Antihypertensive Effects of Two Novel Angiotensin I-Converting Enzyme (ACE) Inhibitory Peptides from Gracilariopsis lemaneiformis (Rhodophyta) in Spontaneously Hypertensive Rats (SHRs)

Abstract
1. Introduction
2. Results
2.1. Screening of the Active ACE Inhibitory Peptides.
2.2. Properties of Peptides
2.3. Stability Study for ACE Inhibitory Activity
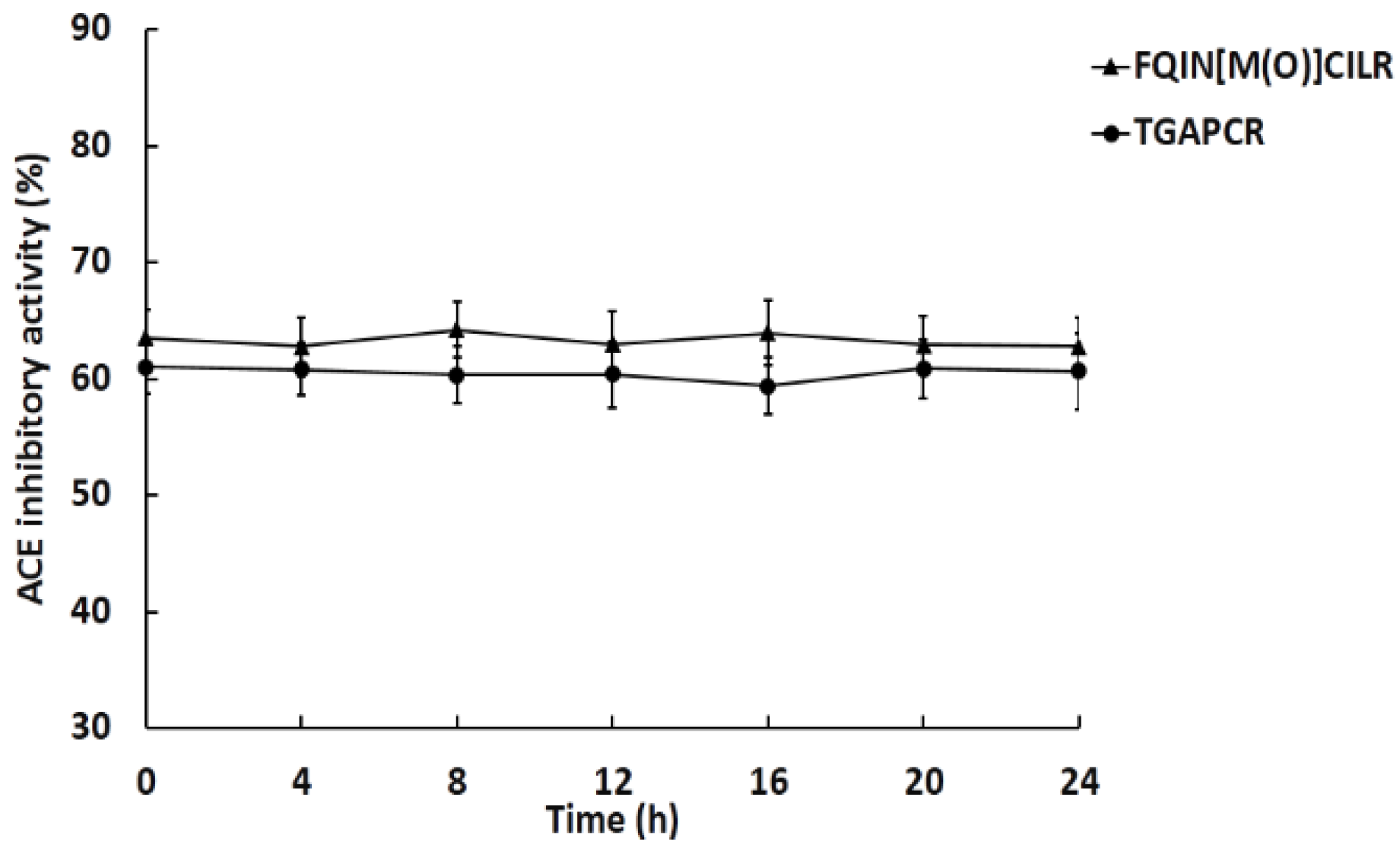
2.4. Stability of Peptides against ACE
2.5. Characterization of the Inhibition Pattern on ACE
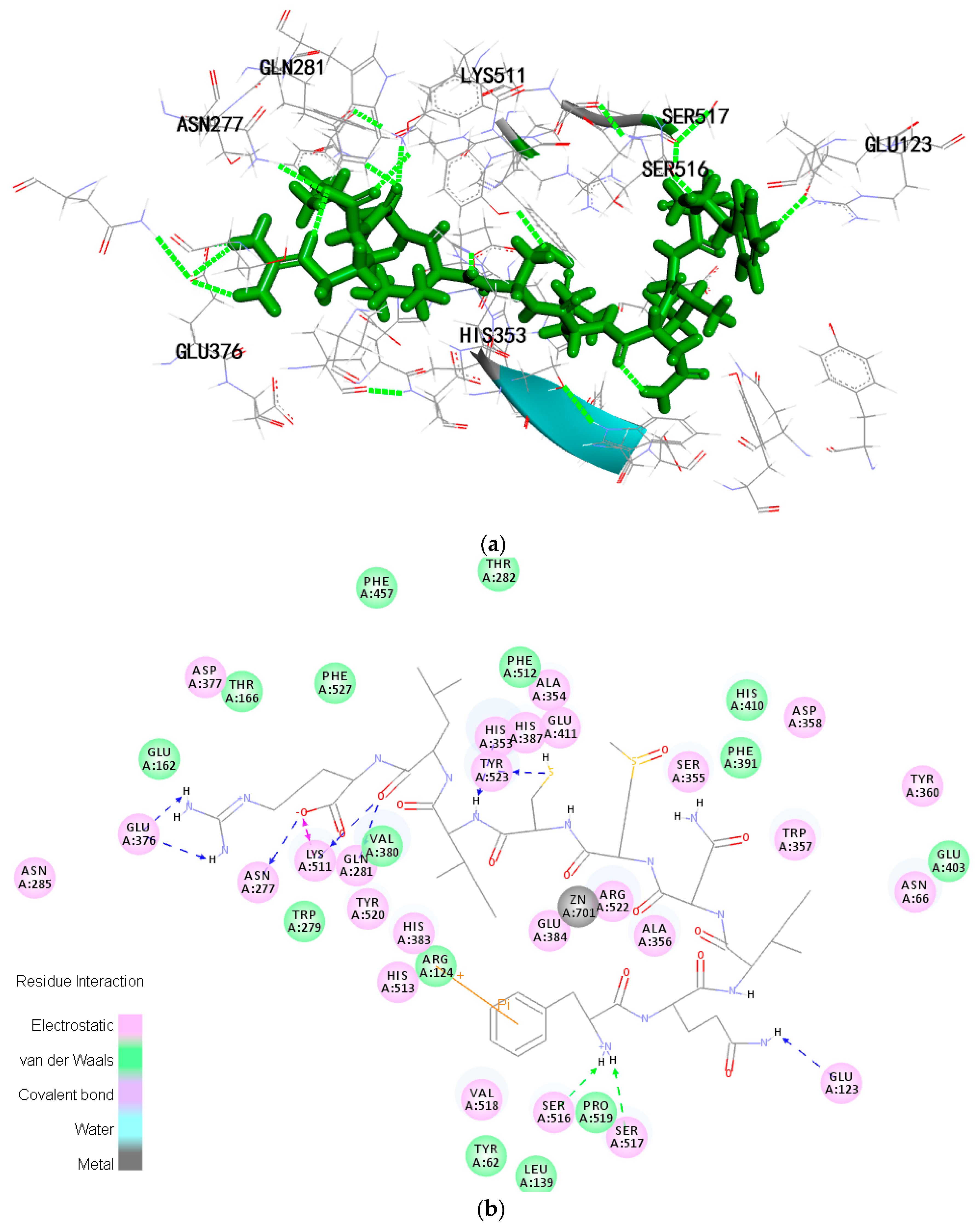
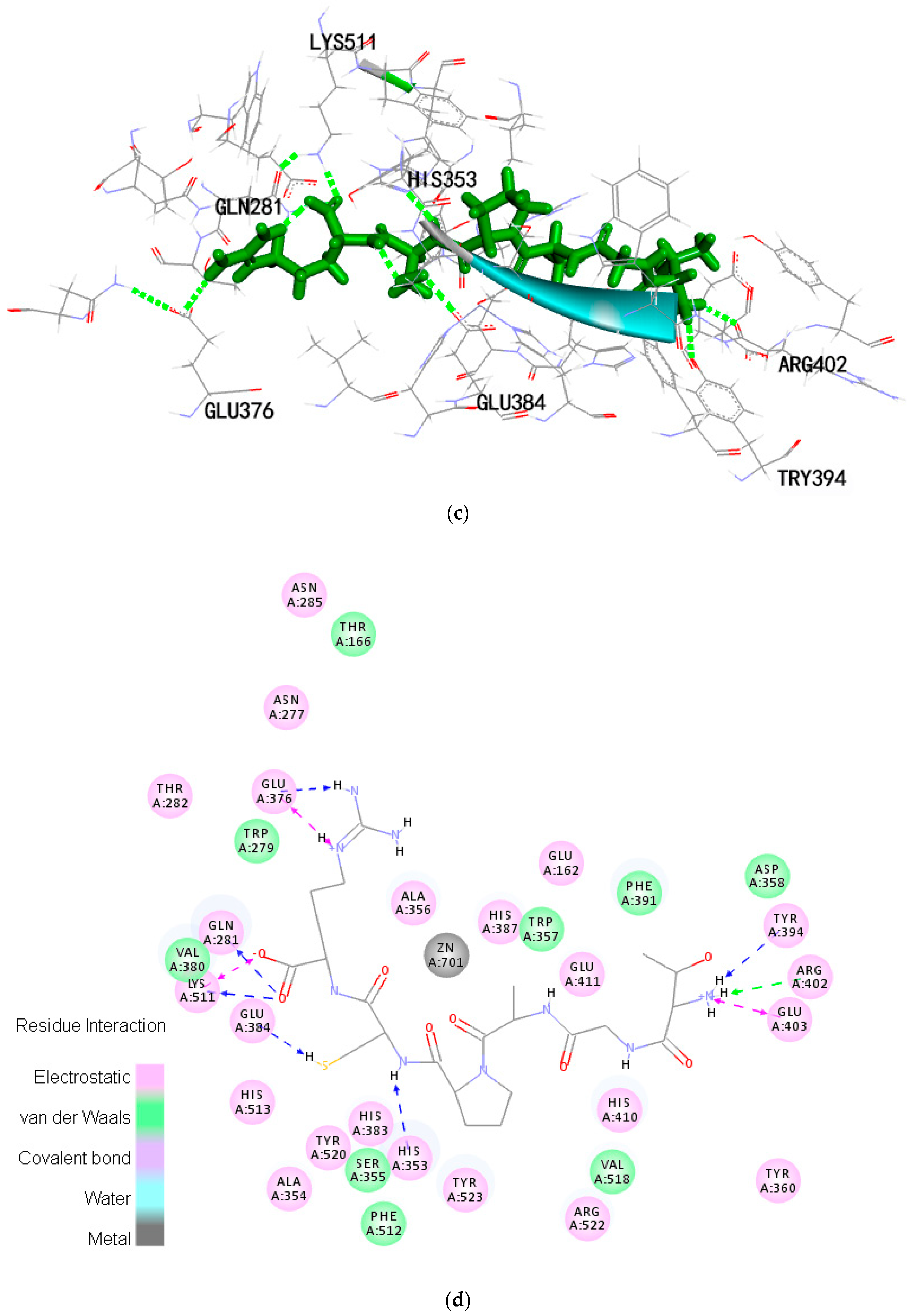
2.6. Molecular Docking
2.7. Antihypertensive Activity of the Two Peptides on SHRs
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Preparation of G. Lemaneiformis Hydrolysates
4.3. Identification of Peptides by LC-MS/MS
4.4. Synthesis of ACE Inhibitory Peptides
4.5. Measurement of ACE Inhibition Activity
4.6. Properties of Peptides
4.7. Stability Study for ACE Inhibitory Activity
4.7.1. Thermal Stability for Peptides
4.7.2. pH Stability for Peptides
4.7.3. Gastrointestinal Stability of Peptides
4.8. Stability of Peptides against ACE
4.9. Determination of Inhibitory Pattern
4.10. Molecular Docking
4.11. Antihypertensive Effect on SHRs
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cornélissen, G.; Delcourt, A.; Toussaint, G.; Otsuka, K.; Watanabe, Y.; Siegelova, J.; Fiser, B.; Dusek, J.; Homolka, P.; Singh, R.B.; et al. Opportunity of detecting pre-hypertension: worldwide data on blood pressure overswinging. Biomed. Pharmacother. 2005, 59, S152–S157. [Google Scholar] [CrossRef]
- Faria, M.; Costa, E.L.; Gontijo, J.A.; Netto, F.M. Evaluation of the hypotensive potential of bovine and porcine collagen hydrolysates. J. Méd. Food 2008, 11, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Natesh, R.; Schwager, S.L.U.; Sturrock, E.D.; Acharya, K.R. Crystal structure of the human angiotensin-converting enzyme–lisinopril complex. Nature 2003, 421, 551. [Google Scholar] [CrossRef] [PubMed]
