Discovery of an Unusual Fatty Acid Amide from the ndgRyo Gene Mutant of Marine-Derived Streptomyces youssoufiensis

 , ,
, ,  and
and
Abstract
:
1. Introduction
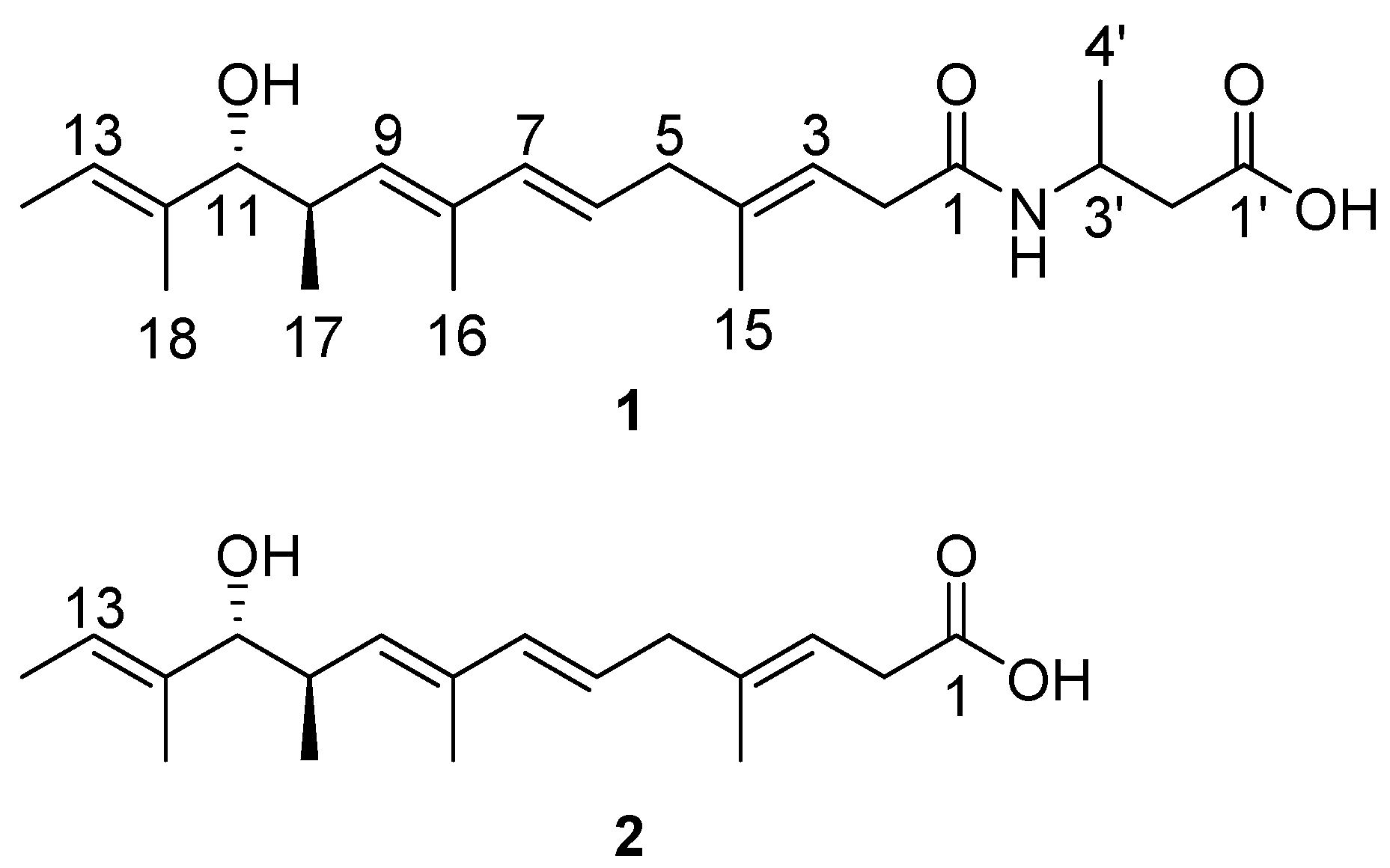
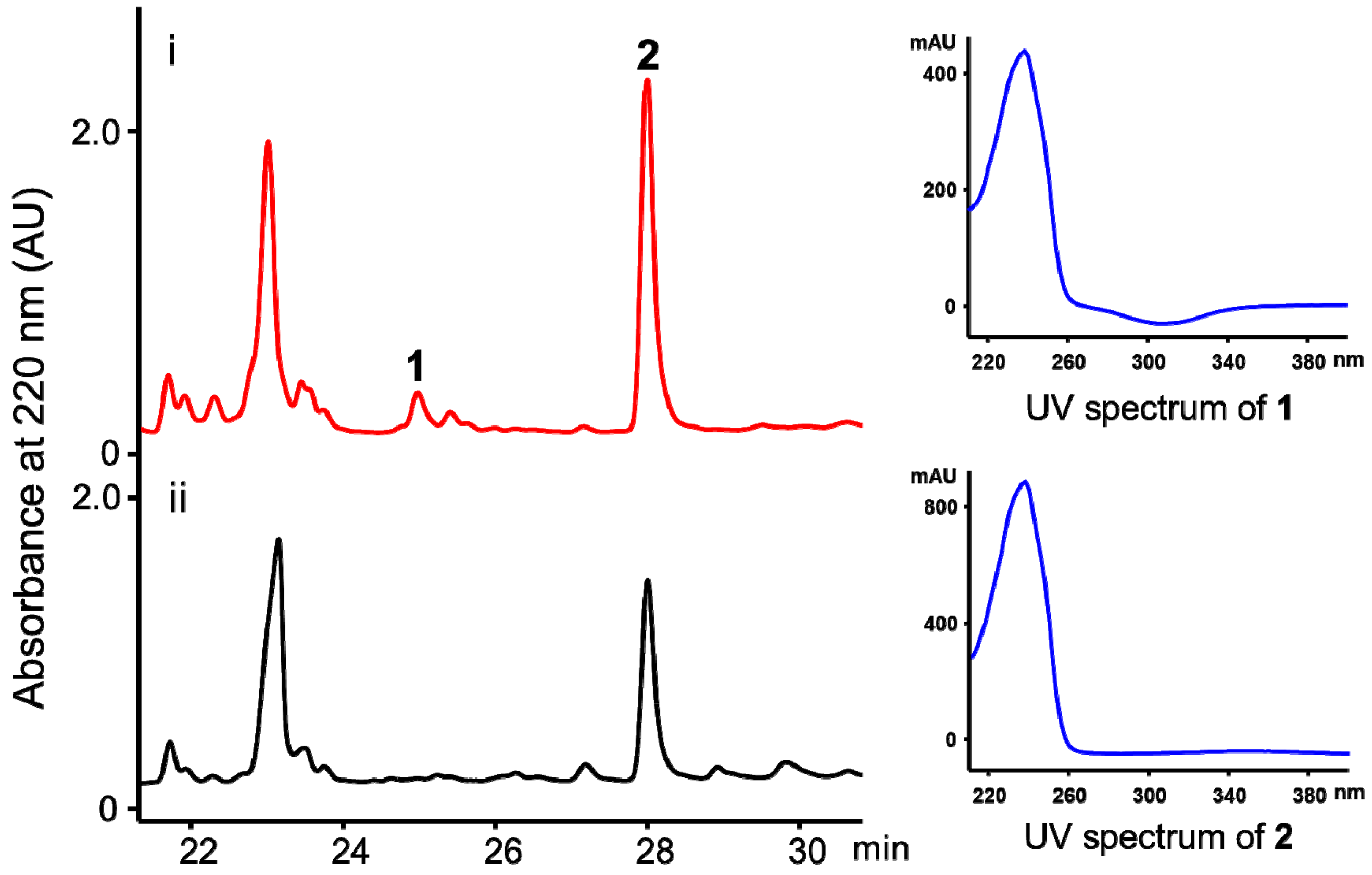
2. Results and Discussion
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Bacterial Strains and Culture Conditions
3.3. DNA Isolation and Manipulation
3.4. Gene Inactivation
3.5. Isolation and Purification of the Compounds
3.6. Nucleotide Sequence Accession Number
4. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Manivasagan, P.; Venkatesan, J.; Sivakumar, K.; Kim, S.K. Pharmaceutically active secondary metabolites of marine actinobacteria. Microbiol. Res. 2014, 169, 262–278. [Google Scholar] [CrossRef] [PubMed]
- Rutledge, P.J.; Challis, G.L. Discovery of microbial natural products by activation of silent biosynthetic gene clusters. Nat. Rev. Microbiol. 2015, 13, 509–523. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Liao, G.; Zhang, J.; Tan, H. Identification of novel tylosin analogues generated by a wblA disruption mutant of Streptomyces ansochromogenes. Microb. Cell Fact. 2015, 14, 173. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tan, H. Biosynthesis and molecular regulation of secondary metabolites in microorganisms. Sci. China Life Sci. 2017, 60, 935–938. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Chater, K.F.; Chandra, G.; Niu, G.; Tan, H. Molecular regulation of antibiotic biosynthesis in Streptomyces. Microbiol. Mol. Biol. Rev. 2013, 77, 112–143. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.N.; Jeong, Y.; Yoo, J.S.; Roe, J.H.; Cho, B.K.; Kim, B.G. Genome-scale analysis reveals a role for NdgR in the thiol oxidative stress response in Streptomyces coelicolor. BMC Genom. 2015, 16, 116. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.H.; Song, E.; Kim, E.J.; Lee, K.; Kim, W.S.; Park, S.S.; Hahn, J.S.; Kim, B.J. NdgR, an IclR-like regulator involved in amino-acid-dependent growth, quorum sensing, and antibiotic production in Streptomyces coelicolor. Appl. Microbiol. Biotechnol. 2009, 82, 501–511. [Google Scholar] [CrossRef] [PubMed]
