Biosynthesis and Secretion of Human Tissue Kallikrein in Transgenic Chlamydomonas reinhardtii

1
Guangdong Technology Research Center for Marine Algal Bioengineering, Guangdong Key Laboratory of Plant Epigenetic, College of Life Sciences and Oceanography, Shenzhen University, Shenzhen 518060, China
2
Shenzhen Key Laboratory of Marine Bioresource and Eco-environmental Science, Shenzhen Engineering Laboratory for Marine Algal Biotechnology, College of Life Science and Oceanography, Shenzhen University, Shenzhen 518060, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work
Mar. Drugs 2018, 16(12), 493; https://doi.org/10.3390/md16120493
Submission received: 23 November 2018
/
Revised: 3 December 2018
/
Accepted: 4 December 2018
/
Published: 7 December 2018
(This article belongs to the Special Issue Pre-Clinical Marine Drug Discovery)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The green alga Chlamydomonas reinhardtii was recently been shown to be an effective bio-manufacturing platform for the production of recombinant proteins. The advantage of using C. reinhardtii is that it is fast to grow, inexpensive to culture, and relatively safe. However, the expression of foreign proteins is always low and difficult to purify in C. reinhardtii. Human kallikrein has the potential to be developed into certain drugs, like insulin. Therefore, its biosynthesis is important to drug development. In this study, we synthesized the sg gene, a signal peptide sequence of alkaline phosphatase, and inserted it into a pH124 plasmid, which contains a HSP70A-RBCS2 promoter and a RBCS2 terminator. Then, we inserted the human kallikrein gene klk1 behind the sg sequence to make a pHsgk124 vector. The pHsgk124 were transferred into a cell-wall deficient strain of C. reinhardtii, cc-503, by using the glass bead method. Southern blot analysis showed that sg and klk1 were incorporated into genes of the transgenic C. reinhardtii. RT-PCR analysis showed that it had an active transcription and its expression increased three times under heat stress. Western blot analyses of proteins inside and outside cells (in the culture medium) showed that klk1 was expressed in the cell and the resulting protein was secreted into medium. An enzyme activity assay showed that the recombinant protein had the ability to hydrolyze the specific substrate H-D-Val-Leu-Arg-Pna. In conclusion, we successfully bioengineered C. reinhardtii to produce and secrete human kallikrein protein, which has important biomedical implications.
1. Introduction
Chlamydomonas reinhardtii is a type of green algae grown in freshwater. Since it is relatively safe, easy to culture and inexpensive to maintain, it has become a preferred model organism for biotechnology and bioengineering studies. With a well-characterized genetic profile, it has become an important bio-platform to express and produce recombinant protein and antibodies [1,2,3,4,5,6]. Many exogenous genes have been inserted and expressed in C. reinhardtii successfully, and some have been planned in commercial production, such as biodegradable plastic poly-3-hydroxybutyrate [7] and the human antibody IgG1 [8]. In recent years, our group successfully expressed exogenous genes ble and egfp in the mitochondria of C. reinhardtii [9,10].
Although transgene technology is well developed for nuclear, chloroplast [11] and mitochondria genomes in C. reinhardtii, the low protein expression is a major hurdle for developing it into a commercially viable bio-platform. Current recombinant proteins made by C. reinhardtii are mainly expressed inside the cells. When harvesting proteins, one needs to go through complicated cell breaking, purification, and re-purification processes. This not only reduces the yield of recombinant protein, but also sometimes causes its structural change. Using signal peptides [12] to construct a secreting vector to express exogenous genes has the advantage of secreting recombinant protein outside the cells, which facilitates the target protein’s collection and purification. The most successful applications of signal peptide are in bacteria and yeast. For example, the human growth hormone (20 kDa hGH) gene has been successfully incorporated into the E.coli genome to produce a secreted product [13], human α1-antitrypsin has been produced and secreted by Saccharomyces cerevisiae using inulinase signal, and human insulin has been expressed and secreted by Pichia pastoris [14]. There are many reports that C. reinhardtii as a platform can produce recombinant protein, however, most recombinant proteins are accumulated intracellularly, but there are few reports in the literature about using C. reinhardtii to produce secretory recombinant protein [15]. We performed the signal peptide analysis of the C. reinhardtii PHOX sequence (GenBank accession number: xp_001703098.1) and predicted that this signal peptide can guide the synthesis of PHOX protein from the nucleus to the extracellular, which is the foundation of secretory recombinant protein production.
