Metabolite Profiling of the Microalgal Diatom Chaetoceros Calcitrans and Correlation with Antioxidant and Nitric Oxide Inhibitory Activities via 1H NMR-Based Metabolomics


Abstract
:1. Introduction
2. Results and Discussion
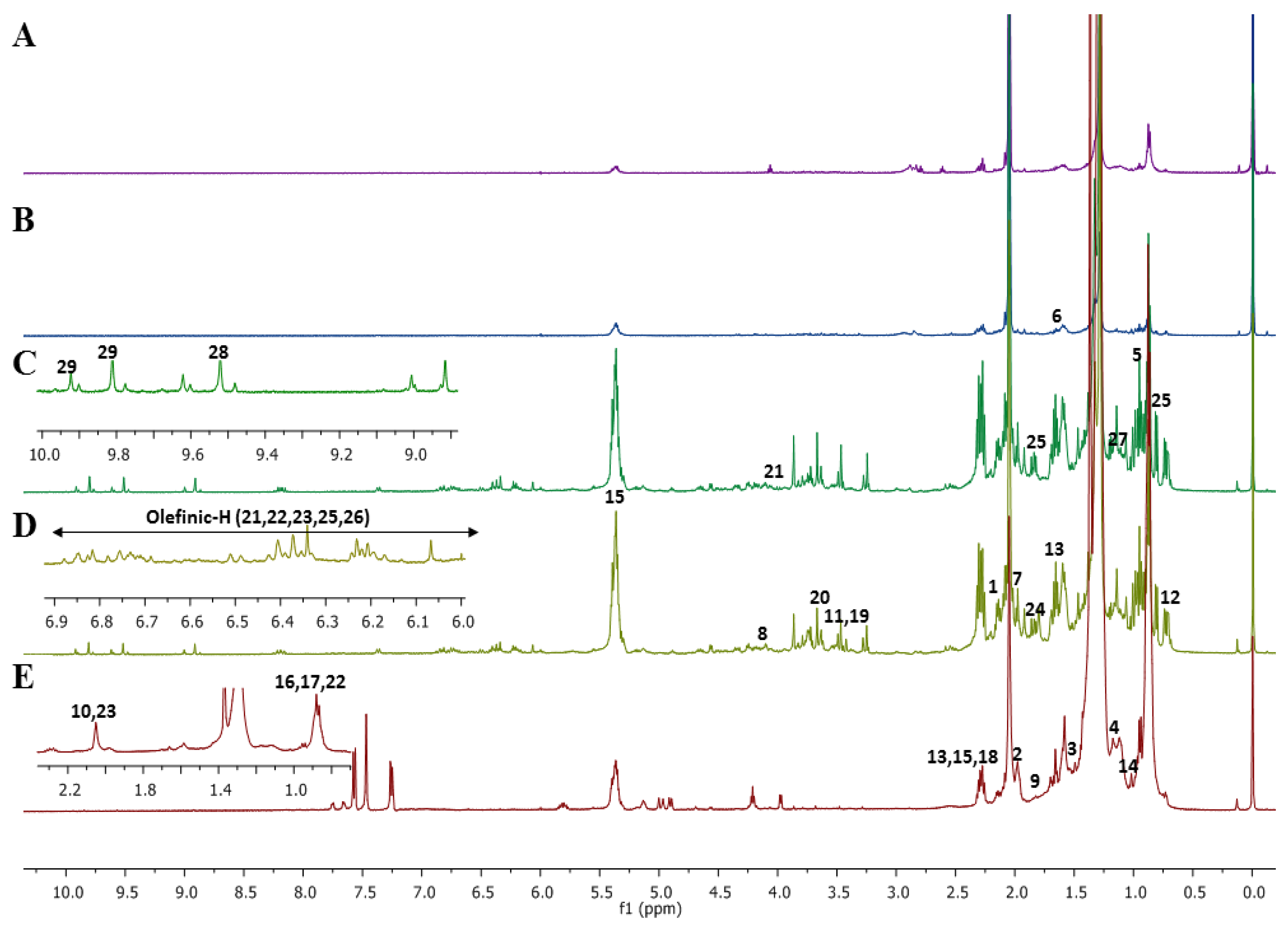
2.1. Assignments of Metabolites by 1D Nuclear Magnetic Resonance (NMR) and 2D NMR Spectra in Microalgal Crude Extracts
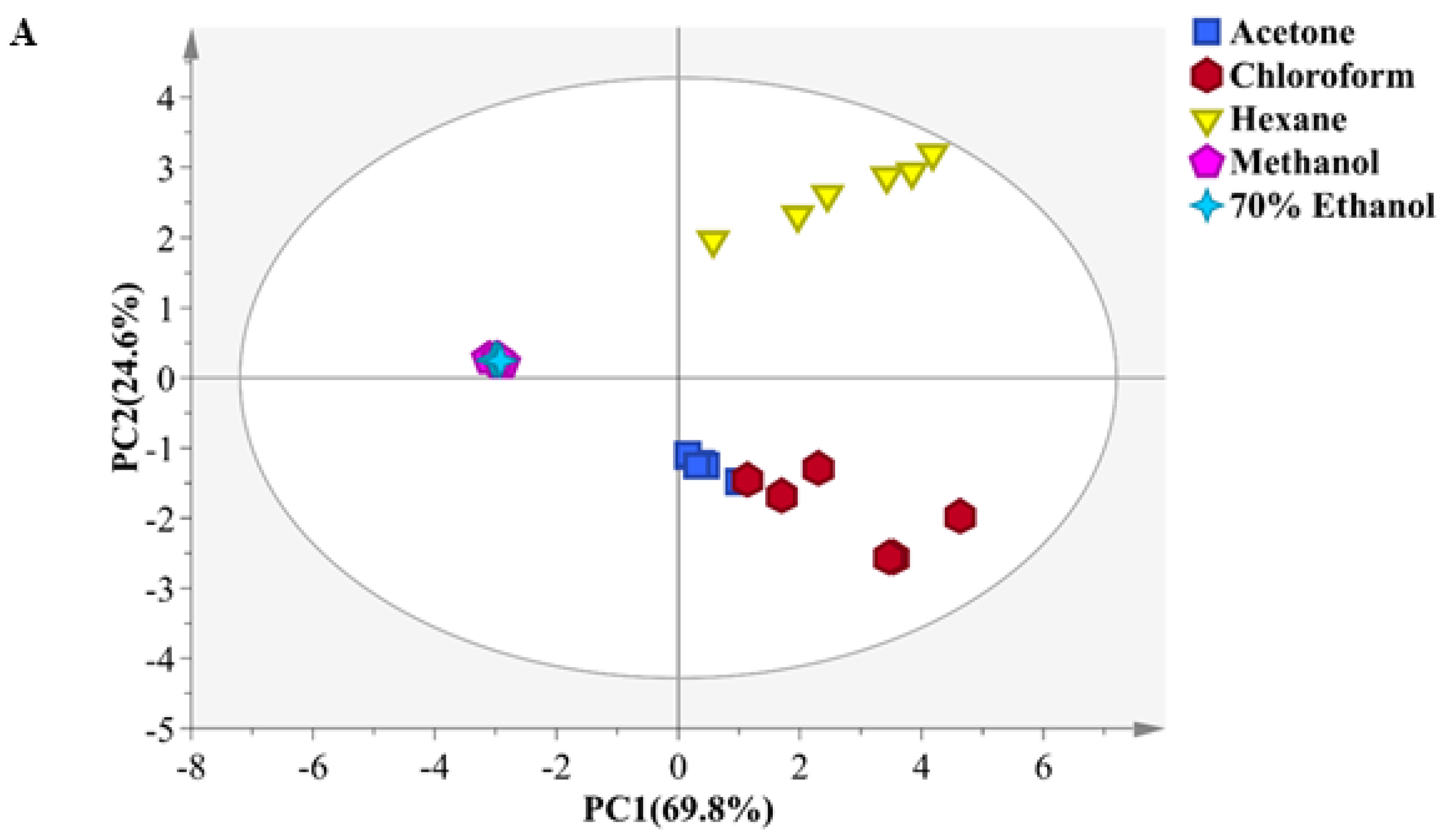
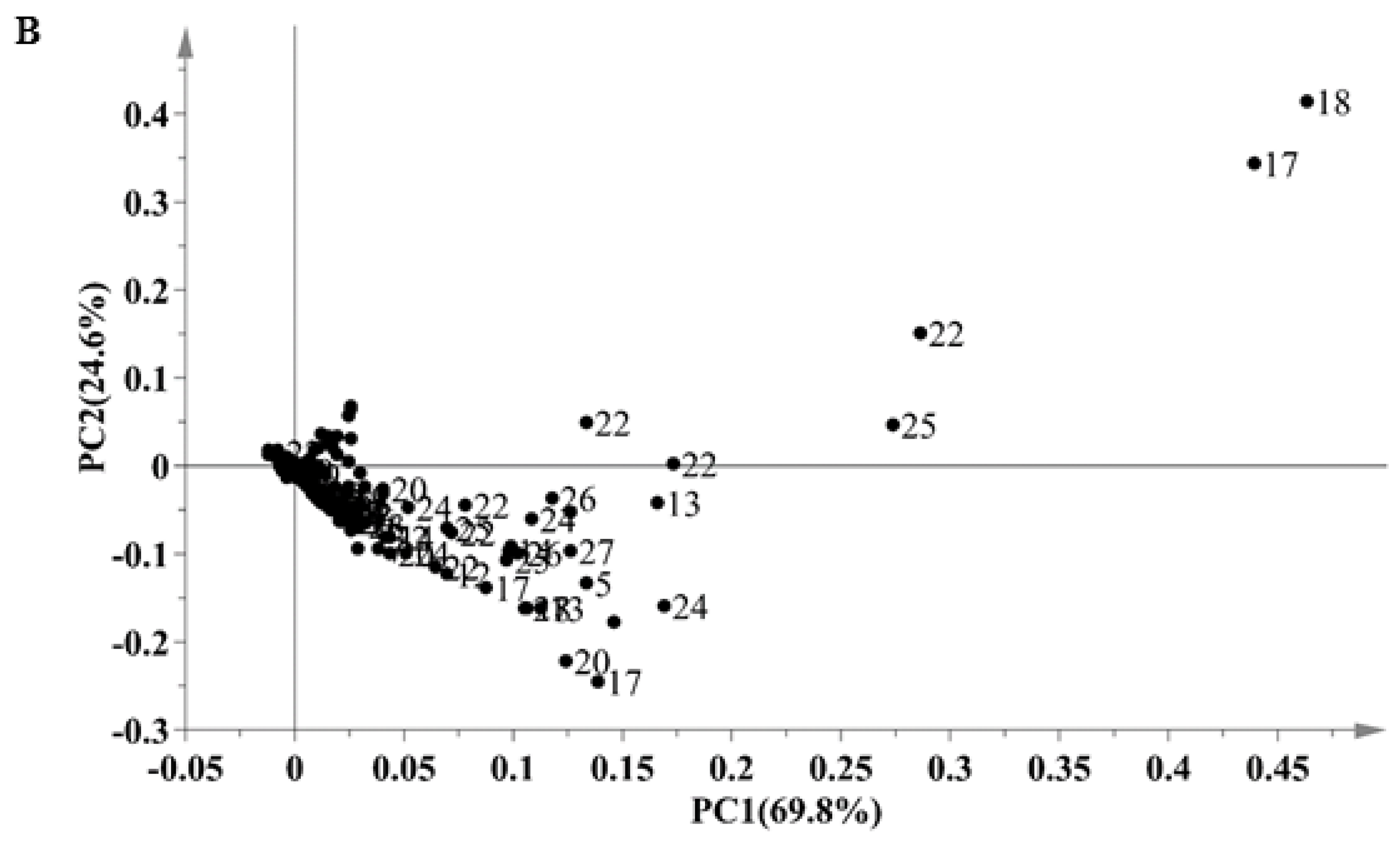
2.2. Classification of Different Solvent Extracts by Principal Component Analysis (PCA)
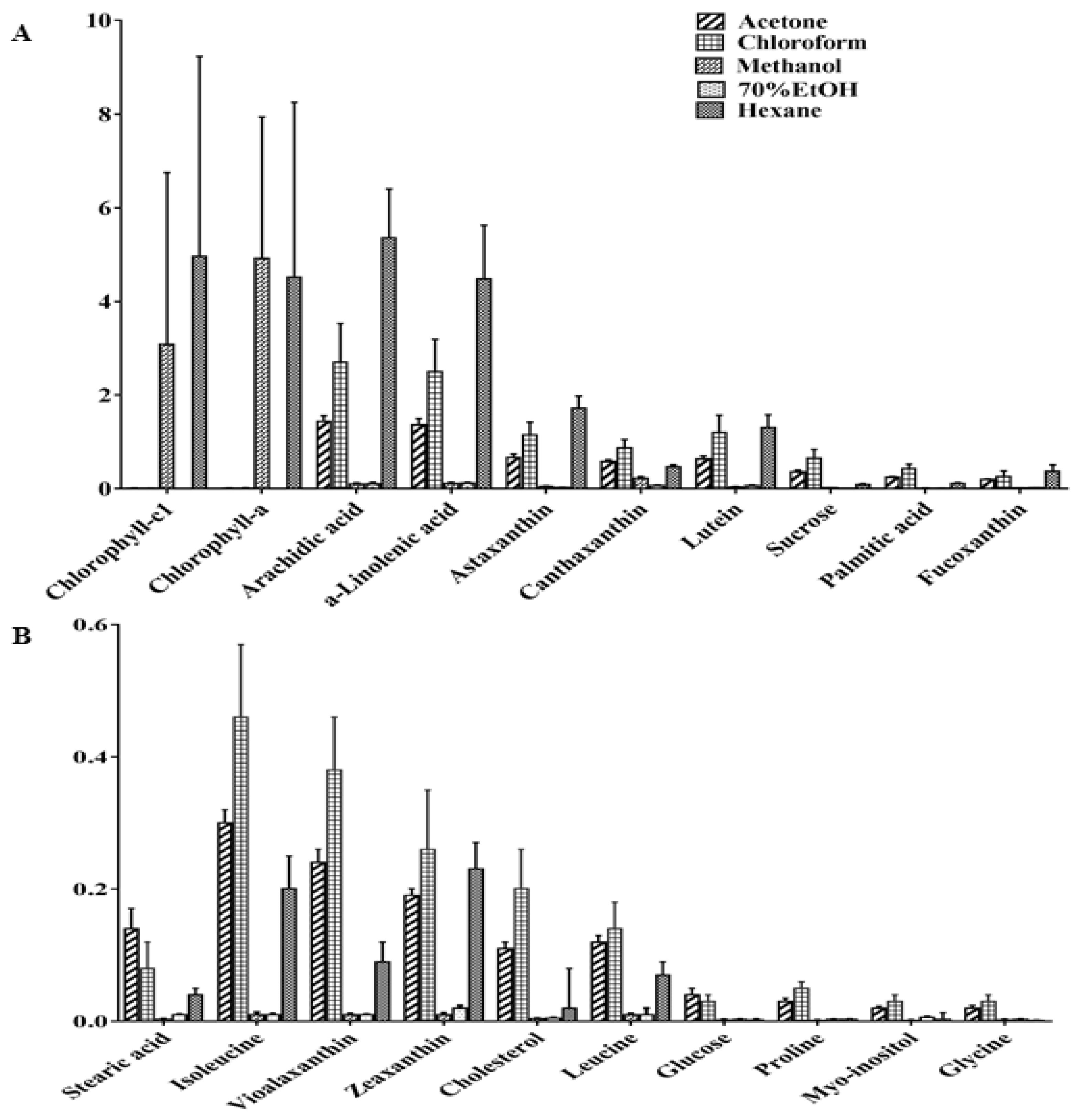
2.3. Relative Quantification
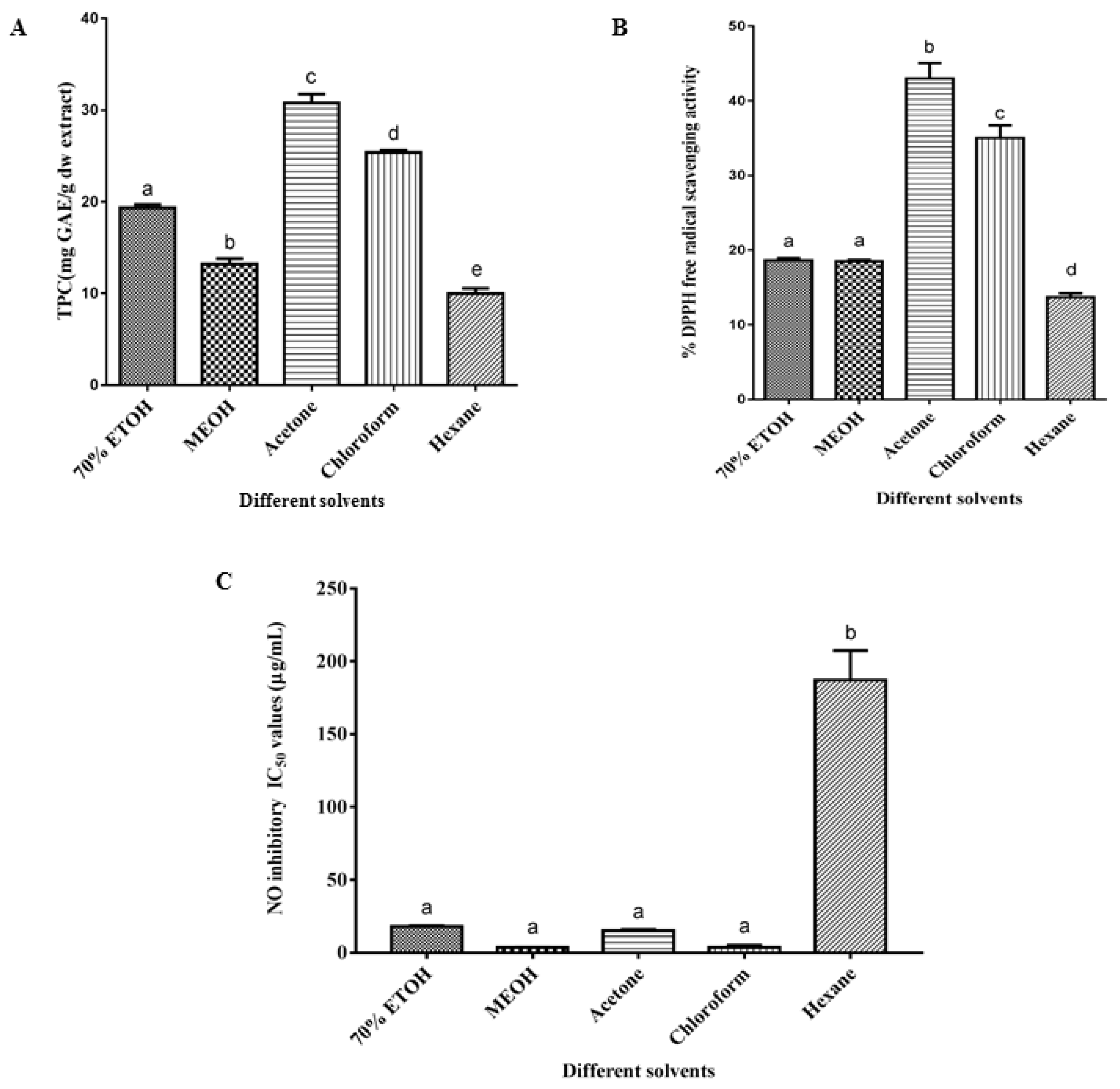
2.4. Effect of Different Solvent Extractions on Total Phenolic Content (TPC), 2,2-Diphenyl-1-Picrylhydrazyl (DPPH) Radical Scavenging and Nitric Oxide Inhibitory Activities
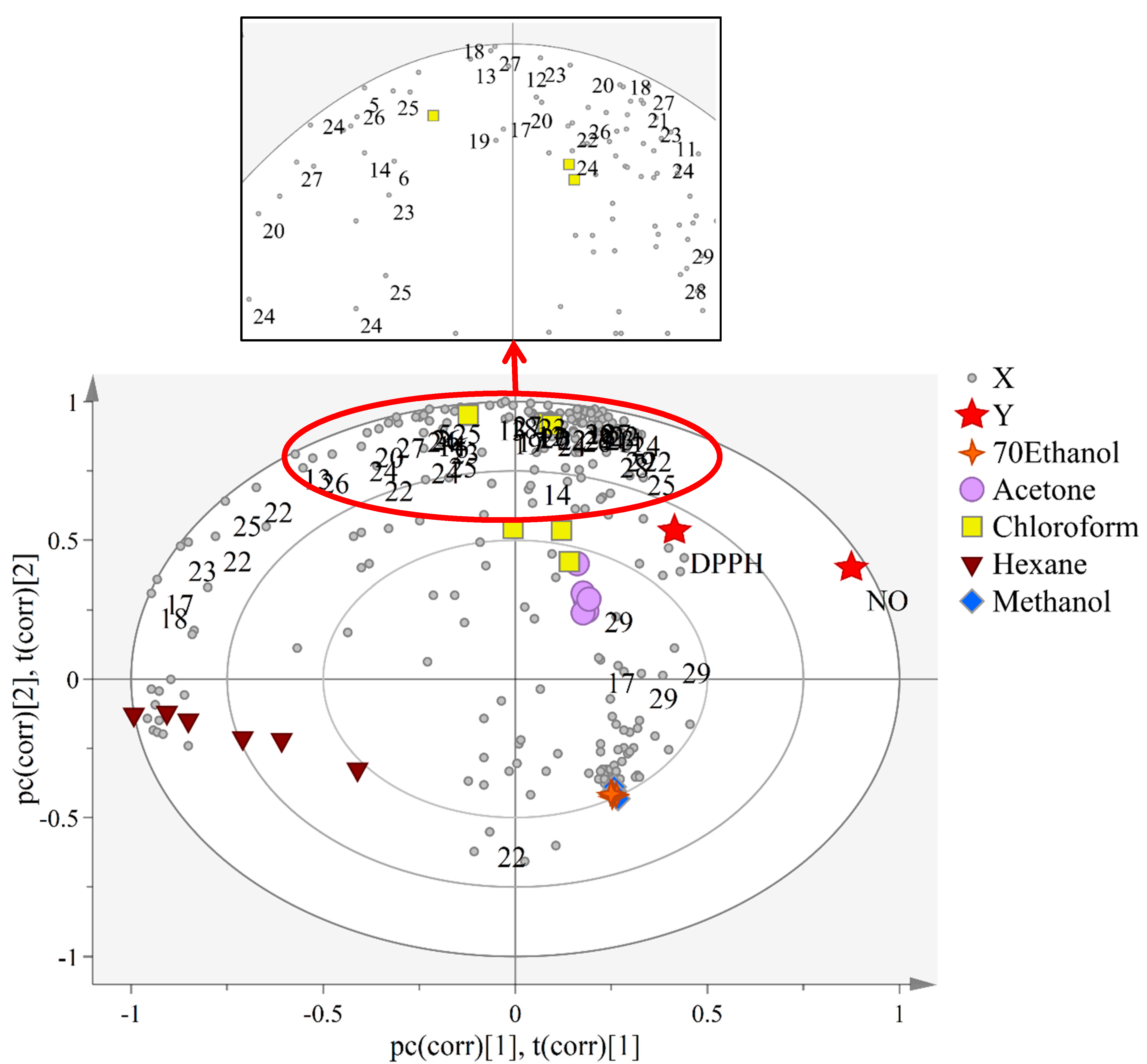
2.5. The Correlation Study between the Metabolites and Biological Activities in C. calcitrans Extracts
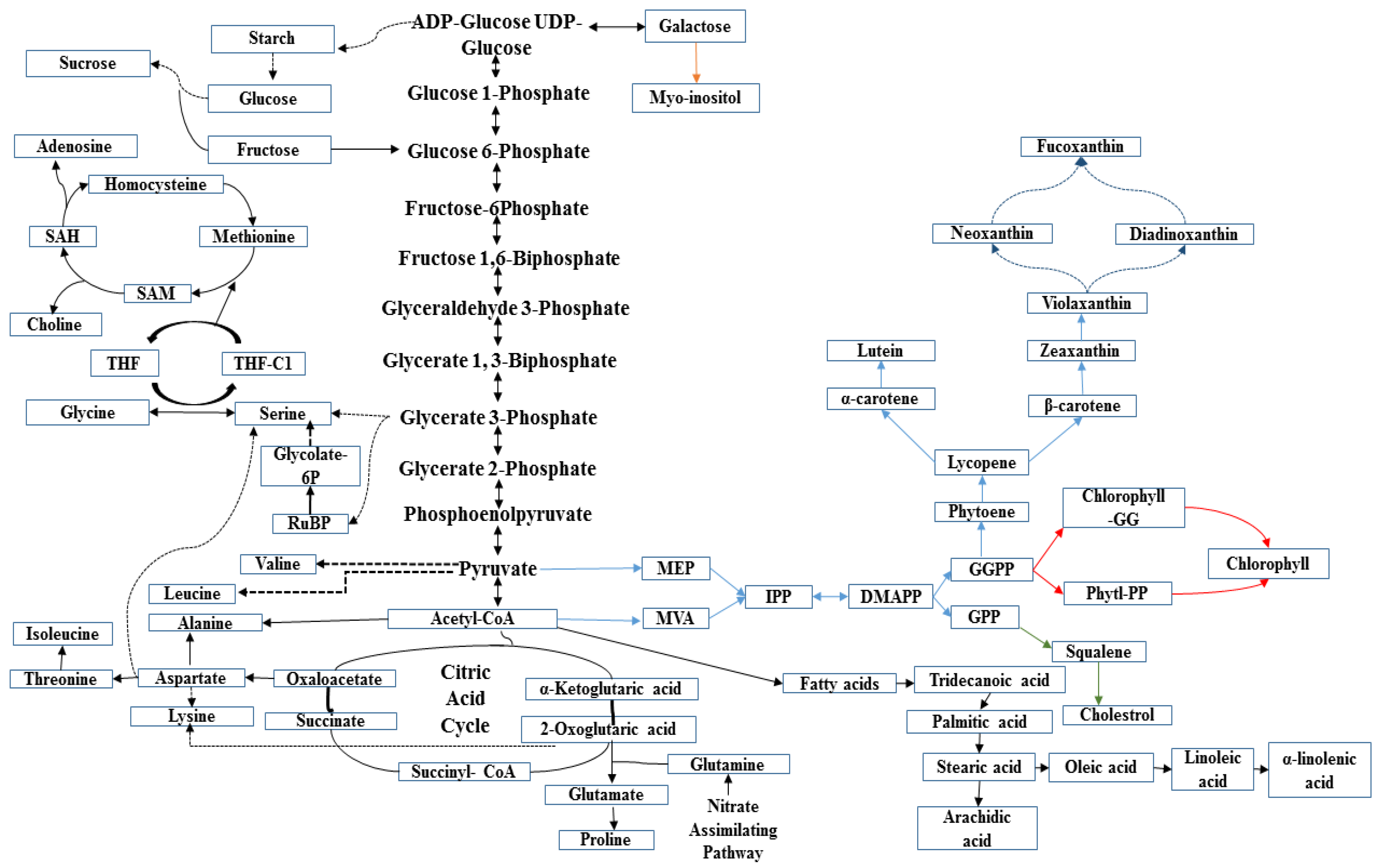
2.6. Metabolite Network Analysis in Diatom C. calcitrans
3. Materials and Methods
3.1. Marine Microalgal Material
3.2. Solvents and Chemicals
3.3. Microalgae Extraction Procedure
3.4. Sample Preparation for NMR Analysis
3.5. NMR Analyses
3.6. NMR Spectra Preprocessing and Multivariate Data Analysis
3.7. TPC Assay
3.8. 2,2-Diphenyl-1-picrylhydrazyl (DPPH) Free Radical Assay
3.9. Nitric Oxide (NO) Inhibitory Assay
3.10. Cell Viability
3.11. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stonik, V.S.; Stonik, I. Low-molecular-weight metabolites from diatoms: Structures, biological roles and biosynthesis. Mar. Drugs 2015, 13, 3672–3709. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwe, M.; Tedesco, L.; Arrigo, K.R.; Assmy, P.; Campbell, K.; Meiners, K.M.; Rintala, J.M.; Selz, V.; Thomas, D.N.; Stefels, J. Microalgal community structure and primary production in Arctic and Antarctic sea ice: A synthesis. Elem. Sci. Anth. 2018, 6, 1–25. [Google Scholar] [CrossRef]
- Gong, M.; Bassi, A. Carotenoids from microalgae: A review of recent developments. Biotechnol. Adv. 2016, 34, 1396–1412. [Google Scholar] [CrossRef] [PubMed]
