Structural Elucidation of a Novel Lipooligosaccharide from the Cold-Adapted Bacterium OMVs Producer Shewanella sp. HM13



Abstract
:1. Introduction
2. Results and Discussion
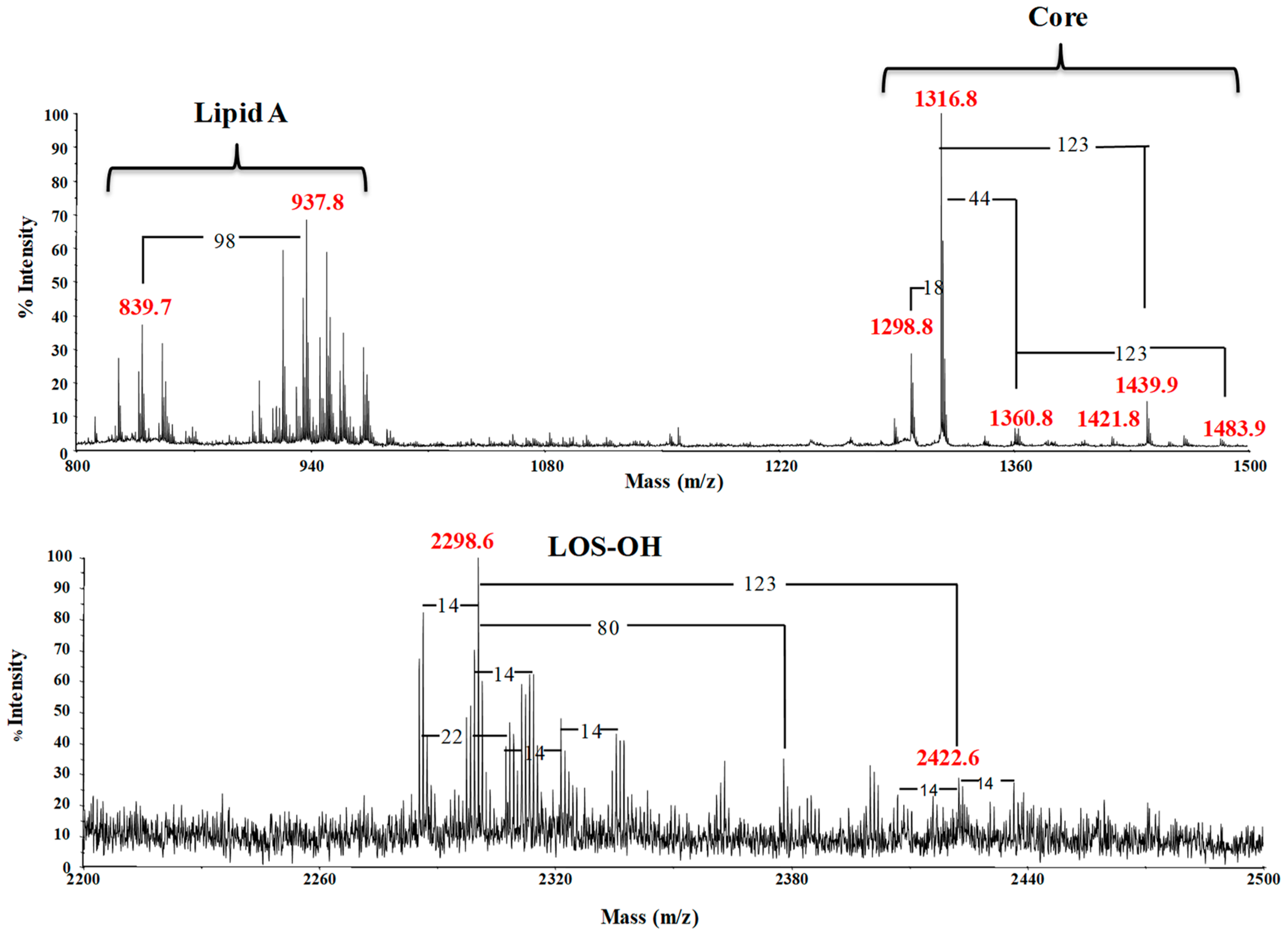
2.1. LOS Extraction and Purification
2.2. LOS Deacylation
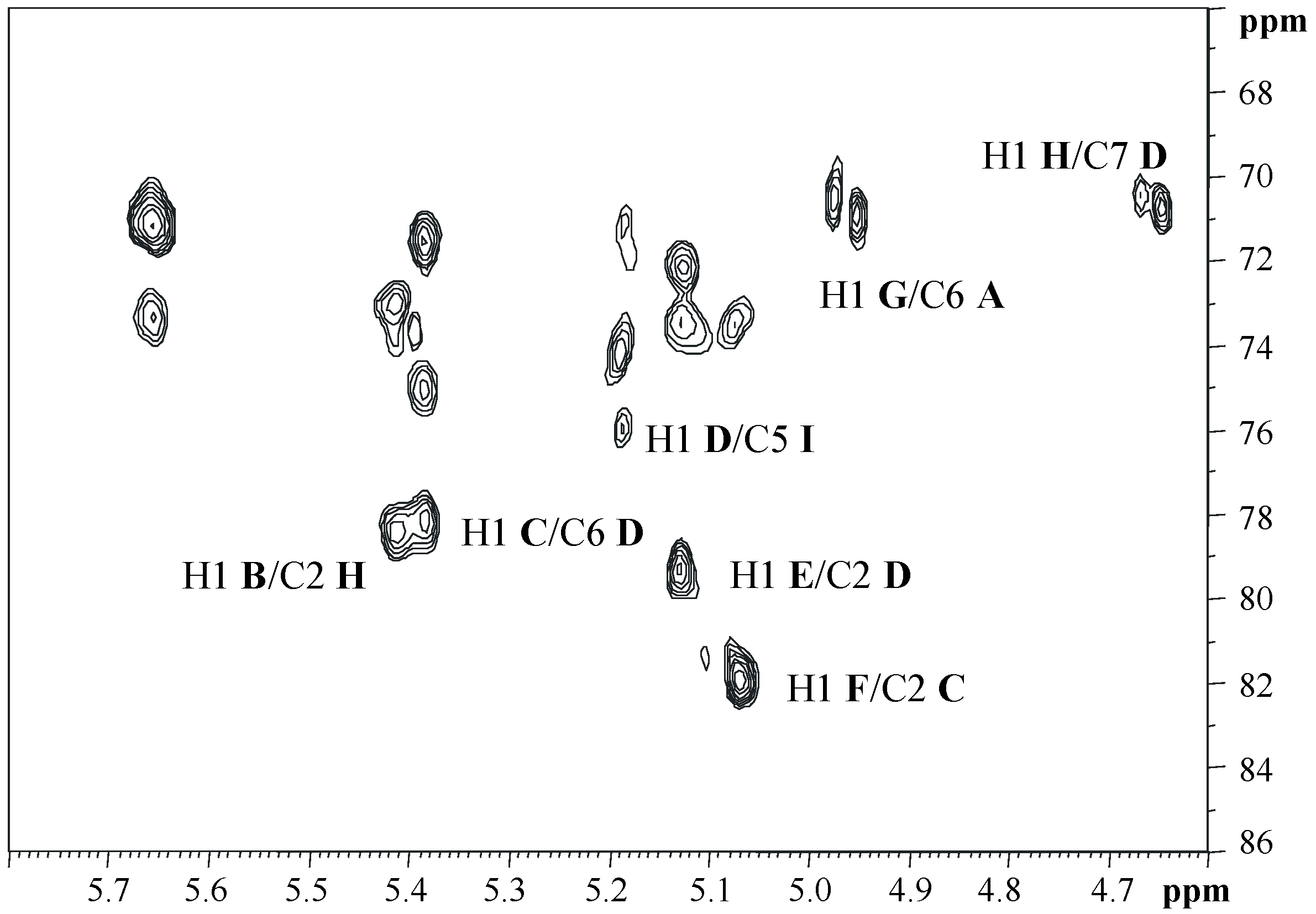
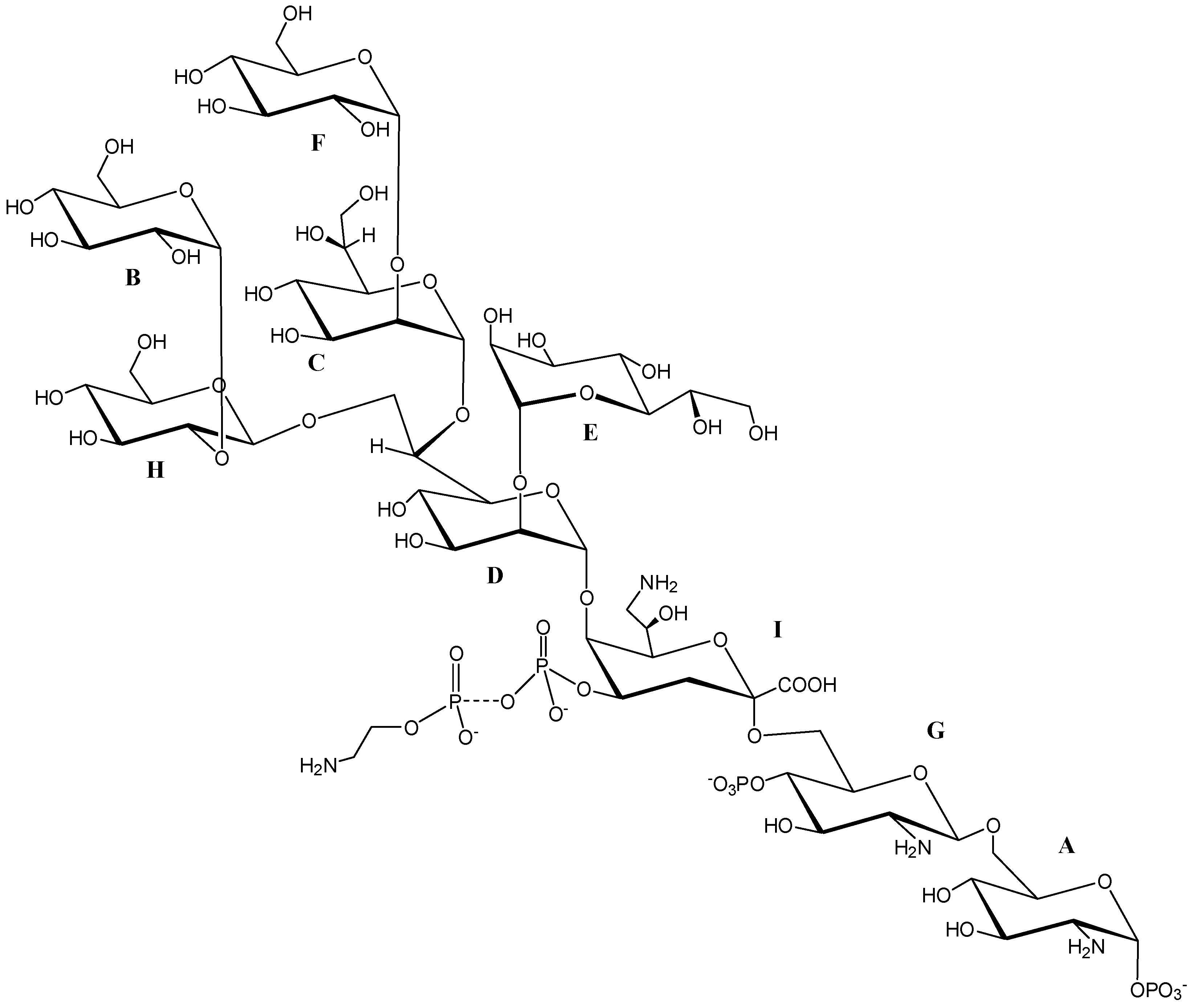
2.3. NMR Spectroscopic Analysis of OS
3. Materials and Methods
3.1. Bacteria Growth and LPS Isolation
3.2. DOC-PAGE Analysis
3.3. Sugar and Fatty Acids Analysis
3.4. Deacylation of the LOS
3.5. Methylation Analysis
3.6. Mass Spectrometry Analysis
3.7. NMR Spectroscopy
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Feller, G.; Gerday, C. Psychrophilic enzymes: Hot topics in cold adaptation. Nat. Rev. Microbiol. 2003, 1, 200–208. [Google Scholar] [CrossRef]
- Casillo, A.; Papa, R.; Ricciardelli, A.; Sannino, F.; Ziaco, M.; Tilotta, M.; Selan, L.; Marino, G.; Corsaro, M.M.; Tutino, M.L.; et al. Anti-biofilm activity of a long-chain fatty aldehyde from Antarctic Pseudoalteromonas haloplanktis TAC125 against Staphylococcus epidermidis biofilm. Front. Cell. Infect. Microbiol. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Perfumo, A.; Banat, I.M.; Marchant, R. Going Green and Cold: Biosurfactants from low-temperature environments to biotechnology applications. Trends Biotechnol. 2018, 36, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Feller, G. Protein stability and enzyme activity at extreme biological temperatures. J. Phys. Condens. Matter 2010, 22, 323101–323117. [Google Scholar] [CrossRef] [PubMed]
- Bar Dolev, M.; Braslavsky, I.; Davies, P.L. Ice-Binding proteins and their function. Annu. Rev. Biochem. 2016, 85, 515–542. [Google Scholar] [CrossRef] [PubMed]
