Cyanopeptolins with Trypsin and Chymotrypsin Inhibitory Activity from the Cyanobacterium Nostoc edaphicum CCNP1411

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Results
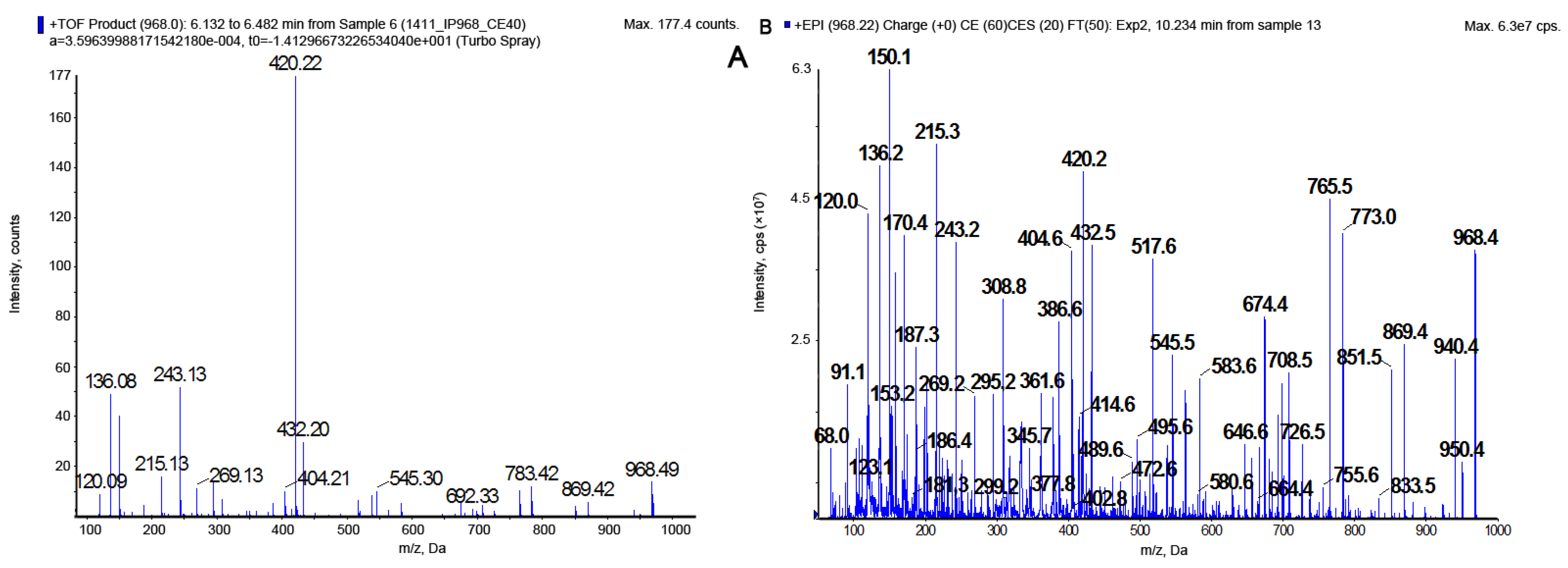
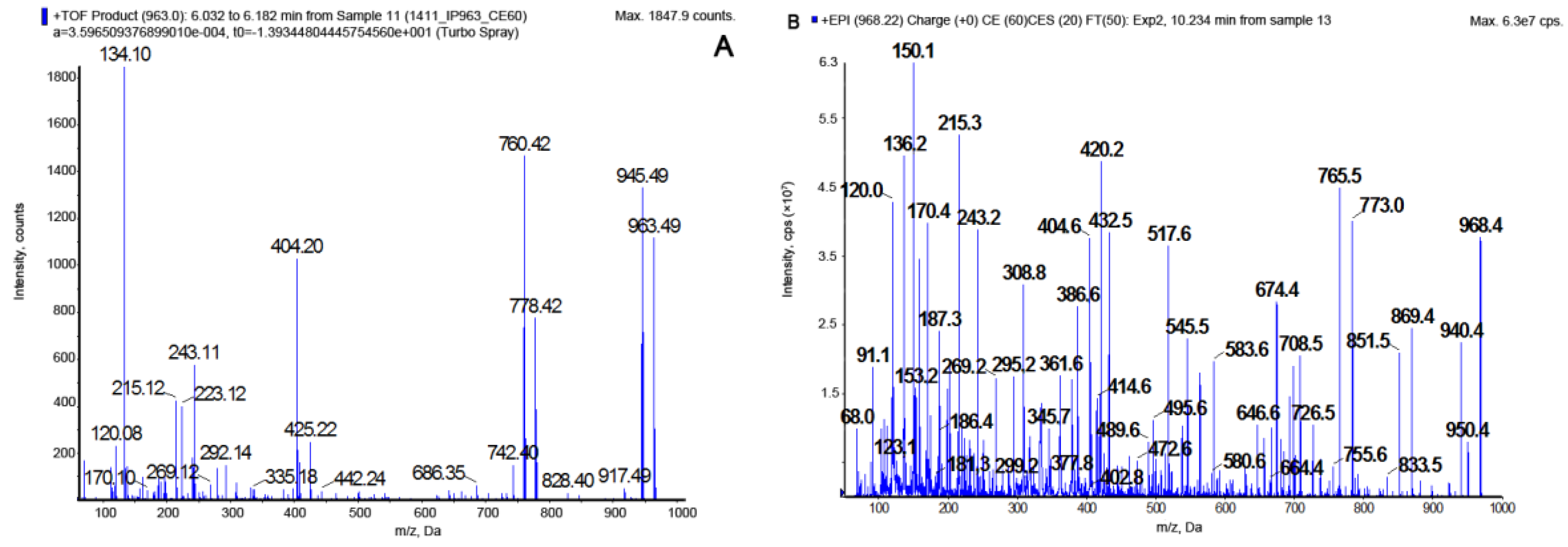
2.1. LC-MS/MS Analysis of Cyanopeptolins
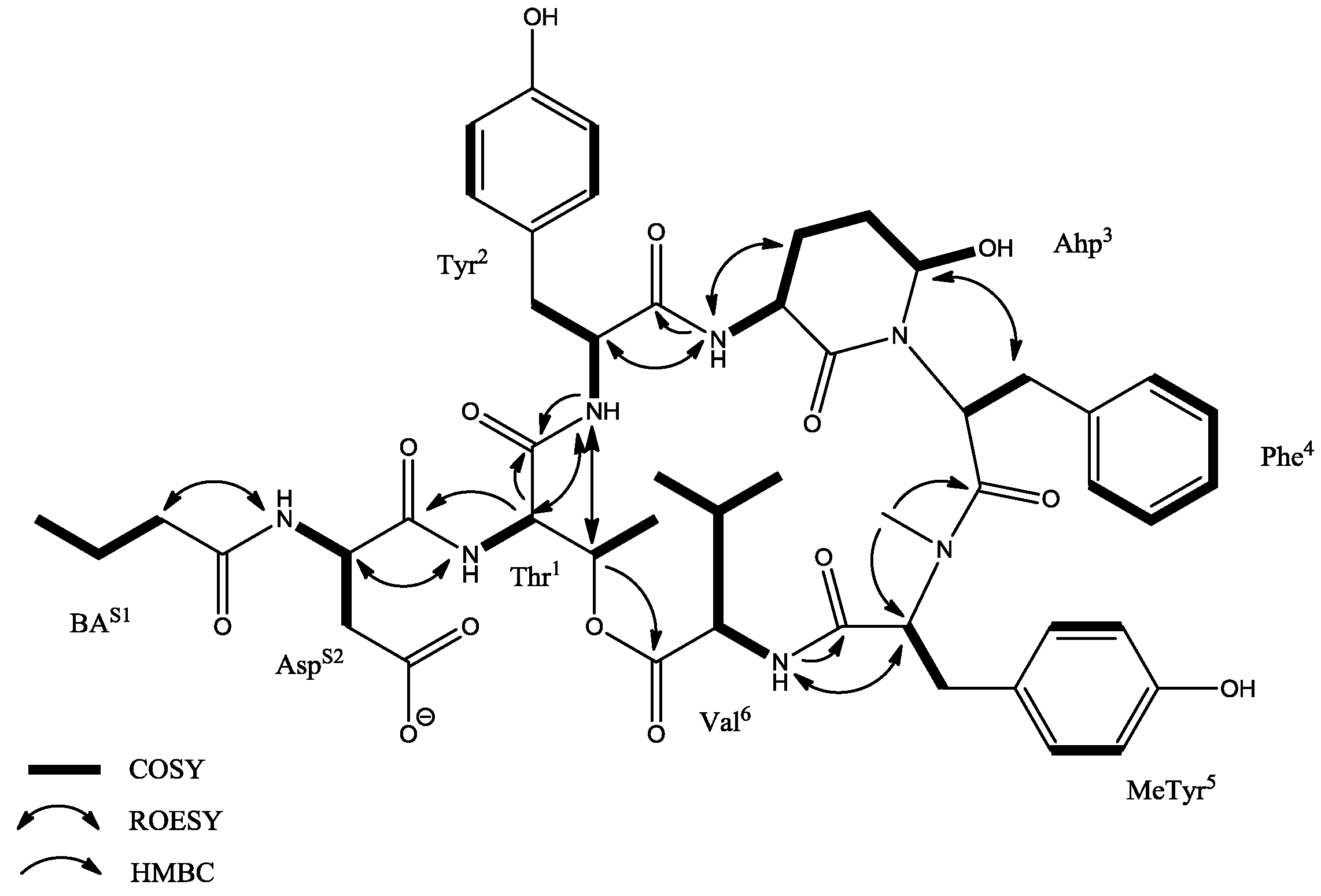
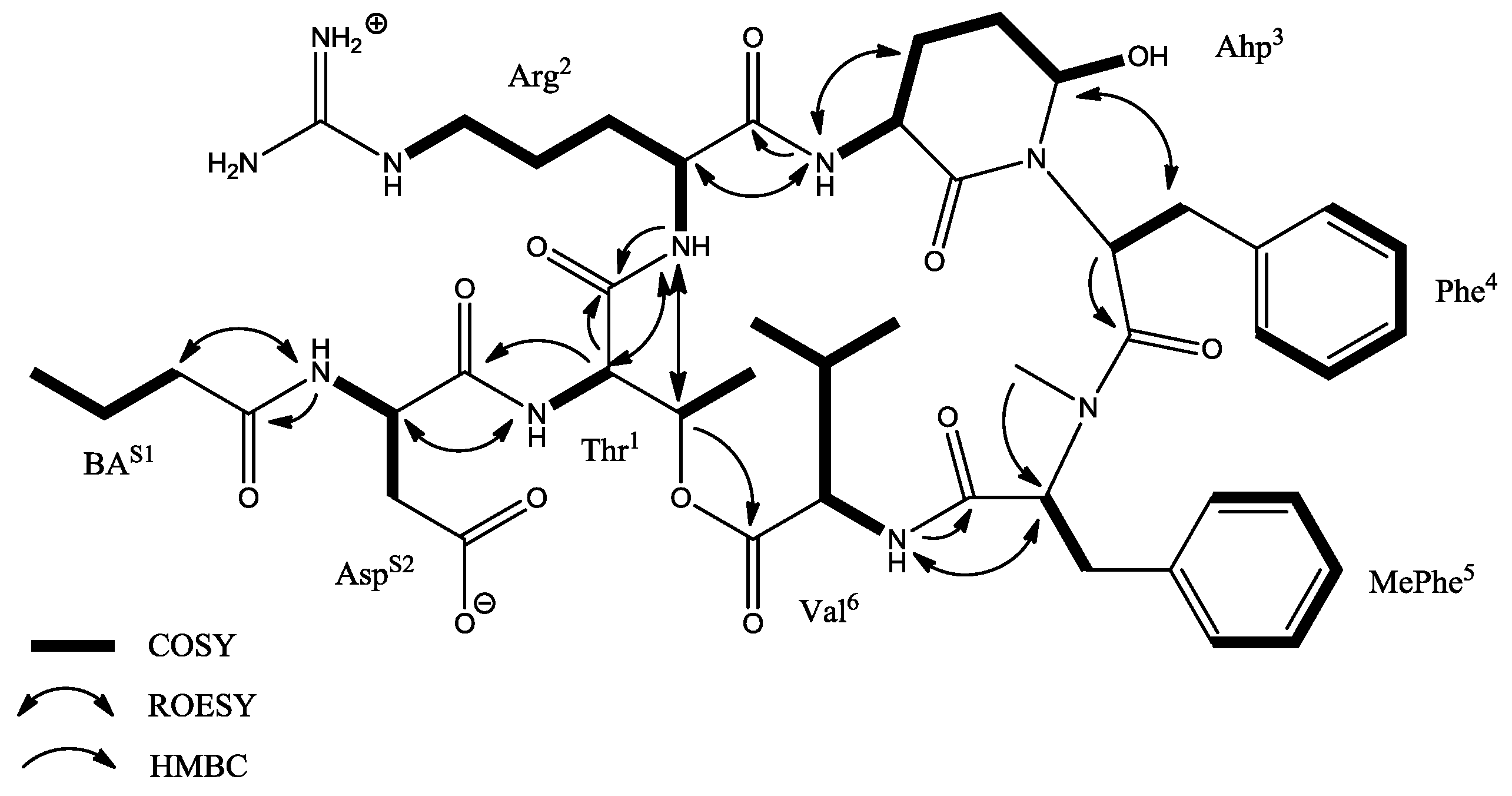
2.2. NMR Analysis
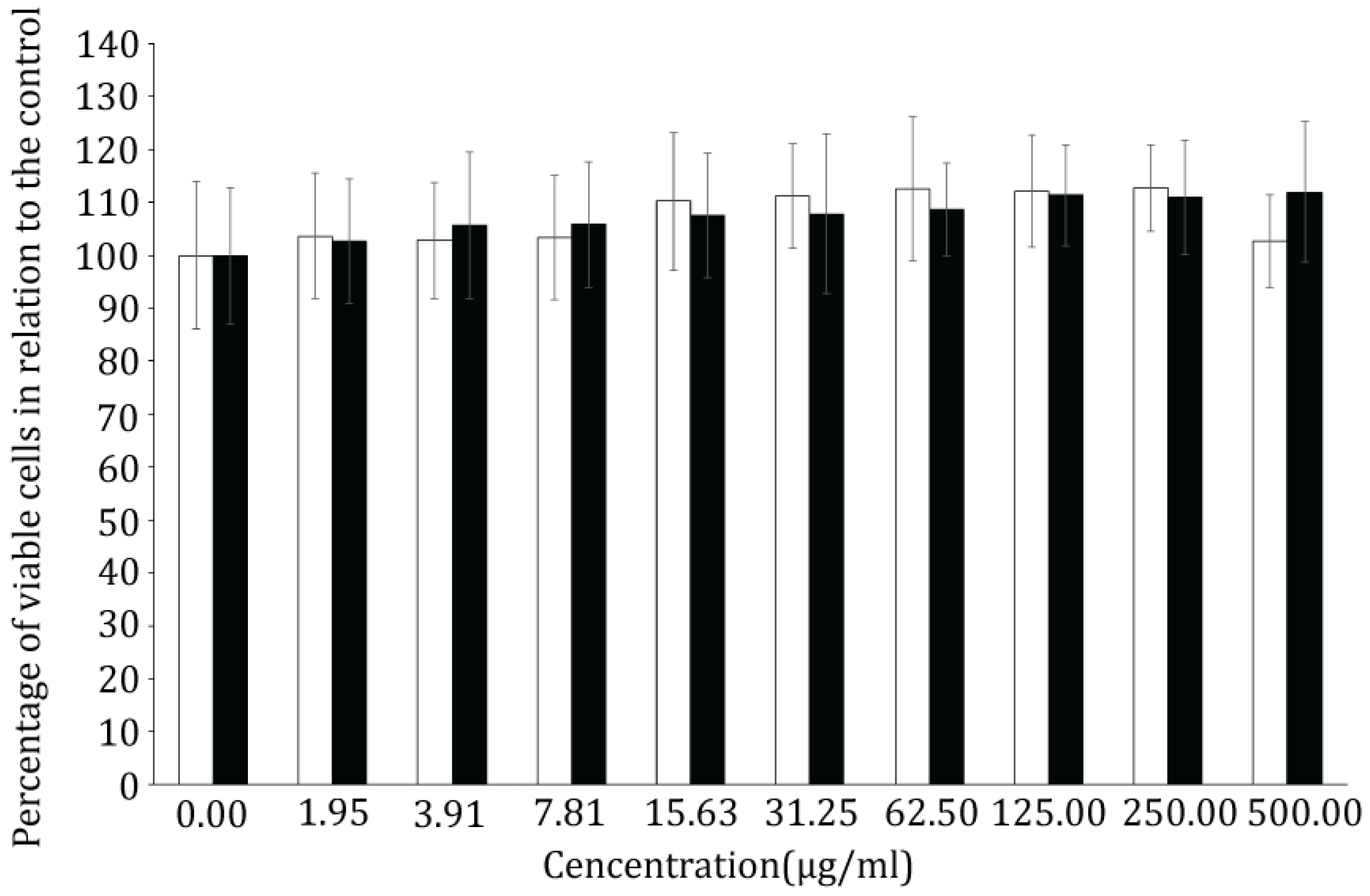
2.3. Bioassays
3. Discussion
4. Materials and Methods
4.1. Culture Conditions
4.2. Extraction and Isolation
4.3. LC-MS/MS Analyses
4.4. NMR Analyses
4.5. Enzyme Inhibition Assay
4.6. Cytotoxicity Assay
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Moore, R.E. Cyclic peptides and depsipeptides from cyanobacteria: A review. J. Ind. Microbiol. 1996, 16, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Gerwick, L.; Gerwick, W.H.; Coates, R.C.; Engene, N.; Grindberg, R.V.; Jones, A.C.; Sorrels, C.M. Giant marine cyanobacteria produce exciting potential pharmaceuticals. Microbe 2008, 3, 277–284. [Google Scholar] [CrossRef]
- Nagarajan, M.; Maruthanayagam, V.; Sundararaman, M. A review of pharmacological and toxicological potentials of marine cyanobacterial metabolites. J. Appl. Toxicol. 2011, 33, 153–185. [Google Scholar] [CrossRef] [PubMed]
- Sainis, I.; Fokas, D.; Vareli, K.; Tzakos, A.G.; Kounnis, V.; Briasoulis, E. Cyanobacterial cyclopeptides as lead compounds to novel targeted cancer drugs. Mar. Drugs 2010, 8, 629–657. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.; Costa-Rodrigues, J.; Fernandes, M.H.; Barros, P.; Vasconcelos, V.; Martins, R. Marine cyanobacteria compounds with anticancer properties: A Review on the implication of apoptosis. Mar. Drugs 2012, 10, 2181–2207. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Luesch, H. Largazole: From discovery to broad-spectrum therapy. Nat. Prod. Rep. 2012, 29, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Salvador-Reyes, L.A.; Luesch, H. Biological targets and mechanisms of action of natural products from marine cyanobacteria. Nat. Prod. Rep. 2015, 32, 478–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ersmark, K.; Del Valle, J.R.; Hanessian, S. Chemistry and biology of the aeruginosin family of serine protease inhibitors. Angew. Chem. Int. Ed. Engl. 2008, 47, 1202–1223. [Google Scholar] [CrossRef] [PubMed]
- Chlipala, G.; Mo, S.; Orjala, J. Chemodiversity in freshwater and terrestrial cyanobacteria—A source for drug discovery. Curr. Drug Targets 2011, 12, 1654–1673. [Google Scholar] [CrossRef] [PubMed]
