A Synthetic Derivative of Antimicrobial Peptide Holothuroidin 2 from Mediterranean Sea Cucumber (Holothuria tubulosa) in the Control of Listeria monocytogenes

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Results
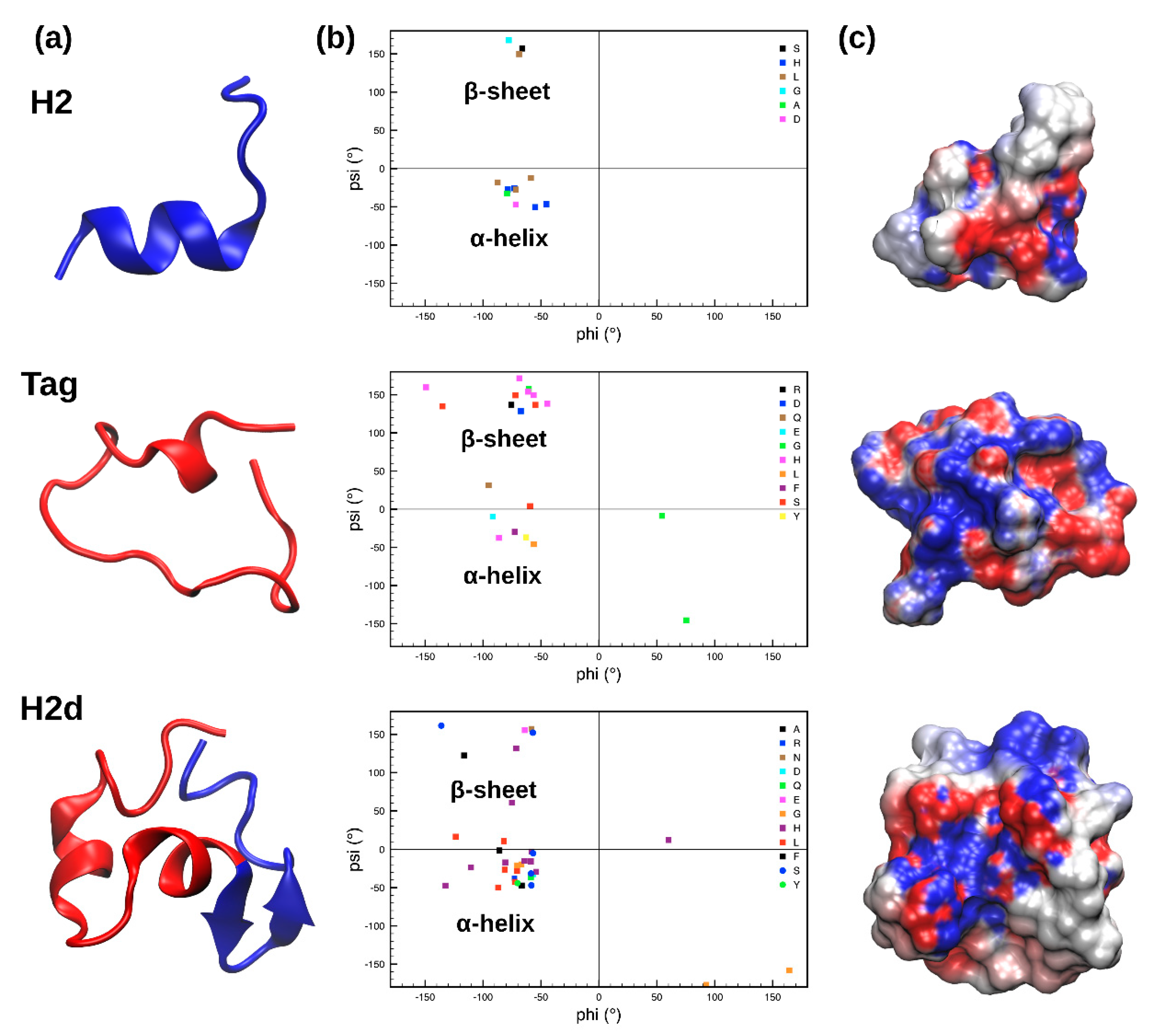
2.1. Molecular Dynamics Simulations
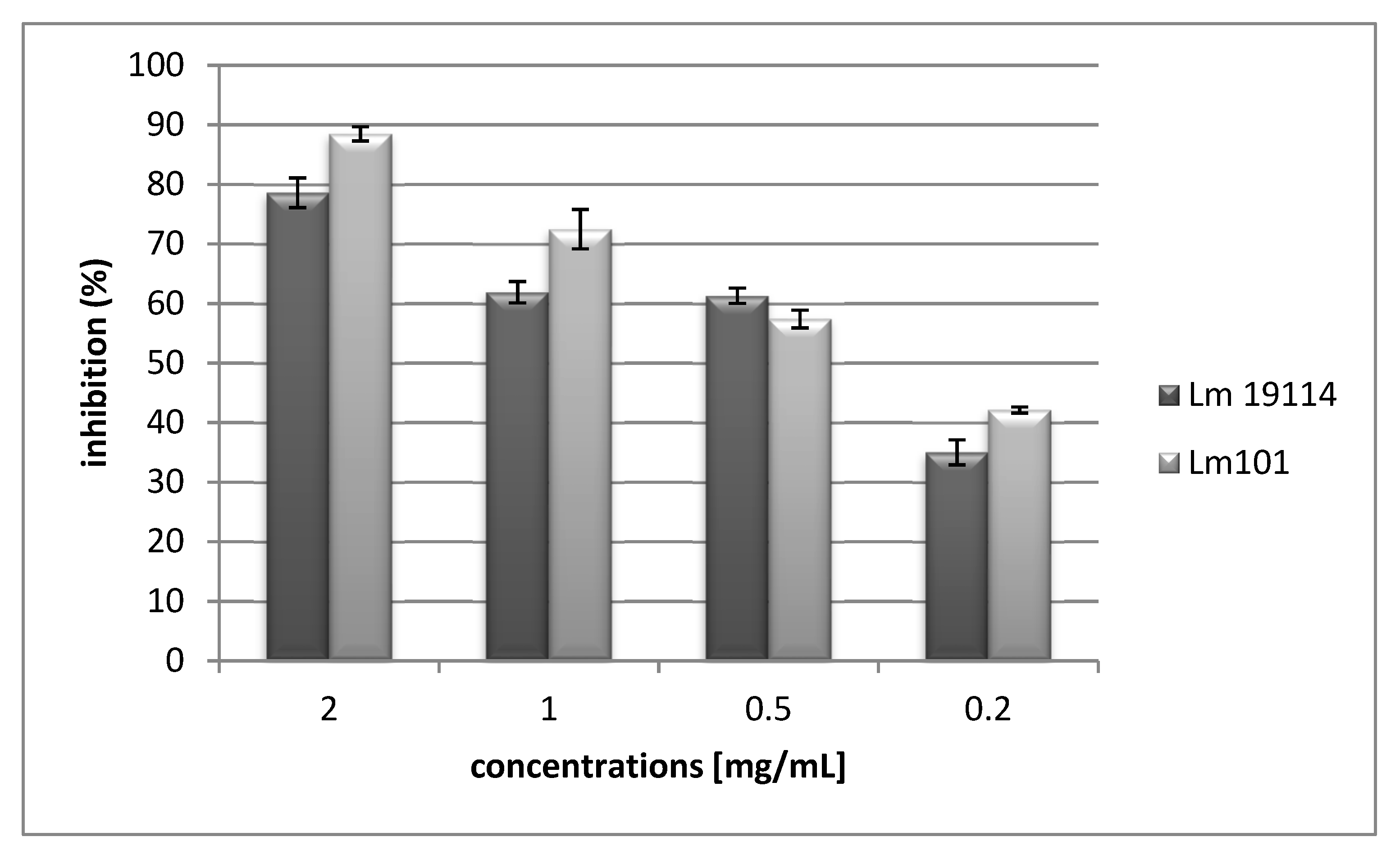
2.2. Anti-Listeria Activity In Vitro against Planktonic and Biofilm Growth
3. Discussion
4. Materials and Methods
4.1. In Silico Analysis
4.2. Molecular Dynamics Simulations
4.3. Peptides Synthesis
4.4. Minimum Inhibitory Concentration Determination
4.5. Evaluation of Biofilm Formation
4.6. Inhibition of Biofilm Formation (Crystal Violet Method)
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Schillaci, D.; Spano, V.; Parrino, B.; Carbone, A.; Montalbano, A.; Barraja, P.; Diana, P.; Cirrincione, G.; Cascioferro, S. Pharmaceutical approaches to target antibiotic resistance mechanisms. J. Med. Chem. 2017, 60, 8268–8297. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.-K.; Kim, C.; Seo, C.H.; Park, Y. The therapeutic applications of antimicrobial peptides (AMPs): A patent review. J. Microbiol. 