- Carillo, S.; Casillo, A.; Pieretti, G.; Parrilli, E.; Sannino, F.; Bayer-Giraldi, M.; Cosconati, S.; Novellino, E.; Ewert, M.; Deming, J.W.; et al. A unique capsular polysaccharide structure from the psychrophilic marine bacterium Colwelliapsychrerythraea 34H that mimicks antifreeze (glyco) proteins. J. Am. Chem. Soc. 2015, 137, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Casillo, A.; Parrilli, E.; Sannino, F.; Mitchell, D.E.; Gibson, M.I.; Marino, G.; Lanzetta, R.; Parrilli, M.; Cosconati, S.; Novellino, E.; et al. Structure-activity relationship of the exopolysaccharide from a psychrophilic bacterium: A strategy for cryoprotection. Carbohydr. Polym. 2017, 156, 364–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frias, A.; Manresa, A.; de Oliveira, E.; López-Iglesias, C.; Mercade, E. Membrane Vesicles: A common feature in the extracellular matter of cold-adapted Antarctic Bacteria. Microb. Ecol. 2010, 59, 476–486. [Google Scholar] [CrossRef]
- Nevot, M.; Deroncelé, V.; Messner, P.; Guinea, J.; Mercadé, E. Characterization of outer membrane vesicles released by the psychrotolerant bacterium Pseudoalteromonas antarctica NF3. Environ. Microbiol. 2006, 8, 1523–1533. [Google Scholar] [CrossRef]
- Pérez-Cruz, C.; Mercadé, E. Outer Membrane Vesicles From Cold-Adapted Antarctic Bacteria; Recent Advances in Pharmaceutical Sciences IV; Muñoz-Torrero, D., Vázquez-Carrera, M., Estelrich, J., Eds.; Research Signpost: Kerala, India, 2014; pp. 55–72. ISBN 978-81-308-0554-2. [Google Scholar]
- Schwechheimer, C.; Kuehn, M.J. Outer-membrane vesicles from Gram-negative bacteria: Biogenesis and functions. Nat. Rev. Microbiol. 2015, 13, 605–619. [Google Scholar] [CrossRef]
- Deatheragea, B.L.; Cooksona, B.T. Membrane vesicle release in Bacteria, Eukaryotes, and Archaea: A conserved yet underappreciated aspect of microbial life. Infect. Immun. 2012, 80, 1948–1957. [Google Scholar] [CrossRef] [PubMed]
