Pyrogallol-Phloroglucinol-6,6-Bieckol Alleviates Obesity and Systemic Inflammation in a Mouse Model by Reducing Expression of RAGE and RAGE Ligands


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Oral Administration of Pyrogallol-Phloroglucinol-6,6-Bieckol Ameliorates Adiposity in a Mouse Model of Diet-Induced Obesity
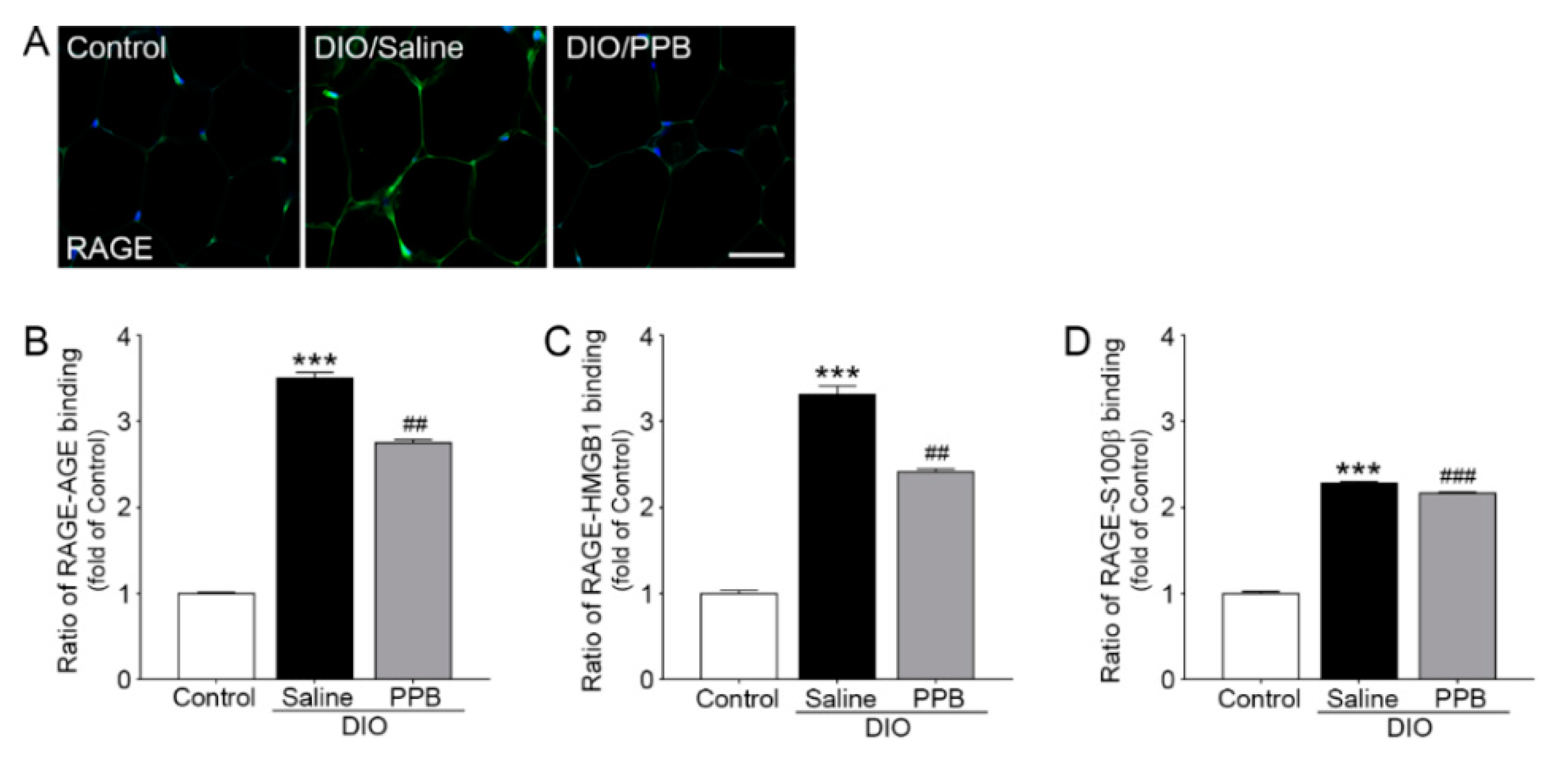
2.2. Reduction in the Expression of AGEs, HMGB1, and S100β Following Supplementation with PPB in DIO Mice
2.3. Reduction of RAGE Expression and RAGE-RAGE Ligand Bonding Following PPB Supplementation in DIO Mice
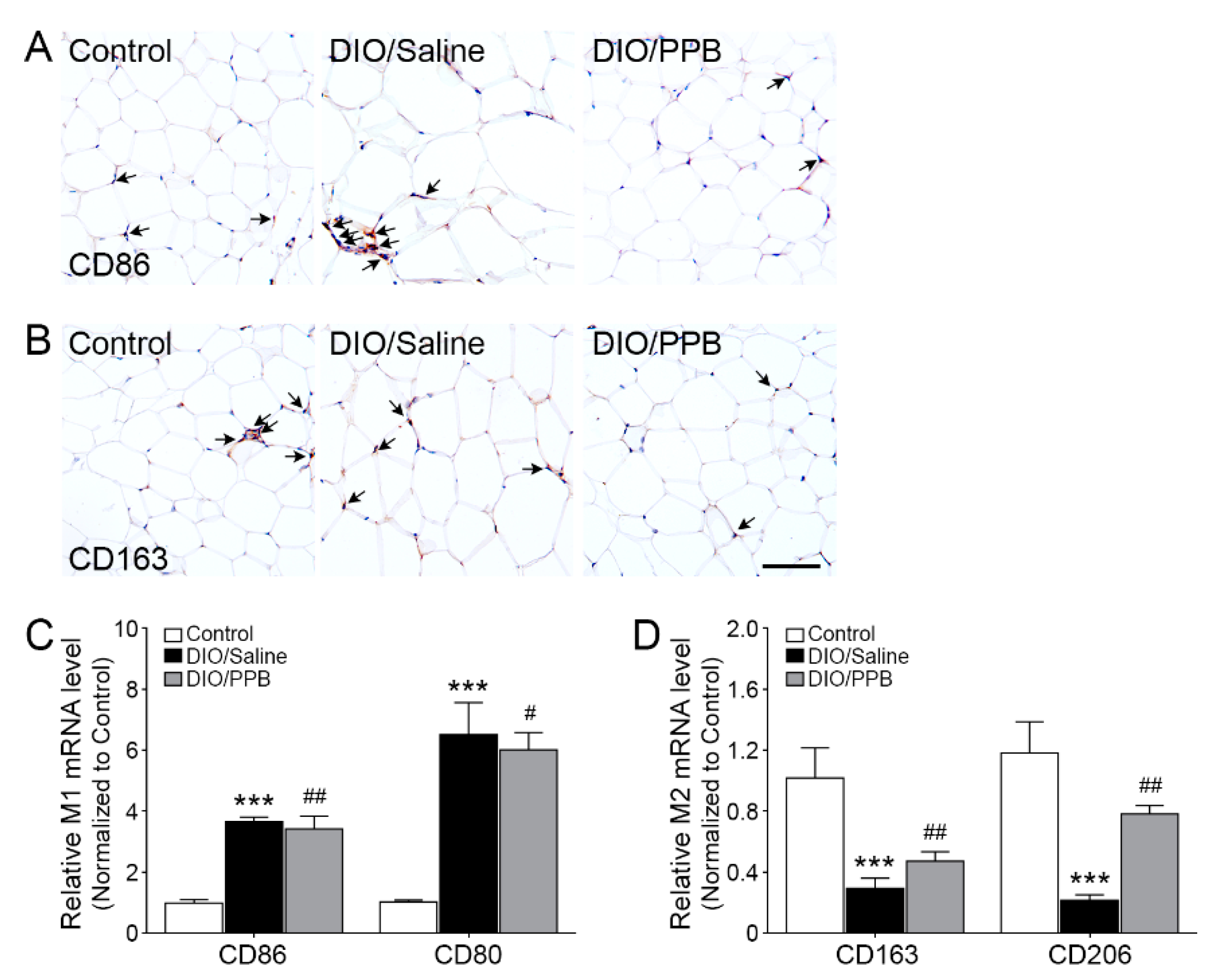
2.4. Modulation of M1/M2 Macrophage Infiltration Following PPB Supplementation of DIO Mice
2.5. Modulation of Inflammatory Cytokine Expression Following PPB Supplementation in DIO Mice
3. Materials and Methods
3.1. Pyrogallol-Phloroglucinol-6,6-Bieckol (PPB) Isolation
3.2. Animals
3.3. Sample Preparation
3.3.1. RNA Extraction and Complimentary DNA (cDNA) Synthesis
3.3.2. Protein Isolation
3.3.3. Paraffin Slide Preparation
3.4. Enzyme-Linked Immunosorbent Assay (ELISA)
3.4.1. Indirect ELISA Assay
3.4.2. Sandwich Enzyme-Linked Immunosorbent Assay
3.5. Immunoblotting
3.6. Quantitative Real Time Polymerase Chain Reaction
3.7. Immunofluorescence
3.8. Histology
3.9. 3,3-Diaminobenzidine (DAB) Staining
3.10. Coomassie Blue Staining
3.11. Fat Size and Intensity Measurement
3.12. Triglyceride Measure Assay
3.13. Total Cholesterol Measure Assay
3.14. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviation
| Arg-1 | Arginase–1 |
| AGEs | Advanced glycation end-products |
| AKT | protein kinase B |
| AMPK | 5′ adenosine monophosphate-activated protein kinase |
| DIO | Diet-induced obesity |
| FFAs | free fatty acids |
| HFD | High-fat diet |
| HMGB1 | High-Mobility Group Box 1 |
| IL | interleukin |
| PI3K | Activating phosphoinositide 3-kinase |
| PPB | pyrogallol-phloroglucinol-6,6-bieckol |
| NF-κB | Nuclear factor kappa-light-chain-enhancer β |
| RAGE | Receptor for advanced glycation end-products |
| TNF-α | tumor necrosis factor alpha |
References
- James, O.H.; Holly, R.W.; John, C.P. Energy Balance and Obesity. Circulation 2012, 126, 126–132. [Google Scholar]
- Han, T.S.; Lean, M.E. A clinical perspective of obesity, metabolic syndrome and cardiovascular disease. JRSM Cardiovasc. Dis. 2016, 25, 2048004016633371. [Google Scholar] [CrossRef] [PubMed]