- Macías-Sánchez, M.D.; Serrano, C.M.; Rodríguez, M.R.; de la Ossa, E.M.; Lubián, L.M.; Montero, O. Extraction of carotenoids and chlorophyll from microalgae with supercritical carbon dioxide and ethanol as cosolvent. J. Sep. Sci. 2008, 31, 1352–1362. [Google Scholar] [CrossRef] [PubMed]
- Grima, E.M.; Sanchez Perez, J.A.; Garcia Camacho, F.; Robles Medina, A.; Giminez, A.; Lopez Alonso, D. The production of polyunsaturated fatty acids by microalgae: From strain selection to product purification. Process Biochem. 1995, 30, 711–719. [Google Scholar] [CrossRef]
- Steinrücken, P.; Erga, S.R.; Miøs, S.A.; Khedival, H.; Prestegard, S.K. Bioprospecting North Atlantic microalgae with fast growth and high polyunsaturated fatty acid (PUFA) content for microalgae-based technology. Algal Res. 2017, 26, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Pangestuti, R.; Kim, S.K. Biological activities and health benefit effects of natural pigments derived from marine algae. J. Funct. Foods 2011, 3, 255–266. [Google Scholar] [CrossRef]
- Kris-Etherton, P.M.; Hecker, K.D.; Binkoski, A.E. Polyunsaturated fatty acids and cardiovascular health. Nutr. Rev. 2004, 62, 414–426. [Google Scholar] [CrossRef] [PubMed]
- Kuppusamy, P.; Soundharrajan, I.; Srigopalram, S.; Yusoff, M.M.; Maniam, G.P.; Govindan, N.; Cho, K.C. Potential pharmaceutical and biomedical applications of diatoms microalgae—An overview. Indian J. Geo-Mar. Sci. 2017, 46, 663–667. [Google Scholar]
- Foo, S.C.; Yusoff, F.M.; Ismail, M.; Basri, M.; Khong, N.M.H.; Chan, K.W.; Yau, S.K. Efficient solvent extraction of antioxidant-rich extract from a tropical diatom, Chaetoceros calcitrans (Paulsen) Takano 1968. Asian Pac. J. Trop. Biomed. 2015, 5, 834–840. [Google Scholar] [CrossRef]
- Foo, S.C.; Yusoff, F.M.; Ismail, M.; Basri, M.; Yau, S.K.; Khong, N.M.H.; Chan, K.W.; Ebrahimi, M. Antioxidant capacities of fucoxanthin-producing algae as influenced by their carotenoid and phenolic contents. J. Biotechnol. 2017, 241, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Foo, S.C.; Yusoff, F.M.; Ismail, M.; Basri, M.; Chan, K.W.; Khong, N.M.H.; Yau, S.K. Production of fucoxanthin-rich fraction (FxRF) from a diatom, Chaetoceros calcitrans (Paulsen) Takano 1968. Algal Res. 2015, 12, 26–32. [Google Scholar] [CrossRef]