- Caroff, M.; Karibian, D. Structure of bacterial lipopolysaccharides. Carbohydr. Res. 2003, 338, 2431–2447. [Google Scholar] [CrossRef] [PubMed]
- Carillo, S.; Pieretti, G.; Bedini, E.; Parrilli, M.; Lanzetta, R.; Corsaro, M.M. Structural investigation of the antagonist LPS from the cyanobacterium Oscillatoria planktothrix FP1. Carbohydr. Res. 2014, 388, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Westphal, O.; Luderitz, O.; Galanos, C.; Mayer, H.; Rietschel, E.T. Advances in Immunopharmacology; Chedid, L., Hadden, J.W., Spreafico, F., Eds.; Pergamon Press: Oxford, UK, 1986; pp. 13–34. [Google Scholar]
- Wilkinson, S.C. Surface Carbohydrates of the Procaryotic Cell; Sutherland, I.W., Ed.; Academic Press Inc.: New York, NY, USA, 1977; pp. 97–105. [Google Scholar]
- Alexander, C.; Rietschel, E.T. Bacterial lipopolysaccharides and innate immunity. J. Endoxin Res. 2001, 7, 167–202. [Google Scholar] [CrossRef]
- Chen, C.; Kawamoto, J.; Kawai, S.; Tame, A.; Kato, C.; Imai, T.; Kurihara, T. Isolation of a novel bacterial strain capable of producing abundant extracellular membrane vesicles carrying a single major cargo protein and analysis of its transport mechanism. Appl. Environ. Microbiol. under review.
- Galanos, C.; Lüderitz, O.; Westphal, O. New Method for the extraction of R Lipopolysaccharides. Eur. J. Biochem. 1969, 9, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Westphal, O.; Jann, K. Bacterial lipopolysaccharides extraction with phenol-water and further applications of the procedure. Methods Carbohydr. Chem. 1965, 5, 83–91. [Google Scholar]
- Vinogradov, E.; Kubler-Kielb, J.; Korenevsky, A. The structure of the carbohydrate backbone of the LPS from Shewanella spp. MR-4. Carbohydr. Res. 2008, 343, 2701–2705. [Google Scholar] [CrossRef]