Human tissue kallikreins (htKs) are widely expressed in various tissues and are a secretory protein mainly from the pancreas and kidney [16]. The htKs are encoded by 15 similar steroid hormone-regulated genes and the main one is klk1. The htK encoded by klk1 is a single-chain serine protease that is synthesized as a preproenzyme. It is then cleaved off the signal peptide to become a pro-enzyme to be secreted. The cleavage of pro-peptide makes it an active enzyme [17]. The primary activity of htK is to cleave low molecular weight kininogen to release lysyl-bradykinin (LBK). LBK binds to its receptors B1 and B2 in target tissues to mediate various physiological processes, such as blood pressure regulation, smooth muscle contraction, neural plasticity, pain induction, vascular permeability, vascular cell growth, electrolyte balancing, and inflammatory cascades [18,19]. LBK also has a role in pregnancy maintenance. Dysregulated htK is associated with various diseases, including cancer. Hence, htK is a valuable protein which has potential in the treatment of hypertension, diabetes, cancer and nephropathy.
In this study, we constructed a secretory vector carrying an exogenous klk1 gene, inserted it into C. reinhardtii, analyzed the gene and protein expression in C. reinhardtii, and measured resulting htK in the culture medium.
2. Results
2.1. DNA Verification of Transgenic Algae
PCR-electrophoresis results, including a wild strain sample, three transgenic samples of C. reinhardtii, (named sgk1, sgk 2 and sgk3, respectively) and a negative control are shown in Figure 1. The transfected genome was named sgk (klk1 genome with an upstream signal peptide genome). Transgenic cells showed extra 780-bp segments on the gel. This confirms that the klk1 gene was incorporated into the cc-503 genome. A further total DNA Southern blot (Figure 2) shows that a band appeared just below 1584 bps in the transfected cells but not in the non-transfected cells. The theoretical DNA segment of HSP70A-RBCS2-sg-klk1-RBCS2 contains 1511 bps. The experimental conditions may have slightly affected the band position, furthermore, a correct sequence have to show in Southern blot. Therefore, the band appearance near 1584 bps should confirm that the segment of HSP70A-RBCS2-sg-klk1-RBCS2 was successfully incorporated into the cc-503 genome.
2.2. Western Blot Analysis of Expressed Protein
A Western blot assay was used to analyze proteins inside cells of currently transfected cc-503, previously constructed strain k6-7 (successfully expressed klk1 but non-secretory), and non-transfected cc-503. The result is shown in Figure 3. A band at around 55 kDa appeared in all transfected cc-503 cells (sgk1, sgk2, sgk3) and k6-7 (used as a positive control) cells, but not in non-transfected cc-503 cells. The molecular weight was bigger than the theoretical 31 kDa, which may be due to the glycosylation of recombinant proteins.
Next, Western blot assay was again used to analyze proteins in the 50-time concentrated C. reinhardtii cell culture medium to detect excreted extracellular htK protein. The result is shown in Figure 4. In the culture medium of k6-7, the band at 55 kDa disappeared, confirming that without signal peptide, htK made from klk1 expression cannot get out of the cells. In the culture medium of transfected cc-503 cells (sgk1, sgk2, sgk3), the band at 55 kDa still exists, indicating htK protein made from klk1 was successfully secreted into the medium from cells and the existence of htK protein in the medium was not due to cell lysate after cell death.
2.3. Gene Expression and Enzyme Activity under Heat Stress
The changes of the klk1 gene expression of the transfected cc-503 after heat stress are depicted in Figure 5. Sixty minutes after heat stimulation, the mRNA level reached a maximum, about three times higher than that before heat stimulation. Then, at 90 min, it drop to lower than level of 0 min and maintained that level from 90 min till 180 min.