- Elkobi-Peer, S.; Carmeli, S. New prenylated aeruginosin, microphycin, anabaenopeptin and micropeptin analogues from microcystis bloom material collected in Kibbutz Kfar Blum, Israel. Mar. drugs 2015, 13, 2347–2375. [Google Scholar] [CrossRef] [PubMed]
- Itou, Y.; Ishida, K.; Shin, H.; Murakami, M. Oscillapeptins A to F, serine protease inhibitors from the three strains of Oscillatoria agardhii. Tetrahedron 1999, 55, 6871–6882. [Google Scholar] [CrossRef]
- Gesner-Apter, S.; Carmeli, S. Protease inhibitors from a water bloom of the cyanobacterium Microcystis aeruginosa. J. Nat. Prod. 2009, 72, 1429–1436. [Google Scholar] [CrossRef] [PubMed]
- Welker, M.; von Döhren, H. Cyanobacterial peptides—Nature’s own combinatorial biosynthesis. FEMS Microbiol. Rev. 2006, 30, 530–563. [Google Scholar] [CrossRef] [PubMed]
- Cadel-Six, S.; Dauga, C.; Castests, A.; Rippka, R.; Bouchier, C.; Tandeau de Marsac, N.; Welker, M. Halogenase genes in nonribosomal peptide synthetase gene clusters of Microcystis (Cyanobacteria): Sporadic distribution and evolution. Mol. Biol. Evol. 2008, 25, 2031–2041. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.; Oberer, L.; Ino, T.; König, W.; Busch, M.; Weckesser, J. Cyanopeptolins, new depsipeptides from the cyanobacterium Microcystis sp. PCC 7806. J. Antibiot. 1993, 46, 1550–1556. [Google Scholar] [CrossRef] [PubMed]
- Czarnecki, O.; Henning, M.; Lippert, I.; Welker, M. Identification of peptide metabolites of Microcystis (Cyanobacteria) that inhibit trypsin-like activity in planktonic herbivorous Daphnia (Cladocera). Environ. Microbiol. 2006, 8, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Welker, M.; Brunke, M.; Preussel, K.; Lippert, I.; von Döhren, H. Diversity and distribution of Microcystis (Cyanobacteria) oligopeptide chemotypes from natural communities studied by single-colony mass spectrometry. Microbiology 2004, 150, 1785–1796. [Google Scholar] [CrossRef] [PubMed]
- Jakobi, C.; Rinehart, K.; Neuber, R.; Mez, K.; Weckesser, J. Cyanopeptolin SS, a disulphated depsipeptide from a water bloom: Structural elucidation and biological activities. Phycologia 1996, 35, 111–116. [Google Scholar] [CrossRef]
- Tsukamoto, S.; Painuly, P.; Young, K.; Yang, X.; Shimizu, Y. Microcystilide A: A novel cell-differentiation-promoting depsipeptide from Microcystis aeruginosa NO-15-1840. J. Am. Chem. Soc. 1993, 115, 11046–11047. [Google Scholar] [CrossRef]
- Harada, K.; Mayumi, T.; Shimada, T.; Fuji, K.; Kondo, F.; Park, H.; Watanabe, M. Co-production of microcystins and aeruginopeptins by natural cyanobacterial bloom. Environ. Toxicol. 2001, 16, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Adiv, S.; Aharonv-Nadborny, R.; Carmeli, S. Micropeptins from Microcystis aeruginosa collected in Dalton reservoir, Israel. Tetrahedron 2010, 66, 7429–7436. [Google Scholar] [CrossRef]