- Vinogradov, E.; Korenevsky, A.; Beveridge, T.J. The structure of the carbohydrate backbone of the LPS from Shewanella putrefaciens CN32. Carbohydr. Res. 2002, 337, 1285–1289. [Google Scholar] [CrossRef]
- Vinogradov, E.; Korenevsky, A.; Beveridge, T.J. The structure of the rough-type lipopolysaccharide from Shewanella oneidensis MR-1, containing 8-amino-8-deoxy-Kdo and an open-chain form of 2-acetamido-2-deoxy-d-galactose. Carbohydr. Res. 2003, 338, 1991–1997. [Google Scholar] [CrossRef]
- Silipo, A.; Leone, S.; Molinaro, A.; Sturiale, L.; Garozzo, D.; Nazarenko, E.L.; Gorshkova, R.P.; Ivanova, E.P.; Lanzetta, R.; Parrilli, M. Complete structural elucidation of a novel lipooligosaccharide from the outer membrane of the marine bacterium Shewanella pacifica. Eur. J. Org. Chem. 2005, 11, 2281–2291. [Google Scholar] [CrossRef]
- Gibson, B.W.; Engstrom, J.J.; Hines, C.M.W.; Falick, A.M. Characterization of bacterial lipooligosaccharides by delayed extraction matrix-assisted laser desorption ionization time-of-flight mass spectrometry. J. Am. Soc. Mass. Spectrom. 1997, 8, 645–658. [Google Scholar] [CrossRef] [Green Version]
- Pieretti, G.; Carillo, S.; Lindner, B.; Kim, K.K.; Lee, K.C.; Lee, J.S.; Lanzetta, R.; Parrilli, M.; Corsaro, M.M. Characterization of the core oligosaccharide and the O-antigen biological repeating unit from Halomonasstevensii LPS: The first case of O-antigen linked to the inner core. Chem. Eur. J. 2012, 18, 3729–3735. [Google Scholar] [CrossRef] [PubMed]
- Vinogradov, E.; Korenevsky, A.; Beveridge, T.J. The structure of the core region of the lipopolysaccharide from Shewanella algae BrY, containing 8-amino-3,8-dideoxy-d-mannooct-2-ulosonic acid. Carbohydr. Res. 2004, 339, 737–740. [Google Scholar] [CrossRef] [PubMed]
- Lipkind, G.M.; Shashkov, A.S.; Knirel, Y.A.; Vinogradov, E.V.; Kochetkov, N.K. A computer-assisted structural analysis of regular polysaccharides on the basis of 13C-NMR data. Carbohydr. Res. 1988, 175, 59–75. [Google Scholar] [CrossRef]