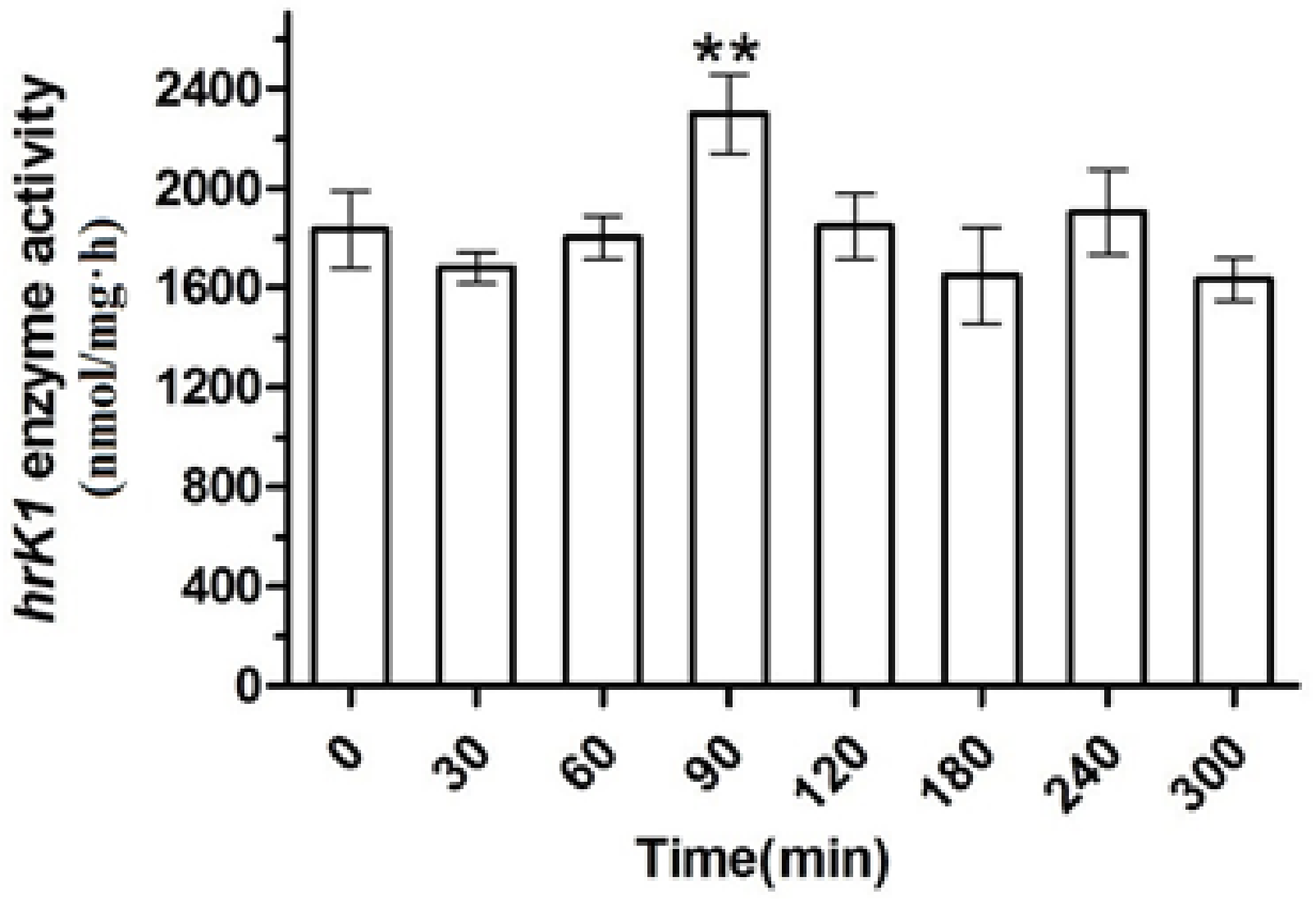
The enzyme activity analysis of the same samples (Figure 6) shows that the initial catalytic activity of htK (resulting from klk1 expression) was at 1800 nmol/mg·h. The activity decreased somewhat at 30 min possibly due to heat stress. It then reached a peak activity of 2400 nmol/mg·h at 90 min, possibly due to heat activated HSP70A, which in turn increased the mRNA expression and then the protein expression.