- Pacurari, M.; Kafoury, R.; Tchounwou, P.B.; Ndebele, K. The Renin-Angiotensin-aldosterone system in vascular inflammation and remodeling. Int. J. Inflamm. 2014, 2014, 689360. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.C.; Ke, W.H.; Ceng, L.H.; Hsieh, C.W.; Wung, B.S. Calcium- and phosphatidylinositol 3-kinase/Akt-dependent activation of endothelial nitric oxide synthase by apigenin. Life Sci. 2010, 87, 743–749. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.; Hartl, F.U. Molecular chaperones in cellular protein folding. Bioessays 1994, 16, 689–692. [Google Scholar] [CrossRef] [PubMed]
- Fujita, H.; Yokoyama, K.; Yoshikawa, M. Classification and antihypertensive activity of angiotensin I-converting enzyme inhibitory peptides derived from food proteins. J. Food Sci. 2000, 65, 564–569. [Google Scholar]
- Alemán, A.; Giménez, B.; Pérez-Santin, E.; Gómez-Guillén, M.C.; Montero, P. Contribution of Leu and Hyp residues to antioxidant and ACE-inhibitory activities of peptide sequences isolated from squid gelatin hydrolysate. Food Chem. 2011, 125, 334–341. [Google Scholar] [CrossRef]
- García-Mora, P.; Martín-Martinez, M.; Bonache, M.A.; Gonzalez-Muniz, M.; Peñas, E.; Frias, J.; Martínez-Villaluenga, C. Identification, functional gastrointestinal stability and molecular docking studies of lentil peptides with dual antioxidant and angiotensin I converting enzyme inhibitory activities. Food Chem. 2017, 221, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Aluko, R.E. Structure and function of plant protein-derived antihypertensive peptides. Curr. Opin. Food Sci. 2015, 4, 44–50. [Google Scholar] [CrossRef]
- Wijesekara, I.; Kim, S. Angiotensin-I-converting enzyme (ACE) inhibitors from marine resources: Prospects in the pharmaceutical industry. Mar. Drugs 2010, 8, 1080–1093. [Google Scholar] [CrossRef] [PubMed]
- Aluko, R.E. Antihypertensive peptides from food proteins. Annu. Rev. Food Sci. Technol. 2015, 6, 235–262. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Zou, D.; Zhu, M.; Yang, Y. Effects of CO2 levels and light intensities on growth and amino acid contents in red seaweed Gracilaria lemaneiformis. Aquac. Res. 2016, 48, 2683–2690. [Google Scholar] [CrossRef]