- Birnbaum, G.I.; Roy, R.; Brisson, J.R.; Jennings, H.J.J. Conformations of ammonium 3-deoxy-d-manno-2-octulosonate (KDO) and methyl α-and β-ketopyranosides of KDO: X-ray structure and 1H NMR analyses. Carbohydr. Chem. 1987, 6, 17–39. [Google Scholar] [CrossRef]
- Bock, K.; Vinogradov, E.V.; Holst, O.; Brade, H. Isolation and structural analysis of oligosaccharide phosphates containing the complete carbohydrate chain of the lipopolysaccharide from Vibrio cholerae Strain H11 (Non-O1). Eur. J. Biochem. 1994, 225, 1029–1039. [Google Scholar] [CrossRef]
- Laemmli, U.K. Most commonly used discontinuous buffer system for SDS electrophoresis. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Casillo, A.; Ståhle, J.; Parrilli, E.; Sannino, F.; Mitchell, D.E.; Pieretti, G.; Gibson, M.I.; Marino, G.; Lanzetta, R.; Parrilli, M.; et al. Structural characterization of an all-aminosugar-containing capsular polysaccharide from Colwellia psychrerythraea 34H. Antonie Leeuwenhoek 2017, 110, 1377–1387. [Google Scholar] [CrossRef]
- Tsai, C.M.; Frasch, C.E. A sensitive silver stain for detecting lipopolysaccharides in polyacrylamide gels. Anal. Biochem. 1982, 119, 115–119. [Google Scholar] [CrossRef]
- Casillo, A.; Ziaco, M.; Lindner, B.; Parrilli, E.; Schwudke, D.; Holgado, A.; Verstrepen, L.; Sannino, F.; Beyaert, R.; Lanzetta, R.; et al. Unusual lipid A from a cold-adapted bacterium: Detailed structural characterization. ChemBioChem 2017, 18, 1845–1854. [Google Scholar] [CrossRef] [PubMed]
- Leontein, K.; Lindberg, B.; LoÈnngren, J. Assignment of absolute configuration of sugars by g.l.c. of their acetylated glycosides from chiral alcohols. Carbohydr. Res. 1978, 62, 359–362. [Google Scholar] [CrossRef]