- Poirier, P.; Giles, T.D.; Bray, G.A.; Hong, Y.; Stern, J.S.; Pi-Sunyer, F.X.; Eckel, R.H.; American Heart Association; Obesity Committee of the Council on Nutrition; Physical Activity; et al. Obesity and cardiovascular disease: Pathophysiology, evaluation, and effect of weight loss: An update of the 1997 American Heart Association Scientific Statement on Obesity and Heart Disease from the Obesity Committee of the Council on Nutrition, Physical Activity, and Metabolism. Circulation 2006, 113, 898–918. [Google Scholar] [PubMed]
- Despres, J.P.; Lemieux, I. Abdominal obesity and metabolic syndrome. Nature 2006, 444, 881–887. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Ren, F.; Lv, J.; Zhang, H.; Guo, H. Lactoferrin attenuates high-fat diet-induced hepatic steatosis and lipid metabolic dysfunctions by suppressing hepatic lipogenesis and down-regulating inflammation in C57BL/6J mice. Food Funct. 2018, 9, 4328–4339. [Google Scholar] [CrossRef]
- Palanissami, G.; Paul, S.F. RAGE and Its Ligands: Molecular Interplay between Glycation, Inflammation, and Hallmarks of Cancer—A Review. Horm. Cancer 2018, 9, 295–325. [Google Scholar] [CrossRef]
- Leuner, B.; Max, M.; Thamm, K.; Kausler, C.; Yakobus, Y.; Bierhaus, A.; Sel, S.; Hofmann, B.; Silber, R.E.; Simm, A.; et al. RAGE influences obesity in mice. Effects of the presence of RAGE on weight gain, AGE accumulation, and insulin levels in mice on a high fat diet. Z. Gerontol. Geriatr. 2012, 45, 102–108. [Google Scholar] [CrossRef]
- Tikellis, C.; Thomas, M.C.; Harcourt, B.E.; Coughlan, M.T.; Pete, J.; Bialkowski, K.; Tan, A.; Bierhaus, A.; Cooper, M.E.; Forbes, J.M. Cardiac inflammation associated with a Western diet is mediated via activation of RAGE by AGEs. Am. J. Physiol. Endocrinol. Metab. 2008, 295, 323–330. [Google Scholar] [CrossRef]
- Dozio, E.; Vianello, E.; Briganti, S.; Lamont, J.; Tacchini, L.; Schmitz, G.; Romanelli, M.M.C. Expression of the Receptor for Advanced Glycation End Products in Epicardial Fat: Link with Tissue Thickness and Local Insulin Resistance in Coronary Artery Disease. J. Diabetes Res. 2016, 2016, 8. [Google Scholar] [CrossRef]
- Song, F.; Hurtado del Pozo, C.; Rosario, R.; Zou, Y.S.; Ananthakrishnan, R.; Xu, X.; Patel, P.R.; Benoit, V.M.; Yan, S.F.; Li, H.; et al. RAGE Regulates the Metabolic and Inflammatory Response to High-Fat Feeding in Mice. Diabetes 2014, 63, 1948–1965. [Google Scholar] [CrossRef] [Green Version]
