1. Introduction
Fish are the predominant source of the omega-3 (n-3) long-chain polyunsaturated fatty acids (LC-PUFAs), eicosapentaenoic acid (EPA, 20:5n-3) and docosahexaenoic acid (DHA, 22:6n-3) which play a unique role in human nutrition, health, and development [
1]. To produce fish rich in EPA and DHA, aquafeeds contain fish meal and fish oil obtained from wild catches. Current trends demanding a more sustainable and economic fish production have led to an increased development and inclusion of alternative terrestrial and plant ingredients in aquafeeds. Depending on the alternative ingredients used, the health and nutritious properties of the farmed fish may be affected [
2]. The replacement of dietary fish oil by vegetable oils might be feasible in some species, but can be problematic especially in carnivorous marine species [
2]. The gilthead sea bream (
Sparus aurata) is a carnivorous marine fish species of economic importance especially in the Mediterranean region [
3]. Sustainable diet formulations with low levels of fish oil do not necessarily affect the growth and performance, but impair the fillet quality and fatty acid (FA) composition of sea bream and European sea bass (
Dicentrarchus labrax) [
4,
5,
6,
7]. The FA composition of marine species usually reflects that of their diet, since the ability to convert dietary C18 precursor fatty acids to LC-PUFAs is hardly present in marine finfish [
8,
9]. Nowadays, farmed gilthead sea bream can have decreased EPA and DHA contents per gram of fillet in comparison to past years due to fish oil replacement [
5], although the content in farmed sea bream is higher than in the wild specimens [
4].
To counteract this reduction trend, recovering the EPA and DHA levels, and to wash out undesirable FA of vegetable origin, finishing diets high in fish oil have been successfully used in sea bream [
4,
6]. Furthermore, the use of microalgae containing LC-PUFA in the diets of marine finfish, such as
Pavlova viridis in European sea bass diets [
10], is also a promising approach. The use of
de novo n-3 oils from genetically modified oil crops, for example, in Atlantic salmon (
Salmo salar) and sea bream [
11,
12], is innovative, but due to legislation, it is not practicable in all countries.
The aforementioned approaches have one thing in common: They rely on already existing sources of EPA and DHA that are supplied to the fish via the diet. Thus, any improvement of the innate ability of the fish to cope with diets low in EPA and DHA would be an interesting alternative. The modification of the underlying molecular mechanism and exploitation of genetic capacities build the backbone for this approach. In freshwater fish that have a limited natural ability to convert the precursor C18 FA α-linolenic acid (ALA, 18:3n-3) to EPA and DHA [
13,
14], bioactive phytochemicals might stimulate this FA synthesis. An elevation of the EPA and DHA levels has successfully been shown in rainbow trout (
Oncorhynchus mykiss) fed diets containing resveratrol [
15] and sesamin [
16], in zebrafish (
Danio rerio) embryos exposed to wine polyphenols [
17], and salmon hepatocytes treated with genistein [
18] and sesamin [
19]. Apart from the hardly present in vivo FA bioconversion in marine fish, some species like sea bream and sea bass seem to possess the genetic capacity to perform the synthesis at least partly [
20,
21,
22,
23,
24]. Thus, it might be possible to exploit the genetic capacity of marine finfish and activate the expression of dormant genes encoding proteins involved in the FA synthesis. Bioactive secondary plant compounds that increased the endogenous FA synthesis of freshwater fish might be a promising tool to be investigated in marine finfish.
Resveratrol (RV) is a stilbene derivate produced by plants, mainly grape vines, in response to infections [
25] and has potential health-beneficial, anti-inflammatory, and anti-oxidant properties [
26,
27,
28]. The possible modulation of animal lipid metabolism [
17,
29,
30] and the potential to increases elongase and desaturase (∆6- and ∆5-desaturase) activities [
31] are interesting for its application in fish. The fact that RV indicated the increasing properties of LC-PUFA in mammalian cell cultures [
31,
32], zebrafish [
17], and rainbow trout [
15], makes it an interesting phytochemical for nutrition studies in marine fish.