3. Discussion
C. reinhardtii belongs to green algae, the ancestor of land plants [20]. It exhibits a wide capacity in biosynthesis, which presents unique opportunities for commercial exploitation using bioengineering. Much research work was done to use C. reinhardtii as a bio-manufacturing platform to produce mammalian/human recombinant protein [1,3]. Most recombinant proteins, achieved by using C. reinhardtii’s gene expression systems, are accumulated intracellularly in algae cells [21]. The harvesting and purification of intracellular protein present a significant challenge. Therefore, more researchers are starting to focus on the secretory expression of recombinant proteins.
Rasala et al. recently achieved the expression and secretion of xylanase1 in C. reinhardtii by using a signal peptide [15]. In our study, we inserted a signal peptide gene (sg) upstream of the target exogenous gene and achieved the secretion of the recombinant protein htK. Therefore, we show that in principle it is possible to engineer C. reinhardtii to produce secretory human protein. Even though the production is still relatively low, we believe that through proper adjustment and bioengineering, we will be able to enhance the extracellular production of the target human recombinant protein. Interestingly, in Rasala and co-workers’ report, they found that extracellular and intracellular xylanase1, both produced by transgenic C. reinhardtii cells, had different migration speeds in the SDS-PAGE assay. The extracellular one moved slower than the intracellular one, even though they have very similar enzyme activities [15]. This phenomena suggests that the extracellular secreted protein may have gone through a post-translational modification or glycosylation. We also found similar a phenomenon which shows that the recombinant protein’s molecular weight was higher than the theoretical one, both intracellular and extracellular, even though the activities of the enzymes were similar. Further experiments are needed to fully elucidate the nature and process of this post-translational modification. In our design, we used endogenous signal peptides of C. reinhardtii, therefore, it should be easily recognizable by C. reinhardtii. In theory, linking this signal peptide with other exogenous genes to produce various secretory recombinant proteins should be feasible. We are in the process of testing this signal peptide on more exogenous recombinant proteins to verify this feasibility.
In 2000, Schroda et al. inserted HSP70A upstream of the RBCS2 promoter to get an enhanced gene transcription and protein synthesis through light–heat stimulation [22]. In their report, heat stress significantly enhanced arylsulfatase (ARS) gene expression, its mRNA level reached a maximum after 60 min, and its enzyme activity reached a maximum at 90–180 min, 1000 times higher than that without stimulation. We inserted HSP70A upstream of our target genes and have observed the enhancement of the transcription after heat stimulation as well, albeit at a much lower level. Under heat stress, the mRNA of klk1 was only enhanced three times (Figure 5) and enzyme activity only increased 33.3% (Figure 6). This may be due to the fact that klk1 is an exogenous gene to C. reinhardtii. Apart from inserting HSP70A and applying heat stress, other strategies to enhance production may also be considered, which include using priority codon and inserting regulation genes.
4. Materials and Methods
4.1. Construction of Plasmid
SignalP V3.0 (http://www.cbs.dtu.dk/services/SignalP/) was used to predict phosphate-repressible alkaline phosphatase (GenBank entry: XP_001703098.1) signal peptide and the signal peptide gene sg was synthesized accordingly. The vector used was pH124 (kept in our laboratory), which contains the HSP70A-RBCS2 promoter and the RBCS2 terminator [6]. The sg was inserted to obtain a secretory pHsg124 vector. Then, the human klk1 was inserted downstream of sg to obtain the pHsgk124 vector (see Figure 7).
The glass-bead method [6] was used to transfect HSP70A-RBCS2-sg-KLK1-RBCS2 into C. reinhardtii cc-503. Transfected C. reinhardtii cells were cultured in plates containing 10 µg/mL Zeocin.
4.2. Materials and Culture Conditions
The cell wall deficient strain cc-503 of C. reinhardtii was purchased from the Chlamydomonas Center at Duke University (Durham, NC, USA). K6-7 is a transgenic strain of C. reinhardtii developed by our laboratory previously, which does not have a signal peptide. A tris-acetate-phosphate medium was used to culture algae and the algal cells were under continuous irradiation for 24 h a day at 22 °C.