- Fan, Y.; Wang, W.; Song, W.; Chen, H.; Teng, A.; Liu, A. Partial characterization and anti-tumor activity of an acidic polysaccharide from Gracilaria lemaneiformis. Carbohydr. Polym. 2012, 88, 1313–1318. [Google Scholar] [CrossRef]
- Li, P.; Ying, J.; Chang, Q.; Zhu, W.; Yang, G.; Xu, T.; Yi, H.; Pan, R.; Zhang, E.; Zeng, X.; et al. Effects of phycoerythrin from Gracilaria lemaneiformis in proliferation and apoptosis of SW480 cells. Oncol. Rep. 2016, 36, 3536–3544. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Liu, H.; Hou, Y.; Chan, Z.; Di, W.; Li, L.; Zeng, R. Preparation, characterization and alcoholic liver injury protective effects of algal oligosaccharides from Gracilaria lemaneiformis. Food Res. Int. 2017, 100, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Zheng, G.; You, L.; Wen, L.; Li, C.; Fu, X.; Zhou, L. Structural characterization and macrophage immunomodulatory activity of a polysaccharide isolated from Gracilaria lemaneiformis. J. Funct. Foods 2017, 33, 286–296. [Google Scholar] [CrossRef]
- Wen, L.; Zhang, Y.; Sun-Waterhouse, D.; You, L.; Fu, X. Advantages of the polysaccharides from Gracilaria lemaneiformis over metformin in antidiabetic effects on streptozotocin-induced diabetic mice. RSC Adv. 2017, 7, 9141–9151. [Google Scholar] [CrossRef]
- Yuan, S.; Duan, Z.; Lu, Y.; Ma, X.; Wang, S. Optimization of decolorization process in agar production from Gracilaria lemaneiformis and evaluation of antioxidant activities of the extract rich in natural pigments. Biotech. 2018, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, L.; Bondu, S.; Doiron, K.; Rioux, L.; Turgeon, S.L. Characterization of antibacterial activity from protein hydrolysates of the macroalga Saccharina longicruris and identification of peptides implied in bioactivity. J. Funct. Foods 2015, 17, 685–697. [Google Scholar] [CrossRef]
- Beaulieu, L.; Sirois, M.; Tamigneaux, É. Evaluation of the in vitro biological activity of protein hydrolysates of the edible red alga, Palmaria palmata (dulse) harvested from the Gaspe coast and cultivated in tanks. J. Appl. Phycol. 2016, 28, 3101–3115. [Google Scholar] [CrossRef]