Additionally, environmental factors may influence the content of n-3 LC-PUFA or the expression and activity of enzymes involved in the FA synthesis in fish [
33,
34,
35]. In freshwater fish and salmonids, it seems that the FA desaturation, elongation, and β-oxidation activities are increased at lower temperatures [
33,
34]. Studies with marine fish reveal controversial results. Vagner et al. [
36] demonstrated that temperature did not affect the diet-induced upregulation of the ∆6-desaturase (∆6-D) in sea bream larvae. In contrast to that, Skalli et al. [
35] demonstrated that the amount of LC-PUFAs increased in the phospholipid fraction in sea bass held at low water temperatures of 22 °C. Since temperature affects diet intake and growth, it seems likely that the FA synthesis might eventually be affected. Thus, an investigation of phytochemical-induced effects in sea bream under the influence of nutritional and environmental factors is highly interesting.
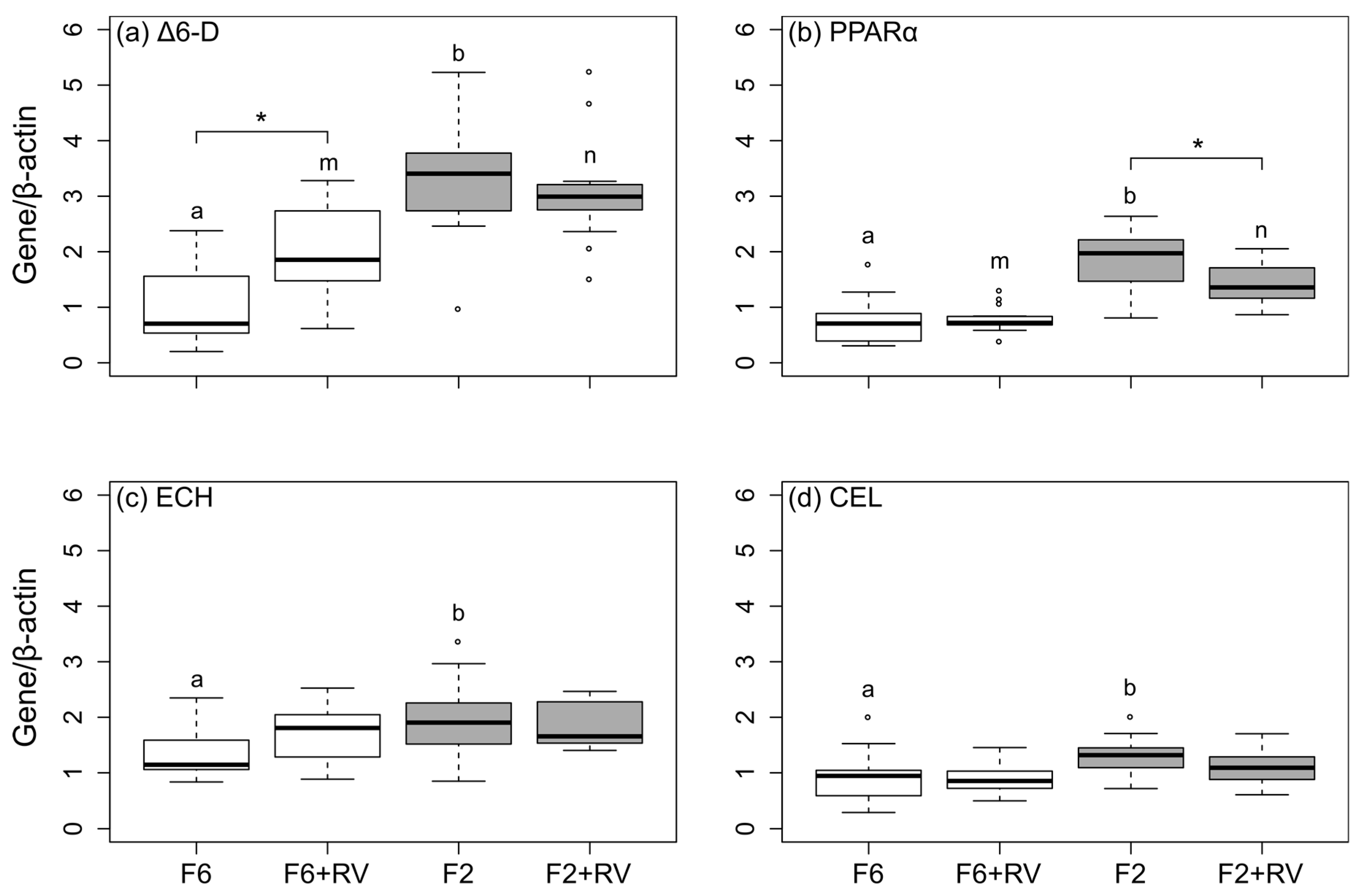
The aim of this study was to investigate the effects of low-fish oil diets supplemented with dietary RV on (1) growth, (2) performance parameters, (3) whole body nutrient composition, (4) FA composition of the whole body, liver and fillet, and (5) the mRNA steady-state levels of selected genes encoding proteins involved in the desaturation (∆6-D), FA metabolism (Carboxyl ester lipase (CEL)), and β-oxidation of FAs (Peroxisome proliferator-activated receptor α (PPARα); Enoyl-CoA hydratase (ECH)) of the gilthead sea bream. Furthermore, we addressed the question whether the holding temperature played a role in the response of the investigated parameters.
4. Materials and Methods
4.1. Experimental Diets
Four different experimental diets (isonitrogenous and isoenergetic) were formulated as shown in
Table 7. All diets consisted of mainly alternative plant protein sources (Soybean concentrate, corn gluten, wheat gluten, and rapeseed expeller) and had a fish meal content of 5% DM. Diet F6, the overall control diet, contained 6% DM fish oil and 3% DM of a mixture of vegetable oils (linseed, rapeseed, and palm oil). Diet F2 had a fish oil content of 2% DM and contained 7% DM of a mixture of linseed, rapeseed, and palm oil. The diets contained considerably different amounts of the fatty acids EPA and DHA as shown in