- Pieretti, G.; Corsaro, M.M.; Lanzetta, R.; Parrilli, M.; Canals, R.; Merino, S.; Tomás, J.M. Structural studies of the O’glycosides from chiral alco Plesiomonas shigelloides Strain 302–73 (Serotype O1). Eur. J. Org. Chem. 2008, 18, 3149–3155. [Google Scholar] [CrossRef]
- Holst, O. De-acylation of lipopolysaccharides and isolation of oligosaccharide phosphates. In Bacterial Toxins: Methods and Protocols; Methods in Molecular Biology; Holst, O., Ed.; Humana Press: Totowa, NJ, USA, 2000; Volume 145, pp. 345–353. [Google Scholar]
- Ciucanu, I.; Kerek, F. A simple and rapid method for the permethylation of carbohydrates. Carbohydr. Res. 1984, 131, 209–217. [Google Scholar] [CrossRef]
- De Castro, C.; Lanzetta, R.; Leone, S.; Parrilli, M.; Molinaro, A. The structural elucidation of the Salmonella enterica subsp. enterica, reveals that it contains both O-factors 4 and 5 on the LPS antigen. Carbohydr. Res. 2013, 370, 9–12. [Google Scholar] [CrossRef]
- Moule, A.L.; Galbraith, L.; Cox, A.D.; Wilkinson, S.G. Characterisation of a tetrasaccharide released on mild acid hydrolysis of LPS from two rough strains of Shewanella species representing different DNA homology groups. Carbohydr. Res. 2004, 339, 1185–1188. [Google Scholar] [CrossRef] [PubMed]
- Corsaro, M.M.; Lanzetta, R.; Parrilli, E.; Parrilli, M.; Tutino, M.L. Structural investigation on the lipooligosaccharide fraction of psychrophilic Pseudoalteromonas haloplanktis TAC 125 bacterium. Eur. J. Biochem. 2001, 268, 5092–5097. [Google Scholar] [CrossRef]
- Corsaro, M.M.; Pieretti, G.; Lindner, B.; Lanzetta, R.; Parrilli, E.; Tutino, M.L.; Parrilli, M. Highly phosphorylated core oligosaccharide structures from cold-adapted Psychromon asarctica. Chem. Eur. J. 2008, 14, 9368–9937. [Google Scholar] [CrossRef]