- Cian, R.E.; Garzón, A.G.; Ancona, D.B.; Guerrero, L.C.; Drago, S.R. Hydrolyzates from Pyropia columbina seaweed have antiplatelet aggregation, antioxidant and ACE I inhibitory peptides which maintain bioactivity after simulated gastrointestinal digestion. LWT Food Sci. Technol. 2015, 64, 881–888. [Google Scholar] [CrossRef]
- Pan, S.; Wang, S.; Jing, L.; Yao, D. Purification and characterisation of a novel angiotensin-I converting enzyme (ACE)-inhibitory peptide derived from the enzymatic hydrolysate of Enteromorpha clathrata protein. Food Chem. 2016, 211, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Paiva, L.; Lima, E.; Neto, A.I.; Baptista, J. Isolation and characterization of angiotensin I-converting enzyme (ACE) inhibitory peptides from Ulva rigida C. Agardh protein hydrolysate. J. Funct. Foods 2016, 26, 65–76. [Google Scholar] [CrossRef]
- Xie, J.; Chen, X.; Wu, J.; Zhang, Y.; Zhou, Y.; Zhang, L.; Tang, Y.; Wei, D. Antihypertensive Effects, Molecular Docking Study, and Isothermal Titration Calorimetry Assay of Angiotensin I-Converting Enzyme Inhibitory Peptides from Chlorella vulgaris. J. Agric. Food Chem. 2018, 66, 1359–1368. [Google Scholar] [CrossRef] [PubMed]
- Qu, W.; Ma, H.; Li, W.; Pan, Z.; Owusu, J.; Venkitasamy, C. Performance of coupled enzymatic hydrolysis and membrane separation bioreactor for antihypertensive peptides production from Porphyra yezoensis protein. Process Biochem. 2015, 50, 245–252. [Google Scholar] [CrossRef]
- Li, H.; Aluko, R.E. Kinetics of the inhibition of calcium/calmodulin-dependent protein kinase II by pea protein-derived peptides. J. Nutr. Biochem. 2005, 16, 656–662. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Jia, J.; Yan, H.; Du, J.; Gui, Z. A novel angiotensin-capital I, Ukrainian converting enzyme (ACE) inhibitory peptide from gastrointestinal protease hydrolysate of silkworm pupa (Bombyx mori) protein: Biochemical characterization and molecular docking study. Peptides 2015, 68, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Rohit, A.C.; Sathisha, K.; Aparna, H.S. A variant peptide of buffalo colostrum beta-lactoglobulin inhibits angiotensin I-converting enzyme activity. Eur. J. Med. Chem. 2012, 53, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Furuta, T.; Miyabe, Y.; Yasui, H.; Kinoshita, Y.; Kishimura, H. Angiotensin I Converting Enzyme Inhibitory Peptides Derived from Phycobiliproteins of Dulse Palmaria palmata. Mar. Drugs 2016, 14, 32. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Lv, X.; Xu, X.; Yu, H.; Sun, X.; Xu, N. Purification and identification of a novel ACE inhibitory peptide from marine alga Gracilariopsis lemaneiformis protein hydrolysate. Eur. Food Res. Technol. 2017, 243, 1829–1837. [Google Scholar] [CrossRef]
- Guo, M.; Chen, X.; Wu, Y.; Zhang, L.; Huang, W.; Yuan, Y.; Fang, M.; Xie, J.; Wei, D. Angiotensin I-converting enzyme inhibitory peptides from Sipuncula (Phascolosoma esculenta): Purification, identification, molecular docking and antihypertensive effects on spontaneously hypertensive rats. Process Biochem. 2017, 63, 84–95. [Google Scholar] [CrossRef]
- Maruyama, S.; Mitachi, H.; Awaya, J.; Kurono, M.; Tomizuka, N.; Suzuki, H. Angiotensin I-Converting Enzyme Inhibitory Activity of the C-Terminal Hexapeptide of αs1-Casein. Agric. Biol. Chem. Tokyo 1987, 51, 2557–2561. [Google Scholar] [CrossRef]
- Escudero, E.; Mora, L.; Toldrá, F. Stability of ACE inhibitory ham peptides against heat treatment and in vitro digestion. Food Chem. 2014, 161, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Tagliazucchi, D.; Martini, S.; Shamsia, S.; Helal, A.; Conte, A. Biological activities and peptidomic profile of invitro-digested cow, camel, goat and sheep milk. Int. Dairy J. 2018, 81, 19–27. [Google Scholar] [CrossRef]
- Maeno, M.; Yamamoto, N.; Takano, T. Identification of an antihypertensive peptide from casein hydrolysate produced by a proteinase from Lactobacillus helveticus CP790. J. Dairy Sci. 1996, 79, 1316–1321. [Google Scholar] [CrossRef]
