 142, 507–522. [Google Scholar] [CrossRef] [PubMed]
- Moldes-Anaya, A.; Rundberget, T.; Fæste, C.K.; Eriksen, G.S.; Bernhoft, A. Neurotoxicity of Penicillium crustosum secondary metabolites: Tremorgenic activity of orally administered penitrem A and thomitrem A and E in mice. Toxicon 2012, 60, 1428–1435. [Google Scholar] [CrossRef] [PubMed]
- Hoshi, T.; Tian, Y.; Xu, R.; Heinemann, S.H.; Hou, S. Mechanism of the modulation of BK potassium channel complexes with different auxiliary subunit compositions by the omega-3 fatty acid DHA. Proc. Natl. Acad. Sci. USA 2013, 110, 4822–4827. [Google Scholar] [CrossRef] [PubMed]
- Hoshi, T.; Wissuwa, B.; Tian, Y.; Tajima, N.; Xu, R.; Bauer, M.; Heinemann, S.H.; Hou, S. Omega-3 fatty acids lower blood pressure by directly activating large conductance Ca2+-dependent K+ channels. Proc. Natl. Acad. Sci. USA 2013, 110, 4816–4821. [Google Scholar] [CrossRef] [PubMed]
- Hussein, G.; Nakamura, M.; Zhao, Q.; Iguchi, T.; Goto, H.; Saakwa, U.; Watanabe, H. Antihypertensive and neuroprotective effects of astaxanthin in experimental animals. Biol. Pharm. 2004, 28, 47–52. [Google Scholar] [CrossRef]
- Lee, U.S.; Cu, J. BK channel activation: Structural and functional insights. Neuroscience 2010, 33, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Potkonjak, D.; Filipovic, N.; Valjevac, K. The Effect of Mycotoxin Penitrem A on the Concentration of Acetylcholine, Dopamine, Noradrenaline and 5-Hydroxytryptamine in Rat Brain. In Abstracts: Proceedings of the Seventh International Congress of Pharmacology; Boissier, J.R., Lechat, P., Fichelle, J., Eds.; Pergamon Press: London, UK, 1978; p. 574. [Google Scholar]
- De Lau, L.M.I.; Breteler, M.M. Epidemiology of Parkinson’s disease. Lancet Neurol. 2006, 5, 525–535. [Google Scholar] [CrossRef]
- Bousquet, M.; Gue, K.; Emond, V.; Julien, P.; Kang, J.X.; Cicchetti, F.; Calon, F. Transgenic conversion of omega-6 into omega-3 fatty acids in a mouse model of Parkinson’s disease. J. Lipid Res. 2011, 52, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Salem, N.; Litman, B.; Kim, H.Y.; Gawrisch, K. Mechanisms of actions of docosahexaenoic acid in the nervous system. Lipids 2001, 36, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.H.; Liang, Q.Y.; Shi, Y. Pure docosahexaenoic acid can improve depression behaviors and affect HPA axis in mice. Eur. Rev. Med. Pharmacol. Sci. 2012, 16, 1765–1773. [Google Scholar] [PubMed]
- Paterniti, I.; Impellizzeri, D.; Di Paolo, R.; Esposito, E.; Gladman, S.; Yip, P.; Priestley, J.V.; Michael-Titus, A.T.; Cuzzocrea, S. Docosahexaenoic acid attenuates the early inflammatory response following spinal cord injury in mice: In-Vivo and in-Vitro studies. J. Neuroinflamm. 2014, 11, 6. [Google Scholar] [CrossRef] [PubMed]
- Coulombe, K.; Saint-Pierre, M.; Cisbani, G.; St-Amour, I.; Gibrat, C.; Giguère-Rancourt, A.; Calon, F.; Cicchetti, F. Partial neurorescue effects of DHA following a 6-OHDA lesion of the mouse dopaminergic system. J. Nutr. Biochem. 2016, 30, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.R.; Sindhuja, H.N.; Dharmesh, S.M.; Sankar, K.U.; Sarada, R.; Ravishankar, G.A. Effective inhibition of skin cancer, tyrosinase, and antioxidative properties by astaxanthin and astaxanthin esters from the green alga Haematococcus pluvialis. J. Agric. Food Chem. 2013, 61, 3842–3851. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Shibata, T.; Hisaka, S.; Osawa, T. Astaxanthin inhibits reactive oxygen species-mediated cellular toxicity in dopaminergic SH-SY5Y cells via mitochondria-targeted protective mechanism. Brain Res. 2009, 1254, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Kielar, M.L.; Jegarajah, D.R.; Penfield, J.G.; Lu, C.Y. Docosahexaenoic acid decreases IRF-1 mRNA and thus inhibits activation of both the IRF-E and NFκd response elements of the iNOS promoter. Transplantation 2000, 69, 2131–2137. [Google Scholar] [CrossRef] [PubMed]