- Carillo, S.; Pieretti, G.; Parrilli, E.; Tutino, M.L.; Gemma, S.; Molteni, M.; Lanzetta, R.; Parrilli, M.; Corsaro, M.M. Structural investigation and biological activity of the lipooligosaccharide from the psychrophilic bacterium Pseudoalteromonas haloplanktis TAB 23. Chem. Eur. J. 2011, 17, 7053–7060. [Google Scholar] [CrossRef]
- Carillo, S.; Pieretti, G.; Lindner, B.; Parrilli, E.; Sannino, F.; Tutino, M.L.; Lanzetta, R.; Parrilli, M.; Corsaro, M.M. Structural Characterization of the core oligosaccharide isolated from the Lipopolysaccharide of the psychrophilic bacterium Colwellia psychrerythraea Strain 34H. Eur. J. Org. Chem. 2013, 3771–3779. [Google Scholar] [CrossRef]
- Casillo, A.; Parrilli, E.; Sannino, F.; Lindner, B.; Lanzetta, R.; Parrilli, M.; Tutino, M.L.; Corsaro, M.M. Structural investigation of the oligosaccharide portion isolated from the lipooligosaccharide of the permafrost psychrophile Psychrobacter arcticus 273-4. Mar. Drugs 2015, 13, 4539–4555. [Google Scholar] [CrossRef] [PubMed]
- Corsaro, M.M.; Casillo, A.; Parrilli, E.; Tutino, M.L. Molecular structure of lipopolysaccharides of cold-adapted bacteria. In Psychrophiles: From Biodiversity to Biotechnology; Margesin, R., Ed.; Springer International Publishing AG: Cham, Switzerland, 2017; Chapter 13; pp. 285–303. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Residue | H1 C1 1JC,H | H2 C2 | H3 C3 | H4 C4 | H5 C5 | H6a C6 | H6b | H7a C7 | H7b | H8a C8 | H8b |
|---|---|---|---|---|---|---|---|---|---|---|---|
| A α-GlcNp1P | 5.65 92.0 179 | 3.36 55.8 | 3.91 71.2 | 3.46 71.4 | 4.20 73.5 | 3.91 70.9 | 4.32 | ||||
| B α-t-Glcp | 5.40 99.1 182 | 3.58 72.5 | 3.76 74.0 | 3.49 70.7 | 4.04 73.5 | 3.76 61.9 | |||||
| C α-2-l,d-Hepp | 5.37 98.8 184 | 3.91 82.1 | 4.05 71.7 | 3.83 68.7 | 3.91 75.2 | 4.07 70.3 | 3.78 64.7 | 3.75 | |||
| D α-2,6,7-d,d-Hepp | 5.17 101.9 170 | 4.08 79.4 | 4.15 71.7 | 3.79 68.7 | 4.06 74.4 | 4.34 78.2 | 4.02 70.8 | 4.13 | |||