- Nigjeh, S.E.; Yusoff, F.M.; Mohamed Alitheen, N.B.; Rasoli, M.; Keong, Y.S.; Omar, A.R. Cytotoxic effect of ethanol extract of microalga, Chaetoceros calcitrans, and its mechanisms in inducing apoptosis in human breast cancer cell line. Biomed. Res. Int. 2013, 2013, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Goh, S.H.; Alitheen, N.B.M.; Yusoff, F.M.; Yap, S.K.; Loh, S.P. Crude ethyl acetate extract of marine microalga, Chaetoceros calcitrans, induces Apoptosis in MDA-MB-231 breast cancer cells. Pharmacogn. Mag. 2014, 10, 1–8. [Google Scholar] [PubMed]
- Gauthier, M.J.; Bernard, P.; Aubert, M. Production of a photo-sensitive lipid antibiotic by the marine diatom Chaetoceros lauderi (Ralfs). Ann. Microbiol. 1978, 129, 63–70. [Google Scholar]
- Parsaeimehr, A.; Chen, Y.-F. Algal bioactive diversities against pathogenic microbes. In Microbiol Pathogens and Strategies for Combating Them: Science, Technology and Education; Méndez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2013; Series 4; pp. 796–803. [Google Scholar]
- Kaspar, H.F.; Keys, E.F.; King, N.; Smith, K.F.; Kesarcodi-Watson, A.; Miller, M.R. Continuous production of Chaetoceros calcitrans in a system suitable for commercial hatcheries. Aquaculture 2014, 420, 1–9. [Google Scholar] [CrossRef]
- Lu, W.; Su, X.; Klein, M.S.; Lewis, I.A.; Fiehn, O.; Rabinowitz, J.D. Metabolite measurement: Pitfalls to avoid and practices to follow. Annu. Rev. Biochem. 2017, 86, 277–304. [Google Scholar] [CrossRef] [PubMed]
- Goiris, K.; Muylaert, K.; Fraeye, I.; Foubert, I.; Brabanter, J.; Cooman, L. Antioxidant potential of microalgae in relation to their phenolic and carotenoid content. J. Appl. Phycol. 2012, 24, 1477–1486. [Google Scholar] [CrossRef]
- Maadane, A.; Merghoub, N.; Ainane, T.; El Arroussi, H.; Benhima, R.; Amzazi, S.; Bakri, Y.; Wahby, I. Antioxidant activity of some Moroccan marine microalgae: Pufa profiles, carotenoids and phenolic content. J. Biotechnol. 2015, 215, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Palanisamy, S.; Vinosha, M.; Marudhupandi, T.; Rajasekar, P.; Prabhu, N.M. Isolation of fucoidan from Sargassum polycystum brown algae: Structural characterization, in vitro antioxidant and anticancer activity. Int. J. Biol. Macromol. 2017, 102, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Kelman, D.; Posner, E.K.; McDermid, K.J.; Tabandera, N.K.; Wright, P.R.; Wright, A.D. Antioxidant activity of Hawaiian marine algae. Mar. Drugs 2012, 10, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Li, Y.; Karadeniz, F.; Kim, M.M.; Kim, S.K. α-Glycosidase and α-amylase inhibitory activities of phloroglucinal derivatives from edible marine brown alga, Ecklonia cava. J. Sci. Food Agric. 2009, 89, 1552–1558. [Google Scholar] [CrossRef]