- Cinti, S.; Mitchell, G.; Barbatelli, G.; Murano, I.; Ceresi, E.; Faloia, E.; Wang, S.; Fortier, M.; Greenberg, A.S.; Obin, M.S. Adipocyte death defines macrophage localization and function in adipose tissue of obese mice and humans. J. Lipid Res. 2005, 46, 2347–2355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujiya, A.; Nagasaki, H.; Seino, Y.; Okawa, T.; Kato, J.; Fukami, A.; Himeno, T.; Uenishi, E.; Tsunekawa, S.; Kamiya, H.; et al. The role of S100B in the interaction between adipocytes and macrophages. Obesity 2013, 22, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, L.; Zhang, S.; Yu, Q.; Xiong, F.; Huang, K.; Wang, C.Y.; Yang, P. HMGB1, an innate alarmin, plays a critical role in chronic inflammation of adipose tissue in obesity. Mole. Cell Endocrinol. 2017, 454, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Bonnie, K.S.; Alyssa, H.H. Macrophage infiltration into adipose tissue initiation, propagation and remodeling. Future Lipidol. 2008, 3, 545–556. [Google Scholar]
- Stern, D.; Yan, S.D.; Yan, S.E.; Schmidt, A.M. Receptor for advanced glycation endproducts: A multiligand receptor magnifying cell stress in diverse pathologic settings. Adv. Drug Deliv. Rev. 2002, 54, 1615–1625. [Google Scholar] [CrossRef]
- Murano, I.; Barbatelli, G.; Parisani, V.; Latini, C.; Muzzonigro, G.; Castellucci, M.; Cinti, S. Dead adipocytes, detected as crown-like structures, are prevalent in visceral fat depots of genetically obese mice. J. Lipid Res. 2008, 49, 1562–1568. [Google Scholar] [CrossRef] [Green Version]
- Fujisaka, S.; Usui, I.; Nawaz, A.; Takikawa, A.; Kado, T.; Igarashi, Y.; Tobe, K. M2 macrophages in metabolism. Diabetol. Int. 2016, 7, 342–351. [Google Scholar] [CrossRef]
- Bray, G.A.; Ryan, D.H. Medical therapy for the patient with obesity. Circulation 2012, 125, 1695–1703. [Google Scholar] [CrossRef]
- Yanovski, S.Z.; Yanovski, J.A. Long-term drug treatment for obesity: A systematic and clinical review. JAMA 2014, 311, 74–86. [Google Scholar] [CrossRef]
- Amiot, M.J.; Riva, C.; Vinet, A. Effects of dietary polyphenols on metabolic syndrome features in humans: Systematic review. Obes. Rev. 2016, 17, 537–586. [Google Scholar] [CrossRef]
- Ding, Y.; Kim, S.H.; Lee, J.J.; Hong, J.T.; Kim, E.A.; Kang, D.H.; Heo, S.J.; Lee, S.H. Anti-melanogenesis activity of Ecklonia cava extract cultured in tanks with magma seawater of Jeju Island. Algae 2019, 34, 177–185. [Google Scholar] [CrossRef]