- Kuba, M.; Tanaka, K.; Tawata, S.; Takeda, Y.; Yasuda, M. Angiotensin I-converting enzyme inhibitory peptides isolated from tofuyo fermented soybean food. Biosci. Biotechnol. Biochem. 2003, 67, 1278–1283. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Fang, L.; Min, W.; Liu, J.; Li, H. Exploration of the molecular interactions between angiotensin-I-converting enzyme (ACE) and the inhibitory peptides derived from hazelnut (Corylus heterophylla Fisch.). Food Chem. 2018, 245, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Balti, R.; Bougatef, A.; Sila, A.; Guillochon, D.; Dhulster, P.; Nedjar-Arroume, N. Nine novel angiotensin I-converting enzyme (ACE) inhibitory peptides from cuttlefish (Sepia officinalis) muscle protein hydrolysates and antihypertensive effect of the potent active peptide in spontaneously hypertensive rats. Food Chem. 2015, 170, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.T.H.; Dedachi, K.; Matsui, T.; Kurita, N.; Borgatti, M.; Gambari, R.; Sylte, I. Dipeptide Inhibitors of Thermolysin and Angiotensin I-Converting Enzyme. Curr. Top. Med. Chem. 2012, 12, 1748–1762. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Du, J.; Jia, J.; Kuang, C. Production of ACE inhibitory peptides from sweet sorghum grain protein using alcalase: Hydrolysis kinetic, purification and molecular docking study. Food Chem. 2016, 199, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Kuramoto, S.; Kaneyoshi, G.; Morinaga, Y.; Matsue, H.; Iwai, K. Angiotensin-Converting enzyme-Inhibitory peptides isolated from pepsin hydrolyzate of apios americana tuber and their hypotensive effects in spontaneously hypertensive rats. Food Sci. Technol. Res. 2013, 19, 399–407. [Google Scholar] [CrossRef]
- Duan, X.; Wu, F.; Li, M.; Yang, N.; Wu, C.; Jin, Y.; Yang, J.; Jin, Z.; Xu, X. Naturally occurring angiotensin I-Converting enzyme inhibitory peptide from a fertilized egg and its inhibitory mechanism. J. Agric. Food Chem. 2014, 62, 5500–5506. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, M.; Jia, A.; Zhang, Y.; Zhu, H.; Zhang, C.; Sun, Z.; Liu, C. Purification and characterization of angiotensin I converting enzyme inhibitory peptides from jellyfish Rhopilema esculentum. Food Res. Int. 2013, 50, 339–343. [Google Scholar] [CrossRef]
- Lin, K.; Zhang, L.; Han, X.; Meng, Z.; Wu, Y.; Zhang, J.; Cheng, D. Quantitative Structure-Activity Relationship Modeling Coupled with Molecular Docking Analysis in Screening of Angiotensin I-Converting Enzyme Inhibitory Peptides from Qula Casein Hydrolysates Obtained by Two-Enzyme Combination Hydrolysis. J. Agric. Food Chem. 2018, 66, 3221–3228. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; He, H.; Chen, X.; Sun, C.; Zhang, Y.; Zhou, B. Purification and identification of novel angiotensin-I-converting enzyme inhibitory peptides from shark meat hydrolysate. Process Biochem. 2008, 43, 457–461. [Google Scholar] [CrossRef]
- Marambe, H.K.; Shand, P.J.; Wanasundara, J.P.D. Release of angiotensin I-converting enzyme inhibitory peptides from flaxseed (Linum usitatissimum L.) protein under simulated gastrointestinal digestion. J. Agric. Food Chem. 2011, 57, 9596–9604. [Google Scholar] [CrossRef] [PubMed]
- Salampessy, J.; Reddy, N.; Phillips, M.; Kailasapathy, K. Isolation and characterization of nutraceutically potential ACE-Inhibitory peptides from leatherjacket (Meuchenia sp.) protein hydrolysates. LWT-Food. Sci. Technol. 2017, 80, 430–436. [Google Scholar] [CrossRef]
- Fujita, H.; Yoshikawa, M. LKPNM: A prodrug-type ACE-inhibitory peptide derived from fish protein. Immunopharmacology 1999, 44, 123–127. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptides | Number of Residues | Molecular Weight (Da) | ToxiPed | Solubility in Water | Iso-Electric Point (PI) |
|---|---|---|---|---|---|
| FQIN [M(O)] CILR | 9 | 1153.43 | non-toxin | poor | 8.61 |
| TGAPCR | 6 | 603.69 | non-toxin | good | 8.55 |
| Kinetics Parameters | Control | FQIN [M(O)] CILR | TGAPCR | ||
|---|---|---|---|---|---|
| 1 mg/mL | 0.5 mg/mL | 1 mg/mL | 0.5 mg/mL | ||
| Km (mM) | 2.66 ± 0.21 | 2.66 ± 0.21 | 2.66 ± 021 | ||
| Vmax (mg−1·mL·min) | 2.23 ± 0.67 | 0.10 ± 0.05 | 0.14 ± 0.02 | 0.11 ± 0.03 | 0.17 ± 0.02 |
| Ki (mM) | 0.71 ± 0.04 | 0.86 ± 0.06 | |||
| Peptides | -CDOCKER ENERGY (kcal·mol−1) | -CDOCKER INTERACTION ENERGY (kcal·mol−1) |
|---|---|---|
| FQIN [M(O)] CILR | 158.117 | 145.01 |
| TGAPCR | 90.4226 | 105.509 |
| Peptides | Donor Atom | Acceptor Atom | Distance (Å) | Active Pocket |
|---|---|---|---|---|
| FQIN [M(O)] CILR | A: ASN277: HD22 | F: O125 | 2.39 | |