- Fassett, R.; Coombes, J. Astaxanthin: A potential therapeutic agent in cardiovascular diseases. Mar. Drugs 2011, 9, 447–465. [Google Scholar] [CrossRef] [PubMed]
- Breton, P.; Bizot, J.C.; Buee, J.; De La Manche, I. Brain neurotoxicity of penitrem A: Electrophysiological, behavioral and histopathological study. Toxicon 1998, 36, 645–655. [Google Scholar] [CrossRef]
- Wang, H.Q.; Sun, X.B.; Xu, Y.X.; Zhao, H.; Zhu, Q.Y.; Zhu, C.Q. Astaxanthin upregulates heme oxygenase-1 expression through ERK1/2 pathway and its protective effect against β-amyloid-induced cytotoxicity in SH-SY5Y cells. Brain Res. 2010, 1360, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, M.; Joshi, D.K.; Tripathi, S.; Kulshrestha, S.; Mahdi, A.A. Docosahexaenoic acid ameliorates aluminum-induced biochemical and morphological alteration in rat cerebellum. Ann. Neurosci. 2014, 21, 5–9. [Google Scholar] [PubMed]
- Rundberget, T.; Wilkins, A.L. Determination of Penicillium toxins in food and feeds using liquid chromatography-mass spectrometry. J. Chromatogr. A 2002, 964, 189–197. [Google Scholar] [CrossRef]
- Katagiri, M.; Satoh, A.; Tsujis, S.; Shirasawa, T. Effect of astaxanthin-rich Haematococcus pluvialis extract on cognitive function: A randomized, double-blind, placebo-controlled study. J. Clin. Biochem. Nutr. 2012, 51, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Bancraft, J.D.; Stevens, A.; Turner, D.R. Theory and Practice of Histological Techniques, 4th ed.; Bancroft, J.D., Stevens, A., Eds.; Churchill Livingston: Edinburgh, UK, 1996. [Google Scholar]
- Heinrikson, R.L.; Meredith, S.C. Amino acid analysis by RP-HPLC: Precolumn derivatization with phenylisothiocyanate. Anal. Biochem. 1984, 136, 65–74. [Google Scholar] [CrossRef]
- Pagel, P.; Blome, J.; Wolf, H.U. High-performance liquid chromatographic separation and measurement of various biogenic compounds possibly involved in the pathomechanism of Parkinson’s disease. J. Chromatogr. B 2000, 746, 297–304. [Google Scholar] [CrossRef]
- Koracevic, D.; Koracevic, G.; Djordjevic, V.; Andrejevic, S.; Cosic, V. Method for measurement of antioxidant activity in human fluids. J. Clin. Pathol. 2001, 54, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, H.A.C.; Dymock, J.F. The determination of nitrite in water. Analyst 1961, 86, 414–416. [Google Scholar]
- Ruiz-Larrea, M.B.; Leal, A.M.; Liza, M.; Lacort, M.; de Groot, H. Antioxidant effects of estradiol and 2-hydroxyestradiol on iron induced lipid peroxidation of rat liver microsome. Steroids 1994, 59, 383–388. [Google Scholar] [PubMed]














© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goda, A.A.; Naguib, K.M.; Mohamed, M.M.; Amra, H.A.; Nada, S.A.; Abdel-Ghaffar, A.-R.B.; Gissendanner, C.R.; El Sayed, K.A. Astaxanthin and Docosahexaenoic Acid Reverse the Toxicity of the Maxi-K (BK) Channel Antagonist Mycotoxin Penitrem A. Mar. Drugs 2016, 14, 208. https://doi.org/10.3390/md14110208
Goda AA, Naguib KM, Mohamed MM, Amra HA, Nada SA, Abdel-Ghaffar A-RB, Gissendanner CR, El Sayed KA. Astaxanthin and Docosahexaenoic Acid Reverse the Toxicity of the Maxi-K (BK) Channel Antagonist Mycotoxin Penitrem A. Marine Drugs. 2016; 14(11):208. https://doi.org/10.3390/md14110208
Chicago/Turabian StyleGoda, Amira A., Khayria M. Naguib, Magdy M. Mohamed, Hassan A. Amra, Somaia A. Nada, Abdel-Rahman B. Abdel-Ghaffar, Chris R. Gissendanner, and Khalid A. El Sayed. 2016. "Astaxanthin and Docosahexaenoic Acid Reverse the Toxicity of the Maxi-K (BK) Channel Antagonist Mycotoxin Penitrem A" Marine Drugs 14, no. 11: 208. https://doi.org/10.3390/md14110208
APA StyleGoda, A. A., Naguib, K. M., Mohamed, M. M., Amra, H. A., Nada, S. A., Abdel-Ghaffar, A.-R. B., Gissendanner, C. R., & El Sayed, K. A. (2016). Astaxanthin and Docosahexaenoic Acid Reverse the Toxicity of the Maxi-K (BK) Channel Antagonist Mycotoxin Penitrem A. Marine Drugs, 14(11), 208. https://doi.org/10.3390/md14110208