Table 3. Control diet F6 was formulated to meet the recommended amount of EPA and DHA for sea bream (EPA + DHA: 0.9% of dry diet when DHA/EPA = 1 [
40,
41]). Diet F2 contained reduced amounts of EPA and DHA (0.37% of dry diet,
Table 3) below the recommendation for sea bream. Experimental diets F6 + RV and F2 + RV were equal to diets F6 and F2, respectively, and supplemented with resveratrol (+ RV;
trans-3,4',5-trihydroxy stilbene, purity ≥ 98%, Chemos GmbH & Co. KG, Regenstauf, Germany) at a concentration of 0.15% DM of the diet. The dietary essential amino acid content of each diet was calculated from the amino acid content of the single ingredients. All diets were formulated based on the macronutrient and the essential amino acid requirements of gilthead sea bream according to Peres and Olivia-Teles [
61] and Wilson [
62]. All diets were pressed into 3- and 4-mm pellets using a feed press L14-175 (Amandus Kahl, Reinbek, Germany).
4.2. Experimental Setup
The study was conducted at the facilities of the Grupo de Investigación en Acuicultura, Universidad de Las Palmas de Gran Canaria (GIA-ULPGC), Telde, Las Palmas, Canary Islands, Spain. All experiments were carried out according to the EU Directive 2010/63/EU for animal experiments and approved by the Ministry of Energy, Agriculture, the Environment, Nature and Digitalization (MELUND), Kiel, Germany (approved on 15th October 2014; project number: V244-7224.121.9-34). Two experimental setups were prepared for the feeding trial to realize a simultaneous diet- and temperature-challenge. (1) A flow-through system with 12 tanks was used for the high-temperature challenge (approx. 23 °C). (2) Identical temperature-controlled small recirculating aquaculture systems equipped with a total of 12 tanks were used for the low-temperature challenge (approx. 19 °C).