- Englert, G.; Bjørnland, T.; Liaaen-Jensen, S. 1D and 2D NMR study of some allenic carotenoids of the fucoxanthin series. Magn. Reson. Chem. 1990, 28, 519–528. [Google Scholar] [CrossRef]
- Chauton, M.S.; Storseth, T.R.; Johnsen, G. High-resolution magic angle spinning 1H NMR analysis of whole cells of Thalassiosira pseudonana (Bacillariophyceae): Broad range analysis of metabolic composition and nutritional value. J. Appl. Phycol. 2003, 15, 533–542. [Google Scholar] [CrossRef]
- Chauton, M.S.; Størseth, T.R.; Krane, J. High-resolution magic angle spinning NMR analysis of whole cells of Chaetoceros muelleri (Bacillariophyceae) and comparison with 13C-NMR and distortion less enhancement by polarization transfer 13C-NMR analysis of lipophilic extracts. J. Phycol. 2004, 40, 611–618. [Google Scholar] [CrossRef]
- Boroujerdi, A.F.B.; Lee, P.A.; DiTullio, G.R.; Janech, M.G.; Vied, S.B.; Bearden, D.W. Identification of isethionic acid and other small molecule metabolites of Fragilariopsis cylindrus with nuclear magnetic resonance. Anal. Bioanal. Chem. 2012, 404, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Roberts, G.C.; Jardetzky, O. Nuclear magnetic resonance spectroscopy of amino acids, peptides, and proteins. Adv. Protein Chem. 1970, 24, 447–545. [Google Scholar] [PubMed]
- Kuczynska, P.; Jemiola-Rzeminska, M.; Strzalka, K. Photosynthetic pigments in diatoms. Mar. Drugs 2015, 13, 5847–5881. [Google Scholar] [CrossRef] [PubMed]
- Zuluaga, M.; Gueguen, V.; Pavon-Djavid, G.; Letourneur, D. Carotenoids from microalgae to block oxidative stress. BioImpacts 2017, 7, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Katz, J.J.; Brown, C.E. Nuclear magnetic resonance spectroscopy of chlorophylls and corrins. Structure 1983, 5, 3–49. [Google Scholar]
- Ginsburg, S.; Tiwari, P.; Kurhanewicz, J.; Madabhushi, A. Variable ranking with PCA: Finding multiparametric MR imaging markers for prostate cancer diagnosis and grading. In Proceedings of the International Workshop on Prostate Cancer Imaging Held in Conjunction with MICCAI 2011, Toronto, ON, Canada, 22 September 2011; Madabhushi, A., Dowling, J., Huisman, H., Barratt, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 146–157. [Google Scholar]