- Rounge, T.; Rohrlack, T.; Tooming-Klunderud, A.; Kristensen, T.; Jakobsen, K. Comparison of cyanopeptolin genes in Planktothrix, Microcystis and Anabaena strains: Evidence for independent evolution within each genus. Appl. Environ. Microbiol. 2007, 73, 7322–7330. [Google Scholar] [CrossRef] [PubMed]
- Tooming-Klunderud, A.; Rohrlack, T.; Shalchian-Tabrizi, K.; Kristensen, T.; Jakobsen, K. Structural analysis of non-ribosomal halogenated cyclic peptide and its putative operon from Microcystis: Implications for evolution of cyanopeptolins. Microbiol. 2007, 153, 1382–1393. [Google Scholar] [CrossRef] [PubMed]
- Rouhiainen, L.; Paulin, L.; Suomalainen, S.; Hyytiäinen, H.; Buikema, W.; Haselkorn, R.; Sivonen, K. Genes encoding synthetases of cyclic depsipeptides, anabaenopeptilides, in Anabaena strain 90. Mol. Microbiol. 2000, 37, 156–167. [Google Scholar] [CrossRef] [PubMed]
- Weckesser, J.; Martin, C.; Jakobi, C. Cyanopeptolins, depsipeptides from cyanobacteria. System. Appl. Microbiol. 1996, 19, 133–138. [Google Scholar] [CrossRef]
- Namikoshi, M.; Rinehart, K. Bioactive compounds produced by cyanobacteria. J. Ind. Microbiol. Biotechnol. 1996, 17, 373–384. [Google Scholar] [CrossRef]
- Bister, B.; Keller, S.; Baumann, H.; Nicholson, G.; Weist, S.; Jung, G.; Süssmuth, R.; Jüttner, F. Cyanopeptolin 963 A, a chymotrypsin inhibitor of Microcystis PCC 7806. J. Nat. Prod. 2004, 67, 1755–1757. [Google Scholar] [CrossRef] [PubMed]
- Yamaki, H.; Sitachitta, N.; Sano, T.; Kaya, K. Two new chymotrypsin inhibitors isolated from the cyanobacterium Microcystis aeruginosa NIES-88. J. Nat. Prod. 2005, 68, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Linington, R.; Edwards, D.; Shuman, C.; McPhail, K.; Matainaho, T.; Gerwick, W. Symplocamide A, a potent cytotoxin and chymotrypsin inhibitor from the marine cyanobacterium Symploca sp. J. Nat. Prod. 2008, 71, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Okumura, H.; Philmus, B.; Portmann, C.; Hemscheidt, T. Homotyrosine-containing cyanopeptolins 880 & 960 and anabaenopeptins 908 & 915 from Planktothrix agardhii CYA 126/8. J. Nat. Prod. 2009, 72, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Zainuddin, E.; Mentel, R.; Wray, V.; Jansen, R.; Nimtz, M.; Lalk, M.; Mundt, S. Cyclic depsipeptides, ichthyopeptins A and B, from Microcystis ichthyoblabe. J. Nat. Prod. 2007, 70, 1084–1088. [Google Scholar] [CrossRef] [PubMed]
