Water-Soluble Fish Protein Intake Led to Lower Serum and Liver Cholesterol Concentrations in Obese Zucker fa/fa Rats

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Dietary Compositions
2.2. Growth and Energy Intake
2.3. Cholesterol and Bile Acids in Serum
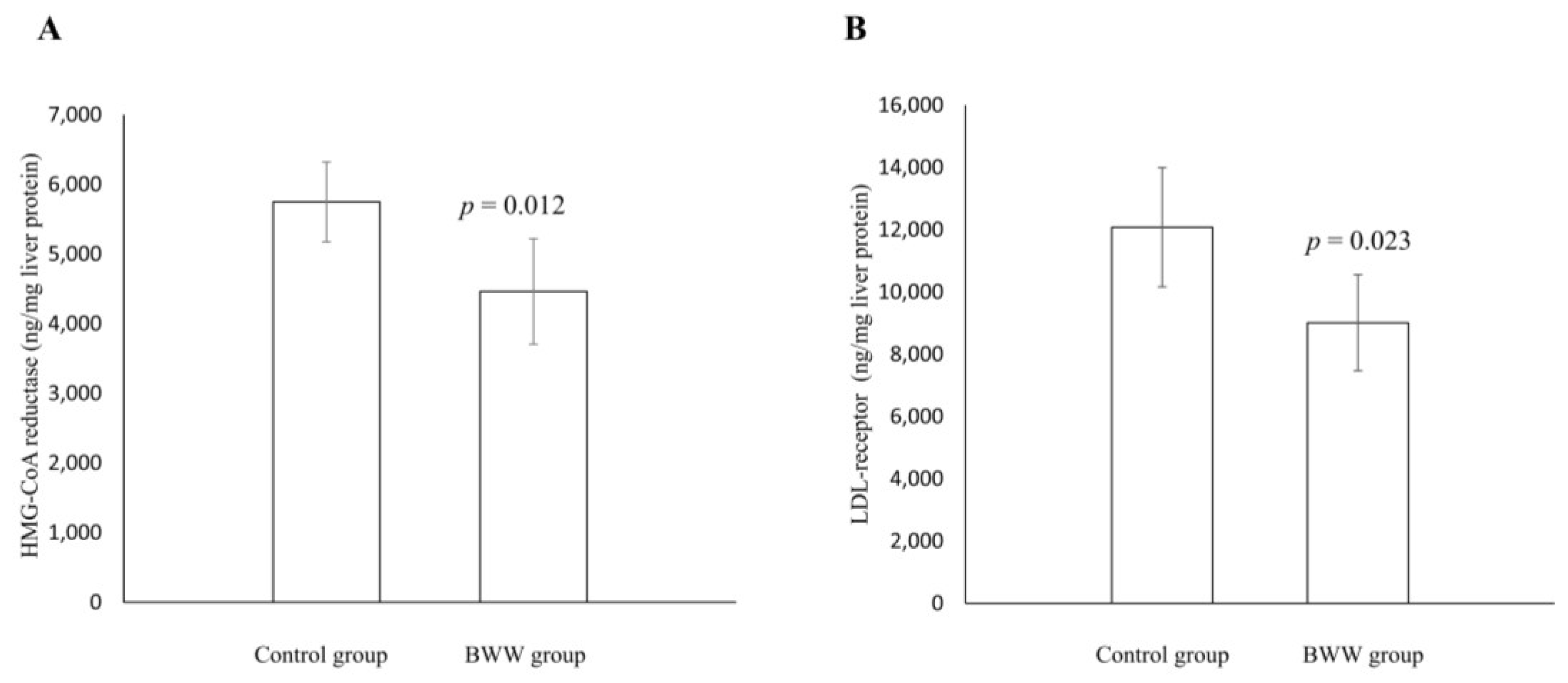
2.4. Cholesterol, HMG-CoA Reductase, and LDL Receptor Concentrations in Liver and Fecal Cholesterol and Bile Acids
3. Discussion
4. Materials and Methods
4.1. Ethical Statement
4.2. Animals and Diets
4.3. Preparation of Blue Whiting Water-Soluble Protein
4.4. Design
4.5. Analyses of Diets
4.6. Serum Analyses
4.7. Cholesterol in Liver and Feces
4.8. Total Bile Acids in Feces
4.9. Liver Concentrations of 3-Hydroxy-3-Methylglutaryl Coenzyme A (HMG-CoA) Reductase, LDL Receptor, and Protein
4.10. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, H.; Naghavi, M.; Allen, C.; Barber, R.M.; Bhutta, Z.A.; Carter, A.; Casey, D.C.; Charlson, F.J.; Chen, A.Z.; Coates, M.M.; et al. Global, regional, and national life expectancy, all-cause mortality, and cause-specific mortality for 249 causes of death, 1980–2015: A systematic analysis for the global burden of disease study. Lancet 2015, 388, 1459–1544. [Google Scholar] [CrossRef] [Green Version]
- Danaei, G.; Lu, Y.; Singh, G.; Stevens, G.; Cowan, M.; Farzadfar, F.; Lin, J.; Finucane, M.; Rao, M.; Khang, Y.; et al. Cardiovascular disease, chronic kidney disease, and diabetes mortality burden of cardiometabolic risk factors from 1980 to 2010: A comparative risk assessment. Lancet Diabetes Endocrinol. 2014, 2, 634–647. [Google Scholar]
- World Health Organization. Prevention of cardiovascular disease. In Guideline for Assessment and Management of Cardiovascular Risk; WHO Press: Geneva, Switzerland, 2007. [Google Scholar]
- American Heart Association. Fish 101. Available online: http://www.heart.org/HEARTORG/HealthyLiving/HealthyEating/Fish-101_UCM_305986_Article.jsp#aha_recommendation (accessed on 27 January 2017).
- National Health Service. Eating a Balanced Diet. Available online: http://www.nhs.uk/Livewell/Goodfood/Pages/Healthyeating.aspx (accessed on 27 January 2017).
- Virtanen, J.K.; Mozaffarian, D.; Chiuve, S.E.; Rimm, E.B. Fish consumption and risk of major chronic disease in men. Am. J. Clin. Nutr. 2008, 88, 1618–1625. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Huang, T.; Yu, Y.; Hu, X.; Yang, B.; Li, D. Fish consumption and chd mortality: An updated meta-analysis of seventeen cohort studies. Public Health Nutr. 2012, 15, 725–737. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Wu, J.H. Omega-3 fatty acids and cardiovascular disease: Effects on risk factors, molecular pathways, and clinical events. J. Am. Coll. Cardiol. 2011, 58, 2047–2067. [Google Scholar] [CrossRef] [PubMed]