- Begum, H.; Yusoff, F.M.; Banerjee, S.; Khatoon, H.; Shariff, M. Availability and utilization of pigments from microalgae. Crit. Rev. Food Sci. Nutr. 2016, 56, 2209–2222. [Google Scholar] [CrossRef] [PubMed]
- Bryk, J.; Ochoa, J.B.; Correia, M.I.T.D.; Munera-Seeley, V.; Popovic, P.J. Effect of citrulline and glutamine on nitric oxide production in RAW 264.7 cells in an arginine-depleted environment. J. Parenter. Enter. Nutr. 2008, 32, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Bromke, M.A. Amino acid biosynthesis pathways in diatoms. Metabolites 2013, 3, 294–311. [Google Scholar] [CrossRef] [PubMed]
- Loewus, F.A.; Loewus, M.W. Myo-Inositol: Its biosynthesis and metabolism. Annu. Rev. Plant Physiol. 1983, 34, 137–161. [Google Scholar] [CrossRef]
- Miller, M.R.; Quek, S.Y.; Staehler, K.; Nalder, T.; Packer, M.A. Changes in oil content, lipid class and fatty acid composition of the microalga Chaetoceros calcitrans over different phases of batch culture. Aquac. Res. 2014, 45, 1634–1647. [Google Scholar] [CrossRef]
- Sirin, S.; Clavero, E.; Salvado, J. Efficient harvesting of Chaetoceros calcitrans for biodiesel production. Environ. Technol. 2015, 36, 1902–1912. [Google Scholar] [CrossRef] [PubMed]
- Talero, E.; García-Mauriño, S.; Ávila-Román, J.; Rodríguez-Luna, A.; Alcaide, A.; Motilva, V. Bioactive compounds isolated from microalgae in chronic inflammation and cancer. Mar. Drugs 2015, 13, 6152–6209. [Google Scholar] [CrossRef] [PubMed]
- Mizoguchi, T.; Isaji, M.; Yamano, N.; Harada, J.; Fujii, R.; Tamiaki, H. Molecular structures and functions of chlorophylls-a esterified with geranylgeranyl, dihydrogeranylgeranyl, and tetrahydrogeranylgeranyl groups at the 17-propionate residue in a diatom, Chaetoceros calcitrans. Biochemistry 2017, 56, 3682–3688. [Google Scholar] [CrossRef] [PubMed]