2017, 55, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Donlan, R.M. Biofilm formation: A clinically relevant microbiological process. Clin. Infect. Dis. 2001, 33, 1387–1392. [Google Scholar] [CrossRef] [PubMed]
- Parrino, B.; Schillaci, D.; Carnevale, I.; Giovannetti, E.; Diana, P.; Cirrincione, G.; Cascioferro, S. Synthetic small molecules as anti-biofilm agents in the struggle against antibiotic resistance. Eur. J. Med. Chem. 2019, 161, 154–178. [Google Scholar] [CrossRef] [PubMed]
- Cascioferro, S.; Cusimano, M.G.; Schillaci, D. Antiadhesion agents against Gram-positive pathogens. Future Microbiol. 2014, 9, 1209–1220. [Google Scholar] [CrossRef] [PubMed]
- Cascioferro, S.; Maggio, B.; Raffa, D.; Raimondi, M.V.; Cusimano, M.G.; Schillaci, D.; Manachini, B.; Plescia, F.; Daidone, G. Synthesis and biofilm formation reduction of pyrazole-4-carboxamide derivatives in some Staphylococcus aureus strains. Eur. J. Med. Chem. 2016, 123, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Carbone, A.; Parrino, B.; Cusimano, M.G.; Spano, V.; Montalbano, A.; Barraja, P.; Schillaci, D.; Cirrincione, G.; Diana, P.; Cascioferro, S. New