- Gademann, K.; Portmann, C.; Blom, J.; Zeder, M.; Jüttner, F. Multiple toxin production in the cyanobacterium Microcystis: Isolation of the toxic protease inhibitor cyanopeptolin 1020. J. Nat. Prod. 2010, 73, 980–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blom, J.; Bister, B.; Bischoff, D.; Nicholson, G.; Jung, G.; Süssmuth, R.; Jüttner, F. Oscillapeptin J, a new grazer toxin of the freshwater cyanobacterium Planktothrix rubescens. J. Nat. Prod. 2003, 66, 431–434. [Google Scholar] [CrossRef] [PubMed]
- Kaya, K.; Sano, T.; Beattie, K.; Codd, G. Nostocyclin, a novel 3-amino-6-hydroxy-2-piperidone-containing cyclic depsipeptide from the cyanobacterium Nostoc sp. Tetrahedron Lett. 1996, 37, 6725–6728. [Google Scholar] [CrossRef]
- Okino, T.; Qi, S.; Matsua, H.; Murakami, M.; Yamaguchi, K. Nostopeptins A and B, elastase inhibitors from the cyanobacterium Nostoc minutum. J. Nat. Prod. 1997, 60, 158–161. [Google Scholar] [CrossRef]
- Ploutno, A.; Carmeli, S. Modified peptides from a water bloom of the cyanobacterium Nostoc sp. Tetrahedron 2002, 58, 9949–9957. [Google Scholar] [CrossRef]
- Dembitsky, V.; Řezanka, T. Metabolites produced by nitrogen-fixing Nostoc species. Folia Microbiol. 2005, 50, 363–391. [Google Scholar] [CrossRef]
- Mehner, C.; Müller, D.; Kehraus, S.; Hautmann, S.; Gütschow, M.; König, G. New peptolides from the cyanobacterium Nostoc insulare as selective and potent inhibitors of human leukocyte elastase. ChemBioChem 2008, 9, 2692–2703. [Google Scholar] [CrossRef] [PubMed]
- Hastie, J.; Borthwick, E.; Morrison, L.; Codd, G.; Cohen, P. Inhibition of several protein phosphatases by a non-covalently interacting microcystin and a novel cyanobacterial peptide, nostocyclin. BBA 2005, 1726, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Piccardi, R.; Frosini, A.; Tredici, M.; Margheri, M. Bioactivity in free-living and symbiotic cyanobacteria of the genus Nostoc. J. Appl. Phycol. 2000, 12, 543–547. [Google Scholar] [CrossRef]
- Ploutno, A.; Carmeli, S. Banyasin A and banyasides A and B, three novel modified peptides from water bloom of the cyanobacterium Nostoc sp. Tetrahedron 2005, 61, 575–583. [Google Scholar] [CrossRef]
- Tidgewell, K.; Clark, B.; Gerwick, W. The Natural Products Chemistry of Cyanobacteria; University of California San Diego: La Jolla, CA, USA, 2010; pp. 142–187. [Google Scholar]
- Liu, L.; Jokela, J.; Herfindal, L.; Wahlsten, M.; Sinkkonen, J.; Permi, P.; Fewer, D.; Ove Døskeland, S.; Sivonen, K. 4-methylproline guided natural product discovery: Co-occurrence of 4-hydroxy- and 4-methylprolines in nostoweipeptins and nostopeptolides. ACS Chem. Biol. 2014, 9, 2646–2655. [Google Scholar] [CrossRef] [PubMed]