- Imaizumi, Y.; Nagao, N.; Yusoff, F.M.; Taguchi, S.; Toda, T. Estimation of optimum specific light intensity per cell on a high-cell-density continuous culture of Chlorella zofingiensis not limited by nutrients or CO2. Bioresour. Technol. 2014, 162, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Mediani, A.; Khatib, A.; Ismail, I.S.; Zawawi, N.; Abas, F. Comparison of partial least squares and random forests for evaluating relationship between phenolics and bioactivities of Neptunia oleracea. J. Sci. Food Agric. 2018, 98, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Adebayo, S.A.; Dzoyem, J.P.; Shai, L.J.; Eloff, J.N. The anti-inflammatory and antioxidant activity of 25 plant species used traditionally to treat pain in southern African. BMC Complement Altern. Med. 2015, 15, 1–10. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Metabolites | 1H NMR (Multiplicity) | 70% Et * | Me * | Ac * | Ch * | He * |
|---|---|---|---|---|---|---|---|
| 1 | Glutamic acid | 2.39 (m) | + | + | + | + | − |
| 2.14 (m) | + | + | + | + | − | ||
| 2.05 (m) | + | + | + | + | − | ||
| 2 | Proline | 4.10 (dd) | + | + | + | + | − |
| 2.35 (m) | + | + | + | + | − | ||
| 2.04 (m) | + | + | + | + | − | ||
| 1.96 (m) | + | + | + | + | − | ||
| 3 | Alanine | 1.49 (d) | + | + | + | + | − |
| 4 | Valine | 2.29 (m) | + | + | + | + | − |
| 1.03 (d) | + | + | + | + | − | ||
| 0.98 (d) | + | + | + | + | − | ||
| 5 | Isoleucine | 0.98 (d) | + | + | + | + | − |
| 0.94 (t) | + | + | + | + | − | ||
| 6 | Leucine | 1.69 (m) | + | + | + | + | − |
| 0.96 (d) | + | + | + | + | − | ||
| 7 | Methionine | 2.13 (m) | + | + | + | + | − |
| 8 | Choline | 4.05 (ddd) | + | + | − | − | − |
| 9 | Lysine | 1.87 (m) | + | + | + | + | + |
| 1.73 (m) | + | + | + | + | + | ||
| 10 | Glutamine | 2.43 (m) | + | + | + | + | + |
| 2.12 (m) | + | + | + | + | + | ||
| 11 | Glycine | 3.54 (s) | + | + | + | + | + |
| 12 | Cholesterol | 0.69 (s) | − | − | + | + | − |
| 0.87 (d) | − | − | + | + | + | ||
| 0.88 (d) | − | − | + | + | + | ||
| 0.92 (d) | − | − | + | + | + | ||
| 13 | Palmitic acid | 2.36 (m) | + | + | + | + | + |
| 1.66 (m) | + | + | + | + | + | ||
| 1.29 (m) | + | + | + | + | + | ||
| 0.90 (t) | + | + | + | + | + | ||
| 14 | Stearic acid | 1.77 (t) | − | − | + | + | + |
| 1.44 (t) | − | − | + | + | + | ||
| 1.01 (t) | − | − | + | + | + | ||
| 15 | Oleic acid | 5.39 (m) | − | − | + | + | + |
| 2.30 (t) | − | − | + | + | + | ||
| 1.96 (m) | − | − | + | + | + | ||
| 1.32 (m) | − | − | + | + | + | ||
| 0.88 (t) | − | − | + | + | + | ||
| 16 | Linoleic acid | 5.37 (m) | + | + | + | + | + |