- Kim, I.H.; Choi, J.W.; Lee, M.K.; Kwon, C.J.; Nam, T.J. Anti-obesity effects of pectinase and cellulase enzyme-treated Ecklonia cava extract in high-fat diet-fed C57BL/6N mice. Int. J. Mol. Med. 2018, 41, 924–934. [Google Scholar] [PubMed]
- Kang, S.M.; Lee, S.H.; Heo, S.J.; Kim, K.N.; Jeon, Y.J. Evaluation of antioxidant properties of a new compound, pyrogallol-phloroglucinol-6,6′-bieckol isolated from brown algae, Ecklonia cava. Nutr. Res. Pract. 2011, 5, 495–502. [Google Scholar] [CrossRef]
- Oh, S.; Son, M.; Lee, H.; Kim, H.S.; Jeon, Y.J.; Byun, K. Protective effect of Pyrogallol-Phloroglucinol-6.6-Bieckol from Ecklonia cava on Monocyte-Associated Vascular Dysfunction. Mar. Drugs 2018, 16, 441. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.S.; Jeon, H.J.; Lee, O.H.; Lee, B.Y. Dieckol, a major phlorotannin in Ecklonia cava, suppresses lipid accumulation in the adipocytes of high-fat diet-fed zebrafish and mice: Inhibition of early adipogenesis via cell-cycle arrest and AMPKα activation. Mol. Nutr. Food Res. 2015, 59, 1458–1471. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.C.; Lee, M.; Lee, J.H.; Lee, S.H.; Lim, Y.; Jeon, Y.J. Dieckol, a phlorotannin isolated from a brown seaweed, Ecklonia cava, inhibits adipogenesis through AMP-activated protein kinase (AMPK) activation in 3T3-L1 preadipocytes. Environ. Toxicol. Pharmacol. 2013, 36, 1253–1260. [Google Scholar] [CrossRef] [PubMed]
- Gaens, K.H.; Stehouwer, C.D.; Schalkwijk, C.G. Advanced glycation endproducts and its receptor for advanced glycation endproducts in obesity. Curr. Opin. Lipidol. 2013, 24, 4–11. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, X.; Jiang, G.; Yu, Y.; Wu, J.; Su, Y.; Sun, A.; Zou, Y.; Jiang, H.; Ge, J. HMGB1 enhances mechanical stress-induced cardiomyocyte hypertrophy in vitro via RAGE/ERK1/2 signaling pathway. Int. J. Mol. Med. 2019, 44, 885–892. [Google Scholar] [CrossRef]
- Monden, M.; Koyama, H.; Otsuka, Y.; Morioka, T.; Mori, K.; Shoji, T.; Mima, Y.; Motoyama, K.; Fukumoto, S.; Shioi, A.; et al. Receptor for Advanced Glycation End Products Regulates Adipocyte Hypertrophy and Insulin Sensitivity in Mice. Diabetes 2013, 62, 478–489. [Google Scholar] [CrossRef] [Green Version]
- Kierdorf, K.; Fritz, G. RAGE regulation and signaling in inflammation and beyond. J. Leukoc. Biol. 2013, 94, 55–68. [Google Scholar] [CrossRef]
- Shakambari, G.; Balasubramaniem, A.K.; Varalakshmi, P. Phlorotannins from Brown Algae: Inhibition of advanced glycation end products formation in high glucose induced Caenorhabditis elegans. Indian J. Exp. Biol. 2015, 53, 371–379. [Google Scholar] [PubMed]