- Santamarta, I.; Lópezgarcía, M.T.; Pérezredondo, R.; Koekman, B.; Martín, J.F.; Liras, P. Connecting primary and secondary metabolism: AreB, an IclR-like protein, binds the AREccaR sequence of S. clavuligerus and modulates leucine biosynthesis and cephamycin C and clavulanic acid production. Mol. Microbiol. 2007, 66, 511–524. [Google Scholar] [CrossRef] [PubMed]
- Che, Q.; Li, T.; Liu, X.; Yao, T.; Li, J.; Gu, Q.; Li, D.; Li, W.; Zhu, T. Genome scanning inspired isolation of reedsmycins A-F, polyene-polyol macrolides from Streptomyces sp. CHQ-64. RSC Adv. 2015, 5, 22777–22782. [Google Scholar] [CrossRef]
- Yao, T.; Liu, Z.; Li, T.; Zhang, H.; Liu, J.; Li, H.; Che, Q.; Zhu, T.; Li, D.; Li, W. Characterization of the biosynthetic gene cluster of the polyene macrolide antibiotic reedsmycins from a marine-derived Streptomyces strain. Microb. Cell Fact. 2018, 17, 98. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Liu, Z.; Zhang, Z.; Zhang, X.; Zhu, T.; Gu, Q.; Li, W.; Che, Q.; Li, D. Geranylpyrrol A and piericidin F from Streptomyces sp. CHQ-64 ΔrdmF. J. Nat. Prod. 2017, 80, 1684–1687. [Google Scholar] [CrossRef] [PubMed]
- Dailey, O.D., Jr.; Wang, X.; Chen, F.; Huang, G. Anticancer activity of branched-chain derivatives of oleic acid. Anticancer Res. 2011, 31, 3165–3169. [Google Scholar] [PubMed]
- Jain, M.K.; Ghomashchi, F.; Yu, B.Z.; Bayburt, T.; Murphy, D.; Houck, D.; Solowiej, J.E. Fatty acid amides: scooting mode-based discovery of tight-binding competitive inhibitors of secreted phospholipases A2. J. Med. Chem. 1992, 35, 3584–3586. [Google Scholar] [CrossRef] [PubMed]
- Blancaflor, E.B.; Kilaru, A.; Keereetaweep, J.; Khan, B.R.; Faure, L.; Chapman, K.D. N-Acylethanolamines: lipid metabolites with functions in plant growth and development. Plant J. 2014, 79, 568–583. [Google Scholar] [CrossRef] [PubMed]
- Tuo, W.; Leleu-Chavain, N.; Spencer, J.; Sansook, S.; Millet, R.; Chavatte, P. Therapeutic potential of fatty acid amide hydrolase, monoacylglycerol lipase, and N-acylethanolamine acid amidase inhibitors. J. Med. Chem. 2016, 60, 4–46. [Google Scholar] [CrossRef] [PubMed]
- Takaishi, M.; Kudo, F.; Eguchi, T. A unique pathway for the 3-aminobutyrate starter unit from L-glutamate through β-Glutamate during biosynthesis of the 24-membered macrolactam antibiotic, incednine. Org. Lett. 2012, 14, 4591–4593. [Google Scholar] [CrossRef] [PubMed]
- Maniatis, T.; Fritsch, E.F.; Sambrook, J. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor: New York, NY, USA, 1982. [Google Scholar]
- Paget, M.S.; Chamberlin, L.; Atrih, A.; Foster, S.J.; Buttner, M.J. Evidence that the extracytoplasmic function sigma factor σE is required for normal cell wall structure in Streptomyces coelicolor A3(2). J. Bacteriol. 1999, 181, 204–211. [Google Scholar] [PubMed]
- Gust, B.; Challis, G.L.; Fowler, K.; Kieser, T.; Chater, K.F. PCR-targeted Streptomyces gene replacement identifies a protein domain needed for biosynthesis of the sesquiterpene soil odor geosmin. Proc. Natl. Acad. Sci. USA 2003, 100, 1541–1546. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, T.; Liu, J.; Yao, T.; Zhang, H.; Xia, J.; Li, H.; Che, Q.; Li, W. Development of the genetic system of mangrove derived Streptomyces sp. OUC6819. Chin. J. Mar. Drugs. 2016, 35, 53–59. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | 1 | 2 | ||
|---|---|---|---|---|
| δH (J in HZ) | δC | δH (J in HZ) | δC | |
| 1 | 172.4 | 174.9 | ||
| 2 | 2.95 (2H, d, 7.2) | 35.0 | 3.06 (2H, d, 7.2) | 33.0 |
| 3 | 5.36 (1H, t, 6.6) | 117.3 | 5.39 (1H, t, 6.6) | 116.7 |
| 4 | 138.4 | 137.7 | ||
| 5 | 2.82 (2H, d, 7.2) | 42.5 | 2.82 (2H, d, 7.2) | 42.5 |
| 6 | 5.57 (1H, dt, 15.6, 7.2) | 124.4 | 5.56 (1H, dt, 15.6, 7.2) | 124.4 |
| 7 | 6.12 (1H, d, 15.6) | 136.7 | 6.13 (1H, d, 15.6) | 136.7 |
| 8 | 133.7 | 133.7 | ||
| 9 | 5.33 (1H, d, 9.0) | 134.4 | 5.33 (1H, d, 9.6) | 134.4 |
| 10 | 2.71 (1H, m) | 36.1 | 2.71 (1H, m) | 36.1 |
| 11 | 3.72 (1H, d, 8.4) | 82.3 | 3.72 (1H, d, 7.8) | 82.3 |
| 12 | 136.7 | 136.7 | ||
| 13 | 5.46 (1H, q, 6.6) | 121.4 | 5.47(1H, q, 6.0) | 121.4 |
| 14 | 1.64 (3H, d, 6.6) | 11.6 | 1.64 (3H, d, 6.0) | 11.7 |
| 15 | 1.66 (3H, s) | 15.0 | 1.66 (3H, s) | 15.0 |
| 16 | 1.78 (3H, m) | 11.6 | 1.78 (3H, m) | 11.7 |
| 17 | 0.83 (3H, d, 6.6) | 16.6 | 0.83 (3H, d, 7.2) | 16.7 |
| 18 | 1.63 (3H, s) | 9.7 | 1.63 (3H, s) | 9.7 |
| 1′ | 173.2 | |||
| 2′ | 2.54 (1H, dd, 15.6, 6.0) 2.42 (1H, dd, 15.6, 6.0) | 40.0 | ||
| 3′ | 4.26 (1H, m) | 42.2 | ||
| 4′ | 1.21 (3H, d, 6.6) | 18.8 | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, J.; Liu, J.; Yang, L.; Liu, Z.; Li, H.; Che, Q.; Zhu, T.; Li, D.; Li, W. Discovery of an Unusual Fatty Acid Amide from the ndgRyo Gene Mutant of Marine-Derived Streptomyces youssoufiensis. Mar. Drugs 2019, 17, 12. https://doi.org/10.3390/md17010012
Hou J, Liu J, Yang L, Liu Z, Li H, Che Q, Zhu T, Li D, Li W. Discovery of an Unusual Fatty Acid Amide from the ndgRyo Gene Mutant of Marine-Derived Streptomyces youssoufiensis. Marine Drugs. 2019; 17(1):12. https://doi.org/10.3390/md17010012
Chicago/Turabian StyleHou, Jing, Jing Liu, Lu Yang, Zengzhi Liu, Huayue Li, Qian Che, Tianjiao Zhu, Dehai Li, and Wenli Li. 2019. "Discovery of an Unusual Fatty Acid Amide from the ndgRyo Gene Mutant of Marine-Derived Streptomyces youssoufiensis" Marine Drugs 17, no. 1: 12. https://doi.org/10.3390/md17010012
APA StyleHou, J., Liu, J., Yang, L., Liu, Z., Li, H., Che, Q., Zhu, T., Li, D., & Li, W. (2019). Discovery of an Unusual Fatty Acid Amide from the ndgRyo Gene Mutant of Marine-Derived Streptomyces youssoufiensis. Marine Drugs, 17(1), 12. https://doi.org/10.3390/md17010012