(1) The flow-through system had a natural photoperiod (12 h light) during the whole adaptation and experimental period. The cylindrical fiberglass tanks (500 L) were supplied with filtered seawater (37‰ salinity) at a rate of 600 L h−1 and were continuously aerated. Temperature (22.6 ± 0.6 °C), oxygen (6.2 ± 0.3 mg L−1), and pH (8.22; pH-Meter Basic 20+, CRISON, Hach Lange Spain, Barcelona, Spain) were monitored daily. Ammonia (NH4: < 0.15) and nitrite (NO2: < 0.1, Royal Ammonia Professional Test and Royal Nitrite Professional Test, Royal Nature, Nesher, Israel) concentrations were determined every second day.
(2) The recirculating systems (kept at a photoperiod of 12 h light) had 3000 L of water volume each and were equipped with a cooling unit and a water treatment unit consisting of a sand filter (filtration area: 0.33 m3; granular size: 0.4–1.2 mm, AstralPool, Fluidra, Barcelona, Spain), a multibed bio filter (filtration area: 0.32 m2; Kripsol, Toledo, Spain), and a protein skimmer. Each recirculating system had three cylindrical fiberglass tanks (500 L) that were supplied with temperature controlled sea water, the same as in the flow-through system. The water quality parameters did not differ between the systems and were measured as described above (temperature: 19.0 ± 1.5 °C; oxygen: 7.1 ± 0.3 mg L−1 O2; pH: 8.06; NH4: <1; NO2: <0.5).
A total of 600 juvenile gilthead sea bream (offspring from brood stock of GIA-ULPGC, initial body weight: 12.5 ± 2.2 g) were acclimated in the flow-through system and randomly and equally distributed among all 24 tanks of both setups, 25 individuals per tank. Over the first seven days of the experimental period, the recirculating systems were continuously cooled down from 23 °C to 19 °C. The four dietary treatments (F6, F6 + RV, F2, and F2 + RV) were randomly distributed among the twelve tanks of each setup, ensuring that each dietary treatment was tested at each temperature in a triplicate approach. During the whole experimental period of 55 days, the fish were fed manually three times per day until apparent satiation in both setups. The administered feed ratios were determined daily for the calculation of the DFI.
4.3. Sampling
The tissue samples were collected before the onset of the feeding trial (day 0) and at the end of the trial (day 56). For the initial sampling at day 0, seven fish were sacrificed (pooled sample) and stored at −80 °C for the determination of the whole body nutrient composition and fatty acid composition. Additionally, six fish were sacrificed for the collection of liver and fillet tissue samples. The whole liver was weighed (±0.01 g) for the determination of the HSI (see Equation (1)). For mRNA quantification via quantitative real-time RT-PCR, one part of the liver tissue was preserved in RNALater (Sigma-Aldrich, Taufkirchen, Germany). For the determination of the liver fatty acid composition, the remaining liver parts of all six individuals were pooled in one sample and immediately frozen at −80 °C. The fillets (left side) from all six fish were pooled and frozen at −80 °C for the determination of the fatty acid composition. For the final sampling at day 56, five fish per tank were sacrificed (pooled sample) and stored at −80 °C for the determination of the whole body nutrient composition and the fatty acid composition. Additionally, five fish per tank were sacrificed for the collection of the liver and fillet tissue samples. The procedure was the same as described for the initial sampling.
For the determination of the growth, performance, and nutrient utilization parameters, all fish were individually weighed (±0.1 g) and measured (±0.1 cm) at day 0 and day 56 for the calculation of the initial and final body weights (IBW and FBW, respectively). The SGR, FCR, PER, PPV, and FCF were calculated according to the Equations (2)–(6).