| A: GLN281: HE22 | F: O89 | 1.97 | S2 | |
| A: LYS511: HZ1 | F: O89 | 2.02 | S2 | |
| A: LYS511: HZ3 | F: O126 | 2.31 | S2 | |
| A: TYR523: HH | F: S63 | 2.24 | S1 | |
| F: H145 | A: SER517: O | 1.95 | ||
| F: H146 | A: SER516: O | 1.98 | ||
| F: H148 | A: GLU123: OE1 | 2.14 | ||
| F: H158 | A: HIS353: NE2 | 2.33 | S2 | |
| F: H161 | A: GLU376: OE1 | 2.44 | ||
| F: H161 | A: GLU: 376: OE2 | 2.03 | ||
| F: H163 | A: GLU: 376: OE1 | 2.32 | ||
| TGAPCR | A: GLN281: HE22 | T: O2 | 2.06 | S2 |
| A: LYS511: HZ1 | T: O2 | 1.78 | S2 | |
| T: H71 | A: GLU376: OE2 | 2.16 | ||
| T: H75 | A: GLU384: OE2 | 2.26 | S1 | |
| T: H76 | A: HIS353: NE2 | 2.24 | S2 | |
| T: H78 | A: TYR394: OH | 2.32 | ||
| T: H79 | A: ARG402: O | 2.41 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, Z.; Liu, Y.; Wang, J.; Wu, S.; Geng, L.; Sui, Z.; Zhang, Q. Antihypertensive Effects of Two Novel Angiotensin I-Converting Enzyme (ACE) Inhibitory Peptides from Gracilariopsis lemaneiformis (Rhodophyta) in Spontaneously Hypertensive Rats (SHRs). Mar. Drugs 2018, 16, 299. https://doi.org/10.3390/md16090299
Deng Z, Liu Y, Wang J, Wu S, Geng L, Sui Z, Zhang Q. Antihypertensive Effects of Two Novel Angiotensin I-Converting Enzyme (ACE) Inhibitory Peptides from Gracilariopsis lemaneiformis (Rhodophyta) in Spontaneously Hypertensive Rats (SHRs). Marine Drugs. 2018; 16(9):299. https://doi.org/10.3390/md16090299
Chicago/Turabian StyleDeng, Zhenzhen, Yingjuan Liu, Jing Wang, Suhuang Wu, Lihua Geng, Zhenghong Sui, and Quanbin Zhang. 2018. "Antihypertensive Effects of Two Novel Angiotensin I-Converting Enzyme (ACE) Inhibitory Peptides from Gracilariopsis lemaneiformis (Rhodophyta) in Spontaneously Hypertensive Rats (SHRs)" Marine Drugs 16, no. 9: 299. https://doi.org/10.3390/md16090299
APA StyleDeng, Z., Liu, Y., Wang, J., Wu, S., Geng, L., Sui, Z., & Zhang, Q. (2018). Antihypertensive Effects of Two Novel Angiotensin I-Converting Enzyme (ACE) Inhibitory Peptides from Gracilariopsis lemaneiformis (Rhodophyta) in Spontaneously Hypertensive Rats (SHRs). Marine Drugs, 16(9), 299. https://doi.org/10.3390/md16090299