- Trimurtulu, G.; Patterson, G.; Corbett, T.; Ohtani, I.; Moore, R.; Valeriote, F. Total structures of cryptophycins, potent antitumor depsipeptides from the blue-green alga Nostoc sp. GSV 224. J. Am. Chem. Soc. 1994, 116, 4729–4737. [Google Scholar] [CrossRef]
- Weiss, C.; Figueras, E.; Borbely, A.; Sewald, N. Cryptophycins: Cytotoxic cyclodepsipeptides with potential for tumor targeting. J. Pept. Sci. 2017, 23, 514–531. [Google Scholar] [CrossRef] [PubMed]
- Dey, B.; Lerner, D.; Lusso, P.; Boyd, M.; Elder, J.; Berger, E. Multiple antiviral activities of cyanovirin-N: Blocking of human immunodeficiency virus type 1 gp120 interaction with CD4 and coreceptor and inhibition of diverse enveloped viruses. J. Virol. 2000, 74, 4562–4569. [Google Scholar] [CrossRef] [PubMed]
- Mayumi, T.; Kato, H.; Kawasaki, Y.; Harada, K. Formation of diagnostic product ions from cyanobacterial cyclic peptides by the two-bond fission mechanism using ion trap liquid chromatography/multi-stage mass spectrometry. Rapid Commun. Mass Spectrom. 2007, 21, 1025–1033. [Google Scholar] [CrossRef] [PubMed]
- Welker, M.; Christiansen, G.; von Döhren, H. Diversity of coexisting Planktothrix (Cyanobacteria) chemotypes deduced by mass spectral analysis of microcystins and other oligopeptides. Arch. Microbiol. 2004, 182, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Oh, S.; Yih, W.; Chin, J.; Kang, H.; Rho, J. Cyanopeptoline CB071: A cyclic depsipeptide isolated from the freshwater cyanobacterium Aphanocapsa sp. Chem. Pharm. Bull. 2008, 56, 1191–1193. [Google Scholar] [CrossRef] [PubMed]
- Okano, T.; Sano, T.; Kaya, K. Micropeptin T-20, a novel phosphate-containing cyclic depsipeptide from the cyanobacterium Microcystis aeruginosa. Tetraherdon Lett. 1999, 40, 2379–2382. [Google Scholar] [CrossRef]
- Srikanth, S.; Chen, Z. Plant protease inhibitors in therapeutics-focus on cancer therapy. Front. Pharmacol. 2016, 7, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Blom, J.; Baumann, H.; Codd, G.; Jüttner, F. Sensitivity and adaptation of aquatic organisms to oscillapeptin J and [D-Asp3, (E)-Dhb7] microcystin-RR. Arch. Hydrobiol. 2006, 167, 547–559. [Google Scholar] [CrossRef]
- Faltermann, S.; Hutter, S.; Christen, V.; Hettich, T.; Fent, K. Anti-inflammatory activity of cyanobacterial serine protease inhibitors aeruginosin 828A and cyanopeptolin 1020 in human hepatoma cell line Huh7 and effects in Zebrafish (Danio rerio). Toxins 2016, 8, 219. [Google Scholar] [CrossRef] [PubMed]