| E α-t-l,d-Hepp | 5.12 103.5 173 | 4.07 70.5 | 3.83 72.0 | 3.86 67.6 | 3.66 73.8 | 4.06 70.5 | 3.84 63.5 | 3.72 | |||
| F α-t-Glcp | 5.05 102.3 170 | 3.59 73.1 | 3.72 74.2 | 3.49 70.8 | 4.04 73.0 | 3.81 61.8 | |||||
| G β-6-GlcNp4P | 4.95 100.8 164 | 3.11 56.9 | 3.80 73.6 | 3.85 74.5 | 3.75 75.4 | 3.51 63.4 | 3.73 | ||||
| H β-2-Glcp | 4.65 105.0 164 | 3.47 77.1 | 3.57 75.9 | 3.42 71.4 | 3.44 78.5 | 3.73 62.3 | 3.93 | ||||
| I α-5-Kdo8Np4P | n.d. | 101.2 | 2.01, 2.25 35.8 | 4.58 71.3 | 4.34 76.3 | 3.92 74.6 | 4.02 67.0 | 3.20 44.4 | 3.50 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Casillo, A.; Di Guida, R.; Carillo, S.; Chen, C.; Kamasaka, K.; Kawamoto, J.; Kurihara, T.; Corsaro, M.M. Structural Elucidation of a Novel Lipooligosaccharide from the Cold-Adapted Bacterium OMVs Producer Shewanella sp. HM13. Mar. Drugs 2019, 17, 34. https://doi.org/10.3390/md17010034
Casillo A, Di Guida R, Carillo S, Chen C, Kamasaka K, Kawamoto J, Kurihara T, Corsaro MM. Structural Elucidation of a Novel Lipooligosaccharide from the Cold-Adapted Bacterium OMVs Producer Shewanella sp. HM13. Marine Drugs. 2019; 17(1):34. https://doi.org/10.3390/md17010034
Chicago/Turabian StyleCasillo, Angela, Rossella Di Guida, Sara Carillo, Chen Chen, Kouhei Kamasaka, Jun Kawamoto, Tatsuo Kurihara, and Maria Michela Corsaro. 2019. "Structural Elucidation of a Novel Lipooligosaccharide from the Cold-Adapted Bacterium OMVs Producer Shewanella sp. HM13" Marine Drugs 17, no. 1: 34. https://doi.org/10.3390/md17010034
APA StyleCasillo, A., Di Guida, R., Carillo, S., Chen, C., Kamasaka, K., Kawamoto, J., Kurihara, T., & Corsaro, M. M. (2019). Structural Elucidation of a Novel Lipooligosaccharide from the Cold-Adapted Bacterium OMVs Producer Shewanella sp. HM13. Marine Drugs, 17(1), 34. https://doi.org/10.3390/md17010034