- Li, Y.X.; Kim, S.K. Utilization of seaweed derived ingredients as potential antioxidants and functional ingredients in the food industry: An overview. Food Sci. Biot. 2011, 20, 1461–1466. [Google Scholar] [CrossRef]
- Thomas, D.; Apovian, C. Macrophage functions in lean and obese adipose tissue. Metabolism 2017, 72, 120–143. [Google Scholar] [CrossRef] [PubMed]
- Lumeng, C.N.; DelProposto, J.B.; Westcott, D.J.; Saltiel, A.R. Phenotypic Switching of Adipose Tissue Macrophages with Obesity Is Generated by Spatiotemporal Differences in Macrophage Subtypes. Diabetes 2008, 57, 3239–3246. [Google Scholar] [CrossRef]
- Oh, S.Y.; Ahn, H.S.; Park, H.J.; Lee, J.I.; Park, K.Y.; Hwang, D.H.; Lee, S.J.; Son, K.H.; Byun, K.H. The attenuating effects of pyridoxamine on adipocyte hypertrophy and inflammation differ by adipocyte location. J. Nutr. Biochem. 2019, 72, 108173. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Yamamoto, H. RAGE-Mediated Inflammation, Type 2 Diabetes, and Diabetic Vascular Complication. Front. Endocrinol. 2013, 21, 105. [Google Scholar] [CrossRef]
- Subramanian, S.; Pallati, P.K.; Sharma, P.; Agrawal, D.K.; Nandipati, K.C. Significant association of TREM-1 with HMGB1, TLRs and RAGE in the pathogenesis of insulin resistance in obese diabetic populations. Am. J. Transl. Res. 2017, 9, 3224–3244. [Google Scholar]
- Coats, B.R.; Schoenfelt, K.Q.; Barbosa-lorenzi, V.C.; Peris, E.; Cui, C.; Hoffman, A.; Zhou, G.; Fernandez, S.; Zhai, L.; Hall, B.A.; et al. Metabolically Activated Adipose Tissue Macrophages Perform Detrimental and Beneficial Functions during Diet-Induced Obesity. Cell Rep. 2017, 20, 3149–3161. [Google Scholar] [CrossRef] [Green Version]
- Pierine, D.T.; Navarro, M.E.L.; Minatel, O.; Luvizotto, R.A.M.; Nascimento, A.F.; Ferreira, A.L.A.; Yeum, K.J.; Correa, C.R. Lycopene supplementation reduces TNF-αvia RAGE in the kidney of obese rats. Nutr. Diabetes 2014, 4, e142. [Google Scholar] [CrossRef]
- Lesna, K.; Cejkova, S.; Kralova, A.; Fronek, J.; Petras, M.; Sekerkova, A.; Thieme, F.; Janousek, L.; Poledne, R. Human adipose tissue accumulation is associated with pro-inflammatory changes in subcutaneous rather than visceral adipose tissue. Nutr. Diabetes 2017, 7, e264. [Google Scholar] [CrossRef]