| 2.36 (t) | + | + | + | + | + | ||
| 1.33 (m) | + | + | + | + | - | ||
| 0.90 (t) | + | + | + | + | + | ||
| 17 | α-Linolenic acid | 5.36 (m) | + | + | + | + | + |
| 2.80 (m) | + | + | + | + | + | ||
| 2.35 (t) | + | + | + | + | + | ||
| 2.04 (m) | + | + | + | + | + | ||
| 1.30 (m) | + | + | + | + | + | ||
| 0.96 (t) | + | + | + | + | + | ||
| 18 | Arachidic acid | 2.35 (t) | − | − | + | + | + |
| 1.63 (m) | − | − | + | + | + | ||
| 1.29 (m) | − | − | + | + | + | ||
| 0.88 (t) | − | − | + | + | + | ||
| 19 | Glucose | 5.20 (d) | + | + | + | + | − |
| 3.82 (m) | + | + | + | + | − | ||
| 3.52 (dd) | + | + | + | + | − | ||
| 20 | Sucrose | 5.39 (d) | + | + | + | + | − |
| 4.19 (d) | + | + | + | + | − | ||
| 3.82 (m) | + | + | + | + | − | ||
| 3.67 (s) | + | + | + | + | − | ||
| 3.46 (t) | + | + | + | + | − | ||
| 21 | Myo-inositol | 4.06 (t) | − | − | + | + | − |
| 22 | Fucoxanthin | 6.81 (dd) | − | − | + | + | − |
| 6.74 (dd) | − | − | + | + | − | ||
| 6.45 (dd) | − | − | + | + | − | ||
| 6.43 (d) | − | − | + | + | − | ||
| 3.64 (m) | − | − | + | + | − | ||
| 2.58 (d) | − | − | + | + | − | ||
| 2.31 (dd) | − | − | + | + | − | ||
| 2.15 (s) | − | − | + | + | − | ||
| 2.07 (s) | − | − | + | + | − | ||
| 1.99 (s) | − | − | + | + | − | ||
| 1.85 (dd) | − | − | + | + | − | ||
| 1.52 (dd) | − | − | + | + | − | ||
| 1.38 (dd) | − | − | + | + | − | ||
| 1.20 (s) | − | − | + | + | − | ||
| 1.08 (s) | − | − | + | + | − | ||
| 1.04 (s) | − | − | + | + | − | ||
| 0.97 (s) | − | − | + | + | − | ||
| 23 | Astaxanthin | 6.79 (d) | − | − | + | + | − |
| 6.20–6.70 (m, olefinic-H) | − | − | + | + | − | ||
| 6.51 (d) | − | − | + | + | − | ||
| 4.34 (dd) | − | − | + | + | − | ||
| 3.67 (s) | − | − | + | + | − | ||
| 2.01(s) | − | − | + | + | − | ||
| 1.98 (s) | − | − | + | + | − | ||
| 1.94 (s) | − | − | + | + | − | ||
| 1.82 (t) | − | − | + | + | − | ||
| 1.33 (s) | − | − | + | + | − | ||
| 1.21 (s) | − | − | + | + | − | ||
| 24 | Canthaxanthin | 6.20–6.70 (m, olefinic-H) | − | − | − | − | − |
| 1.86 (s) | − | + | + | + | − | ||
| 1.19 (s) | + | + | + | + | − | ||
| 25 | Lutein | 6.67–6.57 (m, olefinic-H) | − | − | + | + | − |
| 6.35 (d) | − | − | + | + | − | ||
| 6.26 (d) | − | − | + | + | − | ||
| 6.07–6.08 (m) | − | − | + | + | − | ||
| 5.43 (dd) | − | − | + | + | − | ||