- Faltermann, S.; Zucchi, S.; Kohler, E.; Blom, J.; Pernthaler, J.; Fent, K. Molecular effects of the cyanobacterial toxin cyanopeptolin (CP1020) occurring in algal blooms: Global transcriptome analysis in zebrafish embryos. Aquat. Toxicol. 2014, 149, 33–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotai, J. Introduction for Preparation of Modified Nutrient Solution Z8 for Algae; Norwegian Institute for Water Research Publication B-117669: Oslo, Norway, 1972; 5p. [Google Scholar]
- Ocampo Bennet, X. Peptide au Seiner Cyanobakterien Wasserblütte (1998) aus dem Wannsee/Berli: Strukturen and Biologische Wirksamkeit; University of Freiburg: Freiburg, Germany, 2007; 28p. [Google Scholar]
- Kwan, J.; Taori, K.; Paul, V.; Luesch, H. Lyngbyastatins 8–10, elastase inhibitors with cyclic depsipeptide scaffolds isolated from the marine cyanobacterium Lyngbya semiplena. Mar. Drugs 2009, 7, 528–538. [Google Scholar] [CrossRef] [PubMed]
- Rapala, J.; Erkomaa, K.; Kukkonen, J.; Sivonen, K.; Lahti, K. Detection of microcystins with protein phosphatase inhibition assay, high-performance liquid chromatography–UV detection and enzyme-linked immunosorbent assay. Comparison of methods. Anal. Chim. Acta 2002, 466, 213–231. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecular Mass | Peptide Name | Structure | Enzyme Inhibition | References |
|---|---|---|---|---|
| 921 | Nostopeptin BN920 | [Thr+Leu+Ahp 1 +Phe+MeTyr+Val]Gln+Ac 2 | Chymotrypsin (IC50 0.11 μM) | [36] |
| 926 | Nostopeptin B | [Hmp 3 +Leu+Ahp+Ile+MeTyr+Ile]Gln+Ac | Elastase (IC50 11.0 μg/mL) Chymotrypsin (IC50 1.6 μg/mL) | [35] |
| 937 | Nostopeptin A | [Hmp+Leu+Ahp+Ile+MeTyr+Ile]Gln+BA 4 | Elastase (IC50 1.3 μg/mL) Chymotrypsin (IC50 1.4 μg/mL) | [35] |
| 942 | Insulapeptolide A | [Hmp+Leu+Ahp+Ile+MeTyr+Val]Cit 5 +Ac | HLE 10 (IC50 0.14 μM) * | [38] |
| 956 | Insulapeptolide B | [Hmp+Leu+Ahp+Leu+MeTyr+Ile]Cit+Ac | HLE (IC50 0.10 μM) * | |
| 956 | Insulapeptolide C | [Hmp+Leu+Ahp+Ile+diMeTyr 6 +Val]Cit+Ac | HLE (IC50 0.090 μM) * | |
| 970 | Insulapeptolide D | [Hmp+Leu+Ahp+Ile+diMeTyr+Ile]Cit+Ac | HLE (IC50 0.085 μM) * | |
| 991 | Insulapeptolide G | [Thr+Hph 7 +Ahp+Thr+MePhe+Val]Ser+Pro+BA | HLE (IC50 3.5 μM) * | |
| 1005 | Insulapeptolide H | [Thr+Hph+Ahp+Thr+MeTyr+Ile]Ser+Pro+BA | HLE (IC50 2.7μM) * | |
| 1007 | Insulapeptolide F | [Thr+Hph+Ahp+Thr+MeTyr+Val]Ser+Pro+BA | HLE (IC50 1.6 μM) * | |
| 1021 | Insulapeptolide E | [Thr+Hph+Ahp+Thr+MeTyr+Ile]Ser+Pro+BA | HLE (IC50 3.2 μM) * | |
| 1116 | Nostocyclin | [Thr+Hse 8 +Ahp+Phe+MeTyr+Val]Hse+Ile+Hpla 9 | PP1 (IC50 64.0 μM) | [34] |
| Cyanopeptolin CP | m/z | Structure | Enzyme Inhibition (IC50 [µM]) | ||||
|---|---|---|---|---|---|---|---|
| Chymotrypsin | Trypsin | Elastase | Thrombin | Protein Phosphatase 1 | |||
| CP 1048 | 1049 | [Thr+Arg+Ahp+Phe+MeHty+Val]Asp+OA | * | ** | - | - | - |