4.4. Nutrient Composition Analysis
The nutrient composition was analyzed in all experimental diets and the whole body homogenates of the gilthead sea bream. Frozen whole body samples were freeze-dried (Alpha 1,2 LDplus and Alpha 1–4 LSC, Martin Christ Gefriertrocknungsanlagen GmbH, Osterode am Harz, Germany) until the weight was stable and homogenized using a cutting mill (GM 200, Retsch, Haan, Germany). Diets were homogenized using a mortar and pestle. Analysis of nutrients and gross energy was done according to the EU guideline (EC) 152/2009 [
63] as described in Torno et al. [
46]. The total carbohydrates were calculated according to formula (7):
4.5. Lipid Extraction and Fatty Acid Composition Analysis
The total lipids were extracted from the whole body homogenates, liver samples, fillet samples, and diet samples according to Folch et al. [
64]. The measurement of fatty acid methyl esters (FAMEs) was performed using a gas chromatograph with a flame ionization detector (Agilent Technologies, Santa Clara, CA, USA). A detailed description is given in Torno et al. [
15]. A 13-FAME standard was used to identify the retention times of the individual FAMEs (
Table 4,
Table 5 and
Table 6). The fatty acid composition was calculated as a percentage of a single FAME relative to the total FAMEs. The internal standard 13:0 methyl ester was used to calculate FAs as a % DM of the diet.
4.6. mRNA Extraction and Quantitative Real-Time RT-PCR
Based on the results of growth, performance, and FA composition measurement of the mRNA steady-state levels was performed only in the liver of fish held at 19 °C. The total mRNA was extracted from the liver samples using the Innuprep RNA Mini Kit (Analytik Jena, Jena, Germany) according to the manufacturer’s protocol as described in Torno et al. [
15]. The primers used and the appropriate annealing temperatures are listed in
Table 8. The transcript expression was quantified by calculating the input copy number using a standard curve. Subsequently, the respective target mRNA steady-state levels of ∆6-D, PPARα, ECH and CEL were normalized to the mRNA levels of the housekeeping gene beta-actin (β-actin). Data are shown as relative mRNA steady-state levels of respective target genes normalized to their internal control (β-actin) following absolute quantification (
Figure 1).
4.7. Statistical Analysis
All statistical analyses were performed using R (version 3.1.3) with an RStudio interface. The packages gdata, multcomp, gplots, nparcomp, nlme, piecewiseSEM, SimComp, and car were used for the graphical and the statistical analysis.
The data evaluation started with the definition of an appropriate statistical model based on a graphical residual analysis of the data and Levene’s test to test for the homoscedasticity of variances: (1) statistical model based on generalized least squares (
gls) for normally distributed and heteroscedastic data (IBW, FCR, crude protein) and (2) linear model (
lm) for normally distributed and homoscedastic data (DFI, FBW, FCF, HIS, SGR, PER, PPV, crude ash, crude fat, gross energy, dry matter, and fatty acid composition). Both models included the level of dietary fish oil content (6% and 2% DM), supplement (None, RV), and temperature (19 °C, 23 °C), as well as their interaction term as fixed factors. An analysis of variances (ANOVA) was conducted followed by an appropriate post-hoc test: (1) ANOVA based on
gls was followed by multiple contrast tests for heteroscedastic data according to Hasler and Hothorn [
65]; (2) ANOVA based on
lm was followed by multiple contrast tests according to Schaarschmidt and Vaas [
66]. All post-hoc tests compared the feeding groups based on the fish oil level and supplement within one holding temperature and between the two temperatures.
For mRNA steady-state levels (∆6-D, PPARα, ECH, CEL) the data evaluation was initiated with the definition of an appropriate mixed model (
lme) with a fish oil content (6% and 2% DM), supplement (None, RV) and their interaction term as fixed factors, and fish tank as random factor. A residual analysis revealed the data to be non-normally distributed. Multiple contrast tests for relative effects were conducted in order to compare the influence of varying factors [
66].

 ,
, 



{kind=link}