- Ramasmy, R.; Yan, S.E.; Herold, K.; Clynes, R.; Schmidt, A.M. Receptor for advanced glycation end products: Functional roles in the inflammatory response: Winding the way to the pathogenesis of endothelial dysfunction and atherosclerosis. Ann. N. Y. Acad. Sci. 2008, 1126, 7. [Google Scholar] [CrossRef] [PubMed]
- Fontana, L.; Eagon, J.C.; Trujilo, M.E.; Scherer, P.E.; Klein, S. Visceral fat adipokine secretion is associated with systemic inflammation in obese humans. Diabetes 2007, 56, 1010–1013. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Chen, R.; Wang, H.; Liang, F. Mechanisms Linking Inflammation to Insulin Resistance. Int. J. Endocrinol. 2015, 2015, 508409. [Google Scholar] [CrossRef] [PubMed]
- Park, K.G.; Park, K.S.; Kim, M.J.; Kim, H.S.; Suh, Y.S.; Ahn, J.D.; Park, K.K.; Chang, Y.C.; Lee, I.K. Relationship between serum adiponectin and leptin concentrations and body fat distribution. Diabetes Res. Clin. Pract. 2004, 63, 135–142. [Google Scholar] [CrossRef]
- Lee, J.H.; Ko, J.Y.; Oh, J.Y.; Kim, C.Y.; Lee, H.J.; Kim, J.; Jeon, Y.J. Preparative isolation and purification of phlorotannins from Ecklonia cava using centrifugal partition chromatography by one-step. Food Chem. 2014, 158, 433–437. [Google Scholar] [CrossRef]
- McGowan, M.H.; Artiss, J.D.; Strandbergh, D.R.; Zak, B. A peroxidase-coupled method for the colorimetric determination of serum triglycerides. Clin. Chem. 1983, 29, 538–542. [Google Scholar]
- Penumarthy, S.; Penmetsa, G.S.; Mannem, S. Assessment of serum levels of triglycerides, total cholesterol, high-density lipoprotein cholesterol, and low-density lipoprotein cholesterol in periodontitis patients. J. Indian Soc. Periodontol. 2013, 17, 30–35. [Google Scholar]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, J.; Oh, S.; Son, M.; Byun, K. Pyrogallol-Phloroglucinol-6,6-Bieckol Alleviates Obesity and Systemic Inflammation in a Mouse Model by Reducing Expression of RAGE and RAGE Ligands. Mar. Drugs 2019, 17, 612. https://doi.org/10.3390/md17110612
Choi J, Oh S, Son M, Byun K. Pyrogallol-Phloroglucinol-6,6-Bieckol Alleviates Obesity and Systemic Inflammation in a Mouse Model by Reducing Expression of RAGE and RAGE Ligands. Marine Drugs. 2019; 17(11):612. https://doi.org/10.3390/md17110612
Chicago/Turabian StyleChoi, Junwon, Seyeon Oh, Myeongjoo Son, and Kyunghee Byun. 2019. "Pyrogallol-Phloroglucinol-6,6-Bieckol Alleviates Obesity and Systemic Inflammation in a Mouse Model by Reducing Expression of RAGE and RAGE Ligands" Marine Drugs 17, no. 11: 612. https://doi.org/10.3390/md17110612