| CP 1020 | 1021 | [Thr+Arg+Ahp+Phe+MeHty+Val Asp+HA | 3.1 | 0.25 | - | - | - |
| CP 1018 | 1019 | [Thr+Arg+Ahp+Phe+MePhe+Val]Asp+ OA | - | 0.24 | - | - | - |
| CP 1006 | 1007 | [Thr+Arg+Ahp+Phe+MeTyr+Val]Asp+HA | * | ** | - | - | - |
| CP 992 | 993 | [Thr+Arg+Ahp+Phe+MeHty+Val]Asp+BA | 3.5 | 0.24 | - | - | - |
| CP 990 | 991 | [Thr+Arg+Ahp+Phe+MePhe+Val]Asp+HA | - | ** | - | - | - |
| CP 978 | 979 | [Thr+Arg+Ahp+Phe+MeTyr+Val]Asp+BA | 3.8 | 0.26 | - | - | - |
| CP 962 | 963 | [Thr+Arg+Ahp+Phe+MePhe+Val]Asp+BA | - | ** | - | - | - |
| CP 1027 | 1010 | [Thr+Tyr+Ahp+Phe+MeHty+Val]Asp+HA | 0.26 | - | - | - | - |
| CP 1013 | 996 | [Thr+Tyr+Ahp+Phe+MeTyr+Val]Asp+HA | ** | - | - | - | - |
| CP 999 | 982 | [Thr+Tyr+Ahp+Phe+MeHty+Val]Asp+BA | ** | - | -. | - | - |
| CP 985 | 968 | [Thr+Tyr+Ahp+Phe+MeTyr+Val]Asp+BA | 0.26 | - | - | - | - |
| CP 969 | 952 | [Thr+Tyr+Ahp+Phe+MePhe+Val]Asp+BA | ** | - | - | - | - |
| Unit | Position | δC | δH (J in Hz) | ROESY | HMBC a |
|---|---|---|---|---|---|
| BA | 1 2 3 4 | 172.9, C 37.7, CH2 19.2, CH2 14.2, CH3 | 2.11, m 1.56, m 0.89, t (7.2, 7.2) | Asp-NH | BA-1, BA-3, BA-4 BA-1, BA-2, BA-4 BA-2 |
| Asp | 1 2 3a 3b 4 NH | 171.7, C 49.9, CH 36.1, CH2 172.4, C | 4.58, dd (8.0, 5.0) 2.65, m 2.46, m 8.26, m | Thr-NH | Asp-4 |
| Thr1 | 1 2 3 4 NH | 168.9, C 54.7, CH 72.6, CH 18.0, CH3 | 4.52, d (10.0) 5.36, q (6.8, 6.9, 6.8) 1.16, d (7.2) 7.52, d (9.3) | Tyr-NH Tyr-NH Asp-2 | Thr-1, Asp-1 Val-1 Thr-2, Thr-3 |
| Tyr2 | 1 2 3a 3b 1’ 2′/6′ 3′/5′ 4′ NH | 169.9, C 54.2, CH 35.5, CH2 128.5, C 130.1, CH 115.5, CH 156.1, C | 4.31, m 3.11, m 2.55, m 6.89, d (8.4) 6.58, d (8.4) 8.46, d (8.8) | Ahp-NH Thr-2, Thr-3 | Tyr-2, Tyr-1′ Tyr-1′ Tyr-4′ Thr-1 |
| Ahp3 | 1 2 3 4 5 NH OH | 169.0, C 49.4, CH 22.0, CH2 21.9, CH2 74.1, CH | 3.62, m 2.41, m 1.64, m 5.06, brs 7.06, d (8.8) 5.99, d (3.1) | Ahp-NH Phe-3a, Phe-3b Tyr-2, Ahp-3 | Ahp-1, Ahp-3 Ahp-2 Ahp-1 Tyr-1 |
| Phe4 | 1 2 3a 3b 1′ 2′/6′ 3′/5′ 4′ | 170.8, C 50.7, CH 35.8, CH2 137.2, C 129.9, CH 128.2, CH 126.7, CH | 4.76, dd (7.1, 4.6) 2.88, t (12.9, 12.9) 1.81, dd (10.4, 3.9) 6.84, d (7.0) 7.19, t (7.3, 7.3) 7.14, d (7.0) | Ahp-5 | Phe-2, Phe-1′, Phe-2′/6′ Phe-3a, Phe-3b, Phe-4′ Phe-1′ Phe C2′/6′ |
| MeTyr5 | 1 2 3 1′ 2′/6′ 3′/5′ 4′ NCH3 OH | 169.4, C 61.3, CH 33.3, CH2 128.2, C 130.9, CH 115.5, CH 156.7, C 33.3, CH3 | 4.89, dd (8.9, 2.5) 3.10, m 7.00, d (8.4) 6.78, d (8.4) 2.71, s 9.33, s | Val-NH | MeTyr-1′ MeTyr-1′, MeTyr-3′/5′ MeTyr-2′/6′, MeTyr-4′ MeTyr-2, Phe-1 MeTyr-3′/5′, MeTyr-4′ |
| Val6 | 1 2 3 4 5 NH | 172.4, C 56.3, CH 31.4, CH 19.7, CH3 17.7, CH3 | 4.63, dd (4.9, 4.6) 2.02, m 0.84, d (6.6) 0.71, d (6.6) 7.39, d (9.7) | MeTyr-2 | Val-2, Val-3, Val-5 Val-2, Val-3, Val-4 MeTyr-1 |