4.3. PCR Analysis
Based on known Chinese klk1 (GenBank entry: AY703451), primers were designed as the following:
Prsklk11: 5′-AAGATCTTCATGTGGTTCCTGGTTCTGTG-3′
Prsklk12: 5′-GCCACGTGACGCGTTCAGGAGTTCTCCGCTATGGT-3′
The PCR analysis for sequence confirmation was done by sending synthesized samples to Takara Biotechnology Inc. (Dalian, China) for sequencing. The gel running conditions were: 94 °C for 10 min, followed by 35 cycles of 94 °C for 60 s, 60 °C for 60 s, and 72 °C for 60 s, plus a final extension for 10 min.
4.4. Total DNA Southern Blot
Total DNA extraction was performed using DNeasy kit (Takara Biotechnology, Dalian, China) and following the Roche’s (Mannheim, Germany) manual and digoxin (DIG) high prime DNA labeling and detection starter kit I, as per the manufacturer’s instruction.
4.5. Fluorescence Quantitative PCR
C. reinhardtii was cultured under normal conditions (in a conical flask at 22 °C with a light intensity of 25 μmol.m−2·s−1) up to the logarithmic phase. Just before heat stimulation a 1 mL sample was taken. The culture was then moved to a 40 °C incubator, heat-stimulated for 30 min, and another 1 mL sample was taken. The culture was moved back to normal culture conditions and 1 mL repeated sampling was done every 30 min five times till reaching the 180 min mark. Each sample was immediately centrifuged at 13,800 g for 1 min at 4 °C. Then, the supernatant was removed and the remaining sample stored under −80 °C until analysis.
The total RNA extraction was done on those samples collected at the above mentioned seven time points. The RNA extraction was performed using the RNAfast200 kit (Fastagen, Shanghai, China). A quantitative fluorescence PCR machine (ABI 7300, ThermoFisher, Waltham, MA, USA) was used to measure the RNA. Each sample was diluted to 250 mg/mL before a reverse transcription using Takara RNA PCR kit (V. 3.0, Takara Biotechnology, Dalian, China) was performed according to the manufacturer’s instructions. The primers used for klk1 were:
Prklk1rt1: 5′-TGGGTGCTCACAGCTGCTCA-3′
Prklk1rt2: 5′-GTGTGGGAAGCTCTCACTGA-3′
The primers used for C. reinhardtii β-actin were:
Practin1: 5′-ACCCCGTGCTGCTGACTG-3′
Practin2: 5′-ACGTTGAAGGTCTCGAACA-3′
4.6. Protein Western Blot Analysis and Enzyme Activity Measurement
C. reinhardtii cell protein of 10 µg and 10 µg of super concentrated (50 times) cell culture medium were used for Western blot analysis. htK monoclonal antibody was used as the primary antibody and alkaline phosphatase-labeled sheep anti-mouse IgG was used as the secondary antibody (both purchased from Sigma-Aldrich, St Louis, MO, USA). The htK was reduced by thermolysin (4 units). Then, the htK enzyme activity was measured using a photometer, which measures the transformation of the substrate D-Val-Leu-Arg-PNA (S-2266, Chromogenix, Milano, Italy) by htK at 405 nm. Enzyme activity was standardized to the total protein of the respective sample. The protein concentrations were determined by using the Bradford assay.
4.7. Statistical Analysis
Each experiment was performed in triplicate and results were presented as mean ± SD. One-way ANOVA analysis was performed with Graphpad Prism 5. Significant differences were defined as p < 0.05 via Dunnett’s post-hoc test.
5. Conclusions
We designed a signal peptide and resulting secretory plasmid and incorporated it into C. reinhardtii. We successfully produced secretory human htK using transgenic C. reinhardtii. This technology can be used to produce large amounts of htK at a much lower cost than current bio-production strategy, and has significant medical and economic implications. This technology can also be used to produce other recombinant proteins that are important for medicine or industry.
Author Contributions
J.C., J.W. and Z.H. conceived and designed the experiments; J.C. and J.W performed the experiments; J.C., J.W. and Z.H. analyzed the data; Z.H. contributed reagents, materials and analysis tools; J.C., Q.W. and Z.H. wrote the paper.
Funding
This work was supported by the Chinese National key R & D plan for synthetic biology project, Shenzhen Grant Plan for Science & Technology (Shenzhen basic research projects, subject layout project) and the Grant Plan for the Demonstration Project of Marine Economic Development in Shenzhen to Zhangli Hu.
Acknowledgments
We would like to thank three anonymous reviewers for comments that improved this manuscript, and Yihong Zheng for revising the manuscript.
Conflicts of Interest
The authors declare no conflict of interest. The founding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
References
- Mayfield, S.P.; Manuell, A.L.; Chen, S.; Wu, J.; Tran, M.; Siefker, D.; Muto, M.; Marin-Navarro, J. Chlamydomonas reinhardtii chloroplasts as protein factories. Curr. Opin. Biotechnol. 2007, 18, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Rasala, B.A.; Mayfield, S.P. The microalga Chlamydomonas reinhardtii as a platform for the production of human protein therapeutics. Bioeng. Bugs 2011, 2, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Mu, F.Y.; Li, H.; Hu, Z.L. Expression of tandem repeat Cecropin B in Chlamydomonas reinhardtii and its antibacterial effect. Prog. Biochem. Biophys. 2012, 39, 344–351. [Google Scholar] [CrossRef]
- Wang, C.G.; Hu, Z.L.; Hu, W.; Lei, A.P. Expression and molecular analysis of phbB gene in Chlamydomonas reinhardtii. Chin. Sci. Bull. 2004, 49, 1713–1717. [Google Scholar]
- Zheng, K.J.; Wang, C.G.; Xiao, M.; Chen, J.; Li, J.C.; Hu, Z.L. Expression of bkt and bch genes from Haematococcus pluvialis in transgenic Chlamydomonas. Sci. China Life Sci. 2014, 57, 1028–1033. [Google Scholar] [CrossRef]
- Wang, C.G.; Hu, Z.L.; Lei, A.P. Biosynthesis of poly-3-hydroxybutyrate (phb) in the transgenic green alga Chlamydomonas reinhardtii. Phycol. Soc. Am. 2010, 46, 396–402. [Google Scholar]
- Wu, J.X.; Hu, Z.L.; Wang, C.G.; Li, S.F.; Lei, A.P. Efficient expression of green fluorescent protein (GFP) mediated by a chimeric promoter in Chlamydomonas reinhardtii. Chin. J. Oceanol. Limnol. 2008, 26, 242–247. [Google Scholar] [CrossRef]
- Surzycki, R.; Greenham, K.; Kitayama, K.; Dibal, F.; Wagner, R.; Rochaix, J.D.; Ajam, T.; Surzycki, S. Factors effecting expression of vaccines in microalgae. Biologicals 2009, 37, 133–138. [Google Scholar] [CrossRef]
- Hu, Z.L; Fan, Z.; Zhao, Z.L.; Chen, J.; Li, J.C. Stable expression of antibiotic-resistant gene ble from Streptoalloteichus hindustanus in the mitochondria of Chlamydomonas reinhardtii. PLoS ONE 2011, 7, e35542. [Google Scholar] [CrossRef]
- Hu, Z.L.; Zhao, Z.L.; Wu, Z.H.; Chen, J. Successful expression of heterologous egfp gene in the mitochondria of a photosynthetic eukaryote Chlamydomonas reinhardtii. Mitochondrion 2011, 5, 716–721. [Google Scholar] [CrossRef]
- Han, S.H.; Hu, Z.L.; Lei, A.P. Expression and function analysis of the metallothionein-like (MT-like) gene from Festuca rubra in Chlamydomonas reinhardtii chloroplast. Sci. China Ser. C 2008, 51, 1076–1081. [Google Scholar] [CrossRef] [PubMed]
- Von Gunnar, H. Life and death of a signal peptide. Nature 1998, 396, 111–112. [Google Scholar]
- Uchia, H.; Naito, N.; Asadaa, N.; Wadaa, M.; Ikedaa, M.; Kobayashia, H.; Asanagia, M.; Moria, K.; Fujitaa, Y.; Kondaa, K.; et al. Secretion of authentic 20-kDa human growth hormone (20 kDa hGH) in Escherichia Coli and properties of the purified product. J. Biotechnol. 1997, 55, 101–112. [Google Scholar] [CrossRef]
- Kjeldsen, T.; Pettensson, A.F.; Hach, M. Secretory expression and characterization of insulin in Pichia pastoris. J. Biotechnol. Appl. Biochem. 1999, 29, 79–86. [Google Scholar]
- Rasala, B.A.; Lee, P.A.; Shen, Z.X.; Briggs, S.P.; Mendez, M.; Mayfield, S.P. Robust expression and secretion of Xylanase1 in Chlamydomonas reinhardtii by fusion to a selection gene and processing with the FMDV 2A peptide. PLoS ONE 2012, 7, e43349. [Google Scholar] [CrossRef] [PubMed]
- Yousef, G.M.; Diamandis, E.P. The new human tissue kallikrein gene family: structure, function and association to disease. Endocr. Rev. 2001, 22, 184–204. [Google Scholar] [CrossRef] [PubMed]
- Borgono, C.A.; Michael, I.P.; Diamandis, E.P. Human tissue kallikreins: Physiologic roles and applications in cancer. Mol. Cancer Res. 2004, 2, 257–280. [Google Scholar] [PubMed]
- Bhoola, K.D.; Figueroa, C.D.; Worthy, K. Bioregulation of kinins: Kallikreins, kininogens, and kininases. Pharmacol. Rev. 1992, 44, 1–80. [Google Scholar]
- Clements, J. The molecular biology of the kallikreins and their roles in inflammation. In The Kinin System; Farmer, S., Ed.; Academic Press: New York, NY, USA, 1997; pp. 71–97. [Google Scholar]
- Scaife, M.A.; Nguyen, G.T.D.T.; Rico, J.; Lambert, D.; Helliwell, K.E.; Smith, A.G. Establishing Chlamydomonas reinhardtii as an industrial biotechnology host. Plant J. 2015, 82, 532–546. [Google Scholar] [CrossRef]
- Specht, E.; Miyake-Stoner, S.; Mayfield, S. Micro-algae come of age as a platform for recombinant protein production. Biotechnol. Lett. 2010, 32, 1373–1383. [Google Scholar] [CrossRef] [Green Version]
- Schroda, M.; Blcker, D.; Beck, C.F. The HSP70A promoter as a tool for the improved expression of transgenes in Chlamydomonas. Plant J. 2000, 21, 121–131. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
PCR-electrophoresis result for the sgk genome (klk1 genome with upstream signal peptide genome) detection. Marker, DL-2000; 1, non-transfected cc-503; 2, sgk-1; 3, sgk-2; 4, sgk-3; 5, negative control (water).
Figure 1.
PCR-electrophoresis result for the sgk genome (klk1 genome with upstream signal peptide genome) detection. Marker, DL-2000; 1, non-transfected cc-503; 2, sgk-1; 3, sgk-2; 4, sgk-3; 5, negative control (water).

Figure 2.
Southern blot analysis of transgenic algae with the sgk genome (using DIG high prime DNA labeling and detection starter kit I, see Section 4.4). M, marker; 1, non-transfected cc-503 DNA fragment; 2, sgk-1 DNA fragment; 3, sgk-2 DNA fragment; 4, sgk-3 DNA fragment.
Figure 2.
Southern blot analysis of transgenic algae with the sgk genome (using DIG high prime DNA labeling and detection starter kit I, see Section 4.4). M, marker; 1, non-transfected cc-503 DNA fragment; 2, sgk-1 DNA fragment; 3, sgk-2 DNA fragment; 4, sgk-3 DNA fragment.

Figure 3.
Western blot analysis of intracellular protein produced by transgenic and wild type algae. Marker, Fermantas PageRulerTM (Glen Burnie, MD, USA) pre-stained protein ladder; 1, sgk-1; 2, sgk-2; 3, sgk-3; 4, k6-7; 5, cc-503.