Thiazole Nortopsentin Analogues Inhibit Bacterial Biofilm Formation. Mar. Drugs 2018, 16, 274. [Google Scholar] [CrossRef] [PubMed]
- Li, X.H.; Lee, J.H. Antibiofilm agents: A new perspective for antimicrobial strategy. J. Microbiol. 2017, 55, 753–766. [Google Scholar] [CrossRef] [PubMed]
- Gopal, R.; Lee, J.H.; Kim, Y.G.; Kim, M.S.; Seo, C.H.; Park, Y. Anti-microbial, anti-biofilm activities and cell selectivity of the NRC-16 peptide derived from witch flounder, Glyptocephalus cynoglossus. Mar. Drugs 2013, 11, 1836–1852. [Google Scholar] [CrossRef] [PubMed]
- Schillaci, D.; Arizza, V.; Parrinello, N.; Di Stefano, V.; Fanara, S.; Muccilli, V.; Cunsolo, V.; Haagensen, J.J.; Molin, S. Antimicrobial and antistaphylococcal biofilm activity from the sea urchin Paracentrotus lividus. J. Appl. Microbiol. 2010, 108, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Schillaci, D.; Cusimano, M.G.; Cunsolo, V.; Saletti, R.; Russo, D.; Vazzana, M.; Vitale, M.; Arizza, V. Immune mediators of sea-cucumber Holothuria tubulosa (Echinodermata) as source of novