An Unprecedented Medium-Chain Diunsaturated N-acylhomoserine Lactone from Marine Roseobacter Group Bacteria


Abstract
:1. Introduction
2. Results
2.1. Occurrence of N-acylhomoserine Lactones in Roseobacter Group Bacteria of Fucus Spiralis and the Eastern North Sea
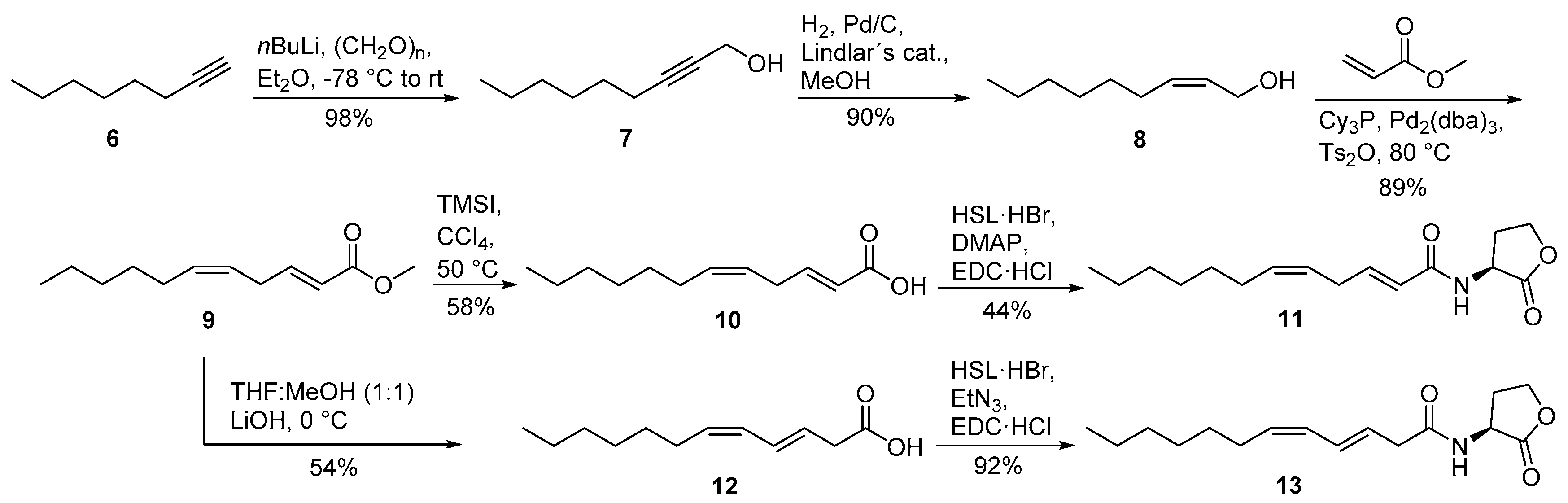
2.2. Identification and Synthesis of New Diunsaturated N-acylhomoserine Lactones from the Roseobacter Group
2.3. Activity of N-acylhomoserine Lactones in AHL Reporter Assays
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Strains and Culture Conditions
4.3. Bacteria Cultivation and XAD Extraction
4.4. AHL Reporter Assays
4.5. Synthetic Procedures
4.5.1. l-Homoserine Lactone Hydrobromide
4.5.2. Ethyl (2E,4E)-2,4-Dodecadienoate (3)
4.5.3. (2E,4E)-2,4-Dodecadienoic Acid (4)
4.5.4. N-((2E,4E)-2,4-Dodecadienoyl)homoserine Lactone (5)
4.5.5. Non-2-yn-1-ol (7)
4.5.6. Non-2-en-1-ol (8)
4.5.7. Methyl (2E,5Z)-2,5-Dodecadienoate (9)
4.5.8. (2E,5Z)-2,5-Dodecadienoic Acid (10)
4.5.9. N-((2E,5Z)-2,5-Dodecadienoyl)homoserine Lactone (11)
4.5.10. (3E,5Z)-3,5-Dodecadienoic Acid (12)
4.5.11. N-((3E,5Z)-3,5-Dodecadienoyl)homoserine Lactone (13)
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zan, J.; Liu, Y.; Fuqua, C.; Hill, R. Acyl-homoserine Lactone Quorum Sensing in the Roseobacter Clade. Int. J. Mol. Sci. 2014, 15, 654–669. [Google Scholar] [CrossRef] [PubMed]
- Buchan, A.; Mitchell, A.; Nathan Cude, N.W.; Campagna, S. Acyl-homoserine Lactone-Based Quorum Sensing in Members of the Marine Bacterial Roseobacter Clade: Complex Cell-to-Cell Communication Controls Multiple Physiologies. In Stress and Environmental Regulation of Gene Expression and Adaptation in Bacteria; de Bruijn, F.J., Ed.; Wiley Blackwell: Hoboken, NJ, USA; pp. 225–233.
- Schulz, S.; Hötling, S. The use of the lactone motif in chemical communication. Nat. Prod. Rep. 2015, 32, 1042–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patzelt, D.; Wang, H.; Buchholz, I.; Rohde, M.; Gröbe, L.; Pradella, S.; Neumann, A.; Schulz, S.; Heyber, S.; Münch, K.; et al. You are what you talk: Quorum sensing induces individual morphologies and cell division modes in Dinoroseobacter shibae. ISME J. 2013, 7, 2274–2286. [Google Scholar] [CrossRef] [PubMed]
- Berger, M.; Neumann, A.; Schulz, S.; Simon, M.; Brinkhoff, T. Tropodithietic acid production in Phaeobacter gallaeciensis is regulated by N-acyl homoserine lactone-mediated quorum sensing. J. Bacteriol. 2011, 193, 6576–6585. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ziesche, L.; Frank, O.; Michael, V.; Martin, M.; Petersen, J.; Schulz, S.; Wagner-Döbler, I.; Tomasch, J. The CtrA phosphorelay integrates differentiation and communication in the marine alphaproteobacterium Dinoroseobacter shibae. BMC Genomics 2014, 15, 130. [Google Scholar] [CrossRef] [PubMed]
- Dickschat, J.S. Quorum sensing and bacterial biofilms. Nat. Prod. Rep. 2010, 27, 343–369. [Google Scholar] [CrossRef] [PubMed]
- Giebel, H.-A.; Kalhoefer, D.; Lemke, A.; Thole, S.; Gahl-Janssen, R.; Simon, M.; Brinkhoff, T. Distribution of Roseobacter RCA and SAR11 lineages in the North Sea and characteristics of an abundant RCA isolate. ISME J. 2011, 5, 8–19. [Google Scholar] [CrossRef]
- Freese, H.M.; Methner, A.; Overmann, J. Adaptation of Surface-Associated Bacteria to the Open Ocean. Front. Microbiol. 2017, 8, 1659. [Google Scholar] [CrossRef]
- Buchan, A.; Gonzalez, J.M.; Moran, M.A. Overview of the Marine Roseobacter Lineage. Appl. Environ. Microbiol. 2005, 71, 5665–5677. [Google Scholar] [CrossRef] [Green Version]
- Brinkhoff, T.; Giebel, H.-A.; Simon, M. Diversity, ecology, and genomics of the Roseobacter clade: A short overview. Arch. Microbiol. 2008, 189, 531–539. [Google Scholar] [CrossRef]
- Ziesche, L.; Bruns, H.; Dogs, M.; Wolter, L.; Mann, F.; Wagner-Döbler, I.; Brinkhoff, T.; Schulz, S. Homoserine Lactones, Methyl Oligohydroxybutyrates, and Other Extracellular Metabolites of Macroalgae-Associated Bacteria of the Roseobacter Clade: Identification and Functions. ChemBioChem 2015, 16, 2094–2107. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, A.L.; Greenberg, E.P.; Oliver, C.M.; Oda, Y.; Huang, J.J.; Bittan-Banin, G.; Peres, C.M.; Schmidt, S.; Juhaszova, K.; Sufrin, J.R.; et al. A new class of homoserine lactone quorum-sensing signals. Nature 2008, 454, 595–599. [Google Scholar] [CrossRef] [PubMed]
- Cude, W.N.; Buchan, A. Acyl-homoserine lactone-based quorum sensing in the Roseobacter clade: Complex cell-to-cell communication controls multiple physiologies. Front. Microbiol. 2013, 4, 336. [Google Scholar] [CrossRef] [PubMed]
- Wagner-Döbler, I.; Thiel, V.; Eberl, L.; Allgaier, M.; Bodor, A.; Meyer, S.; Ebner, S.; Hennig, A.; Pukall, R.; Schulz, S. Discovery of complex mixtures of novel long-chain quorum sensing signals in free-living and host-associated marine Alphaproteobacteria. ChemBioChem 2005, 6, 2195–2206. [Google Scholar] [CrossRef] [PubMed]
- Thiel, V.; Kunze, B.; Verma, P.; Wagner-Döbler, I.; Schulz, S. New Structural Variants of Homoserine Lactones in Bacteria. ChemBioChem 2009, 10, 1861–1868. [Google Scholar] [CrossRef]
- Bruns, H.; Thiel, V.; Voget, S.; Patzelt, D.; Daniel, R.; Wagner-Döbler, I.; Schulz, S. N-acylated alanine methyl esters (NAMEs) from Roseovarius tolerans, structural analogs of quorum-sensing autoinducers, N-acylhomoserine lactones. Chem. Biodivers. 2013, 10, 1559–1573. [Google Scholar] [CrossRef]
- Bruns, H.; Herrmann, J.; Müller, R.; Wang, H.; Wagner Döbler, I.; Schulz, S. Oxygenated N-Acyl Alanine Methyl Esters (NAMEs) from the Marine Bacterium Roseovarius tolerans EL-164. J. Nat. Prod. 2018, 81, 131–139. [Google Scholar] [CrossRef]
- Kanukollu, S.; Wemheuer, B.; Herber, J.; Billerbeck, S.; Lucas, J.; Daniel, R.; Simon, M.; Cypionka, H.; Engelen, B. Distinct compositions of free-living, particle-associated and benthic communities of the Roseobacter group in the North Sea. FEMS Microbiol. Ecol. 2016, 92, fiv145. [Google Scholar] [CrossRef]
- Neumann, A.; Patzelt, D.; Wagner-Döbler, I.; Schulz, S. Identification of new N-acylhomoserine lactone signalling compounds of Dinoroseobacter shibae DFL-12T by overexpression of luxI genes. ChemBioChem 2013, 14, 2355–2361. [Google Scholar] [CrossRef]
- Doberva, M.; Stien, D.; Sorres, J.; Hue, N.; Sanchez-Ferandin, S.; Eparvier, V.; Ferandin, Y.; Lebaron, P.; Lami, R. Large Diversity and Original Structures of Acyl-homoserine Lactones in Strain MOLA 401, a Marine Rhodobacteraceae Bacterium. Front. Microbiol. 2017, 8, 1152. [Google Scholar] [CrossRef]
- Camara, M.; Daykin, M.; Chhabra, S.R. Detection, purification, and synthesis of N-acylhomoserine lactone quorum sensing signal molecules. Meth. Microbiol. 1998, 27, 319–330. [Google Scholar]
- Majewski, M.; Green, J.R.; Snieckus, V. Stereoselective deprotonation of α,β-unsaturated amides. Tetrahedron Lett. 1986, 27, 531–534. [Google Scholar] [CrossRef]
- Theodorou, V.; Gogou, M.; Philippidou, M.; Ragoussis, V.; Paraskevopoulos, G.; Skobridis, K. Synthesis of β,γ-unsaturated primary amides from α,β-unsaturated acids and investigation of the mechanism. Tetrahedron 2011, 67, 5630–5634. [Google Scholar] [CrossRef]
- Tsukada, N.; Sato, T.; Inoue, Y. Palladium-catalyzed allylic alkenylation of allylic alcohols with n-butyl acrylate. Chem. Commun. 2003, 19, 2404–2405. [Google Scholar] [CrossRef]
- Jung, M.F.; Lyster, M.A. Quantitative dealkylation of alkyl esters via treatment with trimethylsilyl iodide. A new method for ester hydrolysis. J. Am. Chem. Soc. 1977, 99, 968–969. [Google Scholar] [CrossRef]
- Riedel, K.; Hentzer, M.; Geisenberger, O.; Huber, B.; Steidle, A.; Wu, H.; Hoiby, N.; Givskov, M.; Molin, S.; Eberl, L. N-Acylhomoserine-lactone-mediated communication between Pseudomonas aeruginosa and Burkholderia cepacia in mixed biofilms. Microbiology 2001, 147, 3249–3262. [Google Scholar] [CrossRef] [PubMed]
- Steidle, A.; Allesen-Holm, M.; Riedel, K.; Berg, G.; Givskov, M.; Molin, S.; Eberl, L. Identification and characterization of an N-acylhomoserine lactone-dependent quorum-sensing system in Pseudomonas putida strain IsoF. Appl. Environ. Microbiol. 2002, 68, 6371–6382. [Google Scholar] [CrossRef]
- Crowe, D.; Nicholson, A.; Fleming, A.; Carey, E.; Sánchez-Sanz, G.; Kelleher, F. Conformational studies of Gram-negative bacterial quorum sensing 3-oxo N-acyl homoserine lactone molecules. Bioorg. Med. Chem. 2017, 25, 4285–4296. [Google Scholar] [CrossRef]
- Ziesche, L.; Rinkel, J.; Dickschat, J.S.; Schulz, S. Acyl-group specificity of AHL synthases involved in quorum-sensing in Roseobacter group bacteria. Beilstein J. Org. Chem. 2018, 14, 1309–1316. [Google Scholar] [CrossRef]
- Schulz, S. Composition of the silk lipids of the spider Nephila clavipes. Lipids 2001, 36, 637–647. [Google Scholar] [CrossRef]
- Sass, H.; Köpke, B.; Rütters, H.; Feuerlein, T.; Dröge, S.; Cypionka, H.; Engelen, B. Tateyamaria pelophila sp. nov., a facultatively anaerobic alphaproteobacterium isolated from tidal-flat sediment, and emended descriptions of the genus Tateyamaria and of Tateyamaria omphalii. Int. J. Syst. Evol. Microbiol. 2010, 60, 1770–1777. [Google Scholar] [CrossRef] [PubMed]
- Kanukollu, S.; Voget, S.; Pohlner, M.; Vandieken, V.; Petersen, J.; Kyrpides, N.C.; Woyke, T.; Shapiro, N.; Göker, M.; Klenk, H.-P.; et al. Genome sequence of Shimia str. SK013, a representative of the Roseobacter group isolated from marine sediment. Stand. Genom. Sci. 2016, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Pohlner, M.; Degenhardt, J.; von Hoyningen-Huene Avril, J.E.; Wemheuer, B.; Erlmann, N.; Schnetger, B.; Badewien, T.H.; Engelen, B. The Biogeographical Distribution of Benthic Roseobacter Group Members along a Pacific Transect Is Structured by Nutrient Availability within the Sediments and Primary Production in Different Oceanic Provinces. Front. Microbiol. 2017, 8, 2550. [Google Scholar] [CrossRef] [PubMed]
- Angle, S.R.; Henry, R.M. Studies toward the Synthesis of (+)-Palustrine. J. Org. Chem. 1998, 63, 7490–7497. [Google Scholar] [CrossRef] [PubMed]
- Clerc, J.; Li, N.; Krahn, D.; Groll, M.; Bachmann, A.S.; Florea, B.I.; Overkleeft, H.S.; Kaiser, M. The natural product hybrid of Syringolin A and Glidobactin A synergizes proteasome inhibition potency with subsite selectivity. Chem. Commun. 2011, 47, 385–387. [Google Scholar] [CrossRef]
- Kleinbeck, F.; Toste, F.D. Gold (I)-catalyzed enantioselective ring expansion of allenylcyclopropanols. J. Am. Chem. Soc. 2009, 131, 9178–9179. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genus Affiliation | C12:0 | C12:1 | C12:2 (11) | C12:2 (13) | 3-OH-C10 | 3-OH-C14 | C14:0 | C14:1 | C14:2 | C15:0 | C15:1 | C16:0 | C16:1 | C16:2 | C17:1 | C18:0 | C18:1 | C18:2 | C18:2 | C18:2 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fucus Spiralis | |||||||||||||||||||||
| D12-1.68 | Roseovarius sp. | 94.4 | 1.3 | 4.3 | |||||||||||||||||
| D3 | Loktanella sp. | 15.7 | 2.4 | 21.2 | 60.7 | ||||||||||||||||
| Lw-22 | Octadecabacter sp. | 6.6 | 3.7 | 0.7 | 4.6 | 72.7 | 4.3 | 1.2 | 1.9 | 0.9 | 0.6 | 1.1 | 1.8 | ||||||||
| D4 (50) | Octadecabacter sp. | 8.3 | 25.8 | 4.4 | 1.1 | 0.7 | 59.8 | ||||||||||||||
| Lw-26b | Loktanella sp. | 100 | |||||||||||||||||||
| D15 (40) | Loktanella sp. | 3.9 | 5.9 | 1.2 | 0.9 | 18.4 | 2.3 | 58.6 | 0.5 | 0.8 | 0.6 | 6.1 | 0.5 | 0.3 | |||||||
| Lw-55a | Loktanella sp. | 100 | |||||||||||||||||||
| MDLw-58 | Dinoroseobacter sp. | 5.4 | 3.9 | 2.5 | 88.2 | ||||||||||||||||
| Sediment | |||||||||||||||||||||
| SK013 | Shimia sp. | 100 | |||||||||||||||||||
| SK032 | Huaishuia sp. | 81.7 | 18.3 | ||||||||||||||||||
| SK040 | Phaeobacter sp. | 5.8 | 3.7 | 1.4 | 7.3 | 4.4 | 4.8 | 64.9 | 3.5 | 4.1 | |||||||||||
| Water Column | |||||||||||||||||||||
| SK038 | Sulfitobacter sp. | 100 | |||||||||||||||||||
| AHLa | MT102 | F117 |
|---|---|---|
| 2E,4E-C12:2 (5) | 1.00 ± 0.01 | 81.22 ± 1.26 |
| 2E,5Z-C12:2 (11) | 1.01 ± 0.01 | 78.03 ± 2.05 |
| 3E,5Z-C12:2 (13) | 1.03 ± 0 | 73.64 ± 2.34 |
| 2E,11Z-C18:2 | 1.02 ± 0.01 | 1.32 ± 0.04 |
| 3-oxo-C8:0 | 13.41± 0.49 | 30.46 ± 2.79 |
| C6:0 | 17.09 ± 1.04 | |
| C8:0 | 10.24 ± 0.55 | |
| 3-oxo-C12:0 | 77.75 ± 2.51 | |
| Z9-C16:1 | 72.37 ± 2.47 |
| Strain | GenBank Acc No (16S) | Genus Affiliation | Strain Origin/Host | Location |
|---|---|---|---|---|
| Lw-III1a | KM268064 | Puniceibacterium sp. | Fucus spiralis | Neuharlingersiel |
| B15 G2 | KM268068 | Sulfitobacter sp. | Fucus spiralis | Neuharlingersiel |
| Lw-13e | KM268063 | Pseudooceanicola sp. | Fucus spiralis | Neuharlingersiel |
| D12-1.68 | KM268065 | Roseovarius sp. | Fucus spiralis | Neuharlingersiel |
| B14 | KM268066 | Roseobacter sp. | Fucus spiralis | Neuharlingersiel |
| B14 27 | KM268072 | Sulfitobacter sp. | Fucus spiralis | Neuharlingersiel |
| A12 | KM268070 | Sulfitobacter sp. | Fucus spiralis | Neuharlingersiel |
| D3 | KC731427 | Loktanella sp. | Fucus spiralis | Neuharlingersiel |
| D4 55 | KM268071 | Sulfitobacter sp. | Fucus spiralis | Neuharlingersiel |
| Lw-22 | KM268073 | Octadecabacter sp. | Fucus spiralis | Neuharlingersiel |
| D4 50 | KM268074 | Octadecabacter sp. | Fucus spiralis | Neuharlingersiel |
| Lw-26b | KM268054 | Loktanella sp. | Fucus spiralis | Neuharlingersiel |
| D15 40 | KM268056 | Loktanella sp. | Fucus spiralis | Neuharlingersiel |
| Lw-55a | KM268057 | Loktanella sp. | Fucus spiralis | Neuharlingersiel |
| MDLw-58 | KM268059 | Dinoroseobacter sp. | Fucus spiralis | Neuharlingersiel |
| Lw-41a | KM268061 | Citreicella sp. | Fucus spiralis | Neuharlingersiel |
| N05I | AJ968647 | Phaeobacter sp. | German North Sea Coast | Tidal-flat sediment |
| G5II | AJ968650 | Roseovarius pelophilus | German North Sea Coast | Tidal-flat sediment |
| SAM4 | AJ968651 | Tateyamaria pelophilaa | German North Sea Coast | Tidal-flat sediment |
| SK012 | HG423260 | Sulfitobacter sp. | Danish North Sea | sediment |
| SK013 | LAJH00000000 | Shimia sp. b | Norwegian North Sea | sediment |
| SK021 | HG423263 | Pseudoruegeria sp. b | Norwegian North Sea | sediment |
| SK032 | HG423269 | Huaishuia sp. | German North Sea | sediment |
| SK033 | HG423270 | Loktanella sp. | Norwegian North Sea | sediment |
| SK040 | HG423272 | Phaeobacter sp. | Danish North Sea | sediment |
| SK038 | HG423271 | Sulfitobacter sp. | German North Sea | seawater |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ziesche, L.; Wolter, L.; Wang, H.; Brinkhoff, T.; Pohlner, M.; Engelen, B.; Wagner-Döbler, I.; Schulz, S. An Unprecedented Medium-Chain Diunsaturated N-acylhomoserine Lactone from Marine Roseobacter Group Bacteria. Mar. Drugs 2019, 17, 20. https://doi.org/10.3390/md17010020
Ziesche L, Wolter L, Wang H, Brinkhoff T, Pohlner M, Engelen B, Wagner-Döbler I, Schulz S. An Unprecedented Medium-Chain Diunsaturated N-acylhomoserine Lactone from Marine Roseobacter Group Bacteria. Marine Drugs. 2019; 17(1):20. https://doi.org/10.3390/md17010020
Chicago/Turabian StyleZiesche, Lisa, Laura Wolter, Hui Wang, Thorsten Brinkhoff, Marion Pohlner, Bert Engelen, Irene Wagner-Döbler, and Stefan Schulz. 2019. "An Unprecedented Medium-Chain Diunsaturated N-acylhomoserine Lactone from Marine Roseobacter Group Bacteria" Marine Drugs 17, no. 1: 20. https://doi.org/10.3390/md17010020