Figure 3.
Western blot analysis of intracellular protein produced by transgenic and wild type algae. Marker, Fermantas PageRulerTM (Glen Burnie, MD, USA) pre-stained protein ladder; 1, sgk-1; 2, sgk-2; 3, sgk-3; 4, k6-7; 5, cc-503.

Figure 4.
Western blot analysis of extracellular protein produced by transgenic and wild type algae. Marker, Fermantas PageRulerTM (Glen Burnie, MD, USA) pre-stained protein ladder; 1, sgk-1; 2, sgk-2; 3, sgk-3; 4, k6-7; 5, cc-503.
Figure 4.
Western blot analysis of extracellular protein produced by transgenic and wild type algae. Marker, Fermantas PageRulerTM (Glen Burnie, MD, USA) pre-stained protein ladder; 1, sgk-1; 2, sgk-2; 3, sgk-3; 4, k6-7; 5, cc-503.

Figure 5.
mRNA expression level of klk1 in transgenic C. reinhardtii (sgk) cells under heat shock and intense light conditions. The relative transcription levels of klk1 were determined after 0, 30, 60, 90, 120, 150, and 180 min by qRTPCR. β-actin was used as a reference gene and the values were normalized to the transcript levels in the control. Data are averages of biological triplicate, and the error bars represent standard deviation. One asterisk, p < 0.05; two asterisks, p < 0.01; three asterisks, p < 0.001.
Figure 5.
mRNA expression level of klk1 in transgenic C. reinhardtii (sgk) cells under heat shock and intense light conditions. The relative transcription levels of klk1 were determined after 0, 30, 60, 90, 120, 150, and 180 min by qRTPCR. β-actin was used as a reference gene and the values were normalized to the transcript levels in the control. Data are averages of biological triplicate, and the error bars represent standard deviation. One asterisk, p < 0.05; two asterisks, p < 0.01; three asterisks, p < 0.001.

Figure 6.
Enzyme activity changes of hrK in transgenic C. reinhardtii (sgk) cells under heat shock and intense light conditions. Significant difference versus 0 min time point sample was indicated with one asterisk (p < 0.05) and two asterisks (p < 0.01). Data were reported as mean ± SD of three independent repeated experiments.
Figure 6.
Enzyme activity changes of hrK in transgenic C. reinhardtii (sgk) cells under heat shock and intense light conditions. Significant difference versus 0 min time point sample was indicated with one asterisk (p < 0.05) and two asterisks (p < 0.01). Data were reported as mean ± SD of three independent repeated experiments.

Figure 7.
Construction of pHSgk124 plasmid. Abbreviations: sg, signal peptide; klk1, human tissue kallikrein 1 gene; ble, bleomycin resistance gene; PmaC I, enzyme cleavage point 1; Bgl II, enzyme cleavage point 2.
Figure 7.
Construction of pHSgk124 plasmid. Abbreviations: sg, signal peptide; klk1, human tissue kallikrein 1 gene; ble, bleomycin resistance gene; PmaC I, enzyme cleavage point 1; Bgl II, enzyme cleavage point 2.

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chen, J.; Wu, J.; Wu, Q.; Hu, Z. Biosynthesis and Secretion of Human Tissue Kallikrein in Transgenic Chlamydomonas reinhardtii. Mar. Drugs 2018, 16, 493. https://doi.org/10.3390/md16120493
AMA Style
Chen J, Wu J, Wu Q, Hu Z. Biosynthesis and Secretion of Human Tissue Kallikrein in Transgenic Chlamydomonas reinhardtii. Marine Drugs. 2018; 16(12):493. https://doi.org/10.3390/md16120493
Chicago/Turabian StyleChen, Jun, Jinxia Wu, Qingyu Wu, and Zhangli Hu. 2018. "Biosynthesis and Secretion of Human Tissue Kallikrein in Transgenic Chlamydomonas reinhardtii" Marine Drugs 16, no. 12: 493. https://doi.org/10.3390/md16120493
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.