| 2.40 (d) | − | − | + | + | − | ||
| 1.96 (s) | − | − | + | + | − | ||
| 1.91 (s) | − | − | + | + | − | ||
| 1.78–1.77 (m) | − | − | + | + | − | ||
| 1.74 (s) | − | − | + | + | − | ||
| 1.63 (s) | − | − | + | + | − | ||
| 1.07 (s) | − | − | + | + | − | ||
| 1.01 (s) | − | − | + | + | − | ||
| 0.86 (s) | − | − | + | + | − | ||
| 26 | Zeaxanthin | 1.98 (s) | + | + | + | + | + |
| 1.97 (s) | + | + | + | + | + | ||
| 1.74 (s) | + | + | + | - | + | ||
| 1.08 (s) | + | + | + | + | + | ||
| 27 | Violaxanthin | 1.98 (s) | + | + | + | + | + |
| 1.94 (s) | + | + | + | + | + | ||
| 1.15 (s) | + | + | + | + | − | ||
| 0.98 (s) | + | + | + | + | + | ||
| 28 | Chlorophyll a | 9.52 (s) | − | − | + | + | + |
| 29 | Chlorophyll c1 | 9.96 (s) | − | − | + | + | + |
| 9.90 (s) | − | − | + | + | + | ||
| 9.81 (s) | − | − | + | + | − | ||
| 8.29(s) | − | − | + | + | − |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azizan, A.; Ahamad Bustamam, M.S.; Maulidiani, M.; Shaari, K.; Ismail, I.S.; Nagao, N.; Abas, F. Metabolite Profiling of the Microalgal Diatom Chaetoceros Calcitrans and Correlation with Antioxidant and Nitric Oxide Inhibitory Activities via 1H NMR-Based Metabolomics. Mar. Drugs 2018, 16, 154. https://doi.org/10.3390/md16050154
Azizan A, Ahamad Bustamam MS, Maulidiani M, Shaari K, Ismail IS, Nagao N, Abas F. Metabolite Profiling of the Microalgal Diatom Chaetoceros Calcitrans and Correlation with Antioxidant and Nitric Oxide Inhibitory Activities via 1H NMR-Based Metabolomics. Marine Drugs. 2018; 16(5):154. https://doi.org/10.3390/md16050154
Chicago/Turabian StyleAzizan, Awanis, Muhammad Safwan Ahamad Bustamam, M. Maulidiani, Khozirah Shaari, Intan Safinar Ismail, Norio Nagao, and Faridah Abas. 2018. "Metabolite Profiling of the Microalgal Diatom Chaetoceros Calcitrans and Correlation with Antioxidant and Nitric Oxide Inhibitory Activities via 1H NMR-Based Metabolomics" Marine Drugs 16, no. 5: 154. https://doi.org/10.3390/md16050154
APA StyleAzizan, A., Ahamad Bustamam, M. S., Maulidiani, M., Shaari, K., Ismail, I. S., Nagao, N., & Abas, F. (2018). Metabolite Profiling of the Microalgal Diatom Chaetoceros Calcitrans and Correlation with Antioxidant and Nitric Oxide Inhibitory Activities via 1H NMR-Based Metabolomics. Marine Drugs, 16(5), 154. https://doi.org/10.3390/md16050154