| Unit | Position | δC | δH (J in Hz) | ROESY | HMBC a |
|---|---|---|---|---|---|
| BA | 1 2 3 4 | 172.2, C 37.4, CH2 19.3, CH2 14.1, CH3 | 2.08, m 1.52, m 0.87, t (7.4, 7.4) | Asp-NH | BA-1, BA-3, BA-4 BA-1, BA-2, BA-4 BA-2 |
| Asp | 1 2 3a 3b 4 NH | 173.7, C 50.8, CH 40.1, CH2 175.3, C | 4.54, m 2.51, m 2.12, m 8.00, d (8.0) | Thr-NH | Asp-2, Asp-4 BA-1 |
| Thr1 | 1 2 3 4 NH | 169.9, C 55.0, CH 72.3, CH 17.9, CH3 | 4.58, d (9.0) 5.30, q (6.8, 6.5, 6.8) 1.15, d (6.5) 7.23, d (7.6) | Arg-NH Arg-NH Asp-2 | Thr-1 Thr-4, Val-1 Thr-2, Thr-3 |
| Arg2 | 1 2 3 4 5 6 NH | 170.4, C 49.0, CH 26.4, CH2 24.5, CH2 39.8, CH2 158.7, C | 4.15, m 1.88, m 1.46, m 2.95, m 8.5, d (8.7) | Ahp-NH Thr-2, Thr-3 | Thr-1 |
| Ahp3 | 1 2 3 4 5 NH OH | 169.6, C 48.6, CH 21.7, CH2 22.0, CH2 74.2, CH | 3.65, m 2.43, m 1.68, m 5.03, brs 6.95, d (9.7) 6.01, d (2.6) | Phe-3a, Phe-3b Arg-2, Ahp-3 | Ahp-1, Ahp-3 Ahp-1 Arg-1 |
| Phe4 | 1 2 3a 3b 1′ 2′/6′ 3′/5′ 4′ | 170.0, C 50.6, CH 35.6, CH2 137.1, C 129.8, CH 128.2, CH 126.7, CH | 4.74, dd (7.2, 3.5) 2.86, m 1.69, m 6.78, d (7.1) 7.17, t (7.1, 7.1) 7.13, d (7.8) | Ahp-5 | Phe-1, Phe-3a, Phe-3b Phe-1′ Phe-3a, Phe-3b, Phe-4′ Phe-1′ Phe-3′/5′ |
| MePhe5 | 1 2 3 1′ 2′/6′ 3′/5′ 4′ NCH3 | 169.4, C 60.9, CH 34.4, CH2 138.4, C 130.0, CH 129.1, CH 127.2, CH 35.6, CH3 | 5.02, m 3.23, m 7.24, d (7.6) 7.41, t (7.7, 7.7) 7.32, d (7.5) 2.84, s | Val-NH | MePhe-1′ MePhe-1′ MePhe-4′ MePhe-3′/5′ MePhe-2 |
| Val6 | 1 2 3 4 5 NH | 172.6, C 56.4, CH 31.6, CH 19.7, CH3 17.8, CH3 | 4.71, dd (5.2, 4.2) 2.05, m 0.88, d (6.9) 0.76, d (6.9) 7.41, d (7.7) | MePhe-2 | Val-3 Val-2, Val-3, Val-5 Val-2, Val-3, Val-4 MePhe-1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazur-Marzec, H.; Fidor, A.; Cegłowska, M.; Wieczerzak, E.; Kropidłowska, M.; Goua, M.; Macaskill, J.; Edwards, C. Cyanopeptolins with Trypsin and Chymotrypsin Inhibitory Activity from the Cyanobacterium Nostoc edaphicum CCNP1411. Mar. Drugs 2018, 16, 220. https://doi.org/10.3390/md16070220
Mazur-Marzec H, Fidor A, Cegłowska M, Wieczerzak E, Kropidłowska M, Goua M, Macaskill J, Edwards C. Cyanopeptolins with Trypsin and Chymotrypsin Inhibitory Activity from the Cyanobacterium Nostoc edaphicum CCNP1411. Marine Drugs. 2018; 16(7):220. https://doi.org/10.3390/md16070220
Chicago/Turabian StyleMazur-Marzec, Hanna, Anna Fidor, Marta Cegłowska, Ewa Wieczerzak, Magdalena Kropidłowska, Marie Goua, Jenny Macaskill, and Christine Edwards. 2018. "Cyanopeptolins with Trypsin and Chymotrypsin Inhibitory Activity from the Cyanobacterium Nostoc edaphicum CCNP1411" Marine Drugs 16, no. 7: 220. https://doi.org/10.3390/md16070220