- Vikoren, L.A.; Nygard, O.K.; Lied, E.; Rostrup, E.; Gudbrandsen, O.A. A randomised study on the effects of fish protein supplement on glucose tolerance, lipids and body composition in overweight adults. Br. J. Nutr. 2013, 109, 648–657. [Google Scholar] [CrossRef] [PubMed]
- Hosomi, R.; Fukunaga, K.; Arai, H.; Kanda, S.; Nishiyama, T.; Yoshida, M. Fish protein decreases serum cholesterol in rats by inhibition of cholesterol and bile acid absorption. J. Food Sci. 2011, 76, H116–H121. [Google Scholar] [CrossRef] [PubMed]
- Hosomi, R.; Fukunaga, K.; Arai, H.; Nishiyama, T.; Yoshida, M. Effects of dietary fish protein on serum and liver lipid concentrations in rats and the expression of hepatic genes involved in lipid metabolism. J. Agric. Food Chem. 2009, 57, 9256–9262. [Google Scholar] [CrossRef] [PubMed]
- Wergedahl, H.; Liaset, B.; Gudbrandsen, O.A.; Lied, E.; Espe, M.; Muna, Z.; Mork, S.; Berge, R.K. Fish protein hydrolysate reduces plasma total cholesterol, increases the proportion of hdl cholesterol, and lowers acyl-coa:Cholesterol acyltransferase activity in liver of zucker rats. J. Nutr. 2004, 134, 1320–1327. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Beynen, A.C. Influence of dietary fish proteins on plasma and liver cholesterol concentrations in rats. Br. J. Nutr. 1993, 69, 767–777. [Google Scholar] [CrossRef] [PubMed]
- Iritani, N.; Narita, R.; Fujita, T.; Tanaka, T. Effects of dietary fish protein, soybean protein and casein on cholesterol turnover in rats. J. Nutr. Sci. Vitaminol. 1985, 31, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Drotningsvik, A.; Mjos, S.A.; Pampanin, D.M.; Slizyte, R.; Carvajal, A.; Remman, T.; Hogoy, I.; Gudbrandsen, O.A. Dietary fish protein hydrolysates containing bioactive motifs affect serum and adipose tissue fatty acid compositions, serum lipids, postprandial glucose regulation and growth in obese zucker fa/fa rats. Br. J. Nutr. 2016, 116, 1336–1345. [Google Scholar] [CrossRef] [PubMed]
- Shukla, A.; Bettzieche, A.; Hirche, F.; Brandsch, C.; Stangl, G.I.; Eder, K. Dietary fish protein alters blood lipid concentrations and hepatic genes involved in cholesterol homeostasis in the rat model. Br. J. Nutr. 2006, 96, 674–682. [Google Scholar] [PubMed]
- Benaicheta, N.; Labbaci, F.Z.; Bouchenak, M.; Boukortt, F.O. Effect of sardine proteins on hyperglycaemia, hyperlipidaemia and lecithin:Cholesterol acyltransferase activity, in high-fat diet-induced type 2 diabetic rats. Br. J. Nutr. 2016, 115, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Geirsdottir, M.; Sigurgisladottir, S.; Hamaguchi, P.Y.; Thorkelsson, G.; Johannsson, R.; Kristinsson, H.G.; Kristjansson, M.M. Enzymatic hydrolysis of blue whiting (micromesistius poutassou); functional and bioactive properties. J. Food Sci. 2011, 76, C14–C20. [Google Scholar] [CrossRef] [PubMed]
- Oterhals, A.; Samuelsen, T.A. Plasticization effect of solubles in fishmeal. Food Res. Int. 2015, 69, 313–321. [Google Scholar] [CrossRef]
- Kousoulaki, K.; Albrektsen, S.; Langmyhr, E.; Olsen, H.J.; Campbell, P.; Aksnes, A. The water soluble fraction in fish meal (stickwater) stimulates growth in atlantic salmon (Salmo salar L.) given high plant protein diets. Aquaculture 2009, 289, 74–83. [Google Scholar] [CrossRef]
- Kousoulaki, K.; Olsen, H.J.; Albrektsen, S.; Langmyhr, E.; Mjos, S.A.; Campbell, P.; Aksnes, A. High growth rates in atlantic salmon (Salmo salar L.) fed 7.5% fish meal in the diet. Micro-, ultra- and nano-filtration of stickwater and effects of different fractions and compounds on pellet quality and fish performance. Aquaculture 2012, 338, 134–146. [Google Scholar] [CrossRef]
- Erdmann, K.; Cheung, B.W.; Schroder, H. The possible roles of food-derived bioactive peptides in reducing the risk of cardiovascular disease. J. Nutr. Biochem. 2008, 19, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Aleixandre de Artinano, A.; Miguel Castro, M. Experimental rat models to study the metabolic syndrome. Br. J. Nutr. 2009, 102, 1246–1253. [Google Scholar] [CrossRef] [PubMed]
- Phillips, M.S.; Liu, Q.; Hammond, H.A.; Dugan, V.; Hey, P.J.; Caskey, C.J.; Hess, J.F. Leptin receptor missense mutation in the fatty zucker rat. Nat. Genet. 1996, 13, 18–19. [Google Scholar] [CrossRef] [PubMed]
- Zucker, L.M.; Zucker, T.F. Fatty, a new mutation in the rat. J. Hered. 1961, 52, 275–278. [Google Scholar] [CrossRef]
- Bray, G.A. The zucker-fatty rat: A review. Fed. Proc. 1977, 36, 148–153. [Google Scholar] [PubMed]
- Drotningsvik, A.; Mjos, S.A.; Hogoy, I.; Remman, T.; Gudbrandsen, O.A. A low dietary intake of cod protein is sufficient to increase growth, improve serum and tissue fatty acid compositions, and lower serum postprandial glucose and fasting non-esterified fatty acid concentrations in obese zucker fa/fa rats. Eur. J. Nutr. 2015, 54, 1151–1160. [Google Scholar] [CrossRef] [PubMed]
- Vik, R.; Tillander, V.; Skorve, J.; Vihervaara, T.; Ekroos, K.; Alexson, S.E.; Berge, R.K.; Bjorndal, B. Three differently generated salmon protein hydrolysates reveal opposite effects on hepatic lipid metabolism in mice fed a high-fat diet. Food Chem. 2015, 183, 101–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingelsson, E.; Schaefer, E.J.; Contois, J.H.; McNamara, J.R.; Sullivan, L.; Keyes, M.J.; Pencina, M.J.; Schoonmaker, C.; Wilson, P.W.; D’Agostino, R.B.; et al. Clinical utility of different lipid measures for prediction of coronary heart disease in men and women. JAMA 2007, 298, 776–785. [Google Scholar] [CrossRef] [PubMed]
- Feingold, K.R.; Grunfeld, C. Cholesterol lowering drugs. In Endotext; De Groot, L.J., Chrousos, G., Dungan, K., Feingold, K.R., Grossman, A., Hershman, J.M., Koch, C., Korbonits, M., McLachlan, R., New, M., et al., Eds.; MDTEXT.COM: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Soares, R.A.; Mendonca, S.; de Castro, L.I.; Menezes, A.C.; Areas, J.A. Major peptides from amaranth (amaranthus cruentus) protein inhibit HMG-CoA reductase activity. Int. J. Mol. Sci. 2015, 16, 4150–4160. [Google Scholar] [CrossRef] [PubMed]
- Rigotti, A.; Miettinen, H.E.; Krieger, M. The role of the high-density lipoprotein receptor SR-BI in the lipid metabolism of endocrine and other tissues. Endocr. Rev. 2003, 24, 357–387. [Google Scholar] [CrossRef] [PubMed]
- Horton, J.D.; Goldstein, J.L.; Brown, M.S. Srebps: Activators of the complete program of cholesterol and fatty acid synthesis in the liver. J. Clin. Investig. 2002, 109, 1125–1131. [Google Scholar] [CrossRef] [PubMed]
- Bremer, J. The conjugation of glycine with cholic acid and benzoic acid in rat liver homogenate. Bile acids and steroids. Acta Chem. Scand. 1955, 9, 2. [Google Scholar] [CrossRef]
- Chen, W.; Guo, J.X.; Chang, P. The effect of taurine on cholesterol metabolism. Mol. Nutr. Food Res. 2012, 56, 681–690. [Google Scholar] [CrossRef] [PubMed]
- Bergen, W.G.; Mersmann, H.J. Comparative aspects of lipid metabolism: Impact on contemporary research and use of animal models. J. Nutr. 2005, 135, 2499–2502. [Google Scholar] [CrossRef] [PubMed]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. Ain-93 purified diets for laboratory rodents: Final report of the american institute of nutrition ad hoc writing committee on the reformulation of the AIN-76a rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef] [PubMed]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Pampanin, D.M.; Larssen, E.; Provan, F.; Sivertsvik, M.; Ruoff, P.; Sydnes, M.O. Detection of small bioactive peptides from atlantic herring (Clupea harengus L.). Peptides 2012, 34, 423–426. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, S.; Futamura, Y.; Miwa, K.; Awano, T.; Yamauchi, K.; Kanamaru, Y.; Tadashi, K.; Kuwata, T. Identification of novel hypocholesterolemic peptides derived from bovine milk beta-lactoglobulin. Biochem. Biophys. Res. Commun. 2001, 281, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, R.; Ohinata, K.; Yoshikawa, M. Beta-lactotensin and neurotensin rapidly reduce serum cholesterol via NT2 receptor. Peptides 2003, 24, 1955–1961. [Google Scholar] [CrossRef] [PubMed]
- Lammi, C.; Zanoni, C.; Arnoldi, A. Iavpgeva, iavptgva, and lpyp, three peptides from soy glycinin, modulate cholesterol metabolism in HepG2 cells through the activation of the LDLR-SREBP2 pathway. J. Funct. Foods 2015, 14, 469–478. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Fujita, H.; Matoba, N.; Takenaka, Y.; Yamamoto, T.; Yamauchi, R.; Tsuruki, H.; Takahata, K. Bioactive peptides derived from food proteins preventing lifestyle-related diseases. Biofactors 2000, 12, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Myasoedov, N.F.; Lyapina, L.A.; Grigorjeva, M.E.; Obergan, T.Y.; Shubina, T.A.; Andreeva, L.A. Mechanisms for glyproline protection in hypercholesterolemia. Pathophysiology 2016, 23, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, S.; Nakamura, A.; Shibata, H.; Kanamaru, Y. Soystatin (VAWWMY), a novel bile acid-binding peptide, decreased micellar solubility and inhibited cholesterol absorption in rats. Biosci. Biotechnol. Biochem. 2010, 74, 1738–1741. [Google Scholar] [CrossRef] [PubMed]
- Takenaka, Y.; Nakamura, F.; Yamamoto, T.; Yoshikawa, M. Enterostatin (vpdpr) and its peptide fragment dpr reduce serum cholesterol levels after oral administration in mice. Biosci. Biotechnol. Biochem. 2003, 67, 1620–1622. [Google Scholar] [CrossRef] [PubMed]
- Suckling, K.E.; Benson, G.M.; Bond, B.; Gee, A.; Glen, A.; Haynes, C.; Jackson, B. Cholesterol lowering and bile acid excretion in the hamster with cholestyramine treatment. Atherosclerosis 1991, 89, 183–190. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]


{kind=link}
| Control Diet | BWW Diet | |
|---|---|---|
| Amino acids (g/kg diet) | ||
| Arginine | 6.9 | 7.2 |
| Glycine | 3.7 | 10.0 |
| Histidine | 5.6 | 4.5 |
| Isoleucine | 10.2 | 8.0 |
| Leucine | 18.3 | 15.0 |
| Lysine | 16.4 | 15.0 |
| Methionine | 6.9 | 6.5 |
| Phenylalanine | 10.1 | 8.2 |
| Threonine | 8.5 | 6.6 |
| Valine | 13.0 | 11.0 |
| Taurine | ND | 0.2 |
| Lysine/Arginine | 2.4 | 2.1 |
| Methionine/Glycine | 1.9 | 0.7 |
| Fatty acids 1 (g/kg diet) | ||
| 16:0 | 6.7 | 6.7 |
| 18:0 | 2.3 | 2.3 |
| 18:1n-9 | 12.4 | 12.6 |
| 18:1n-7 | 0.8 | 0.8 |
| 18:2n-6 | 29.0 | 29.8 |
| 18:3n-3 | 3.4 | 3.5 |
| 20:5n-3 | ND | 0.01 |
| 22:5n-3 | ND | ND |
| 22:6n-3 | ND | 0.02 |
| Parameters | Control Group | BWW Group | p-Value |
|---|---|---|---|
| Body weight at baseline (g) | 318 ± 8 | 303 ± 9 | 0.021 |
| Body weight at time of euthanasia (g) | 548 ± 35 | 510 ± 29 | 0.077 |
| Growth (% from baseline to endpoint) | 72 ± 8 | 68 ± 7 | 0.37 |
| Body weight to square body length ratio (kg/m2) | 10.0 ± 0.6 | 9.5 ± 0.1 | 0.10 |
| Relative liver weight (g/kg body weight) | 36.5 ± 5.6 | 32.2 ± 4.8 | 0.20 |
| Energy intake (kJ/24 h) | 456 ± 62 | 493 ± 27 | 0.21 |
| Biochemical Parameters | Control Group | BWW Group | p-Value |
|---|---|---|---|
| Total cholesterol (mmol/L) | 5.9 ± 1.0 | 4.6 ± 0.9 | 0.039 |
| Cholesteryl ester (mmol/L) | 4.6 ± 0.8 | 3.3 ± 0.7 | 0.017 |
| LDL cholesterol (mmol/L) | 1.5 ± 0.5 | 0.8 ± 0.3 | 0.0062 |
| HDL cholesterol (mmol/L) | 5.6 ± 0.7 | 4.1 ± 0.9 | 0.014 |
| Total bile acids (μmol/L) | 16.6 ± 11.8 | 16.7 ± 9.8 | 0.99 |
| Parameters | Control Group | BWW Group | p-Value |
|---|---|---|---|
| Liver cholesterol (µmol/g) | 9.4 ± 1.9 | 6.1 ± 1.6 | 0.015 |
| Fecal cholesterol (µmol/24 h) | 20.8 ± 7.0 | 16.8 ± 1.7 | 0.27 |
| Fecal bile acids (µmol/24 h) | 5.8 ± 2.6 | 4.5 ± 1.7 | 0.36 |
| Contents (g/kg Diet) | Control Diet | BWW Diet |
|---|---|---|
| Casein 1 | 216.00 | 144.00 |
| Blue Whiting Water-Soluble Protein 2 | - | 108.30 |
| Cornstarch | 511.67 | 475.50 |
| Sucrose | 90.00 | 90.00 |
| Cellulose | 50.00 | 50.00 |
| Soybean Oil | 70.00 | 70.00 |
| t-Butylhydroquinone | 0.015 | 0.015 |
| Mineral Mix (AIN-93-MX) | 35.00 | 35.00 |
| Vitamin Mix (AIN-93-VX) | 10.00 | 10.00 |
| l-Methionine | 1.60 | 1.60 |
| l-Cystine | 3.00 | 3.00 |
| Choline Bitartrate 3 | 2.50 | 2.50 |
| Growth and maintenance supplement (#410751) 4 | 10.00 | 10.00 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drotningsvik, A.; Vikøren, L.A.; Mjøs, S.A.; Oterhals, Å.; Pampanin, D.; Flesland, O.; Gudbrandsen, O.A. Water-Soluble Fish Protein Intake Led to Lower Serum and Liver Cholesterol Concentrations in Obese Zucker fa/fa Rats. Mar. Drugs 2018, 16, 149. https://doi.org/10.3390/md16050149
Drotningsvik A, Vikøren LA, Mjøs SA, Oterhals Å, Pampanin D, Flesland O, Gudbrandsen OA. Water-Soluble Fish Protein Intake Led to Lower Serum and Liver Cholesterol Concentrations in Obese Zucker fa/fa Rats. Marine Drugs. 2018; 16(5):149. https://doi.org/10.3390/md16050149
Chicago/Turabian StyleDrotningsvik, Aslaug, Linn Anja Vikøren, Svein Are Mjøs, Åge Oterhals, Daniela Pampanin, Ola Flesland, and Oddrun Anita Gudbrandsen. 2018. "Water-Soluble Fish Protein Intake Led to Lower Serum and Liver Cholesterol Concentrations in Obese Zucker fa/fa Rats" Marine Drugs 16, no. 5: 149. https://doi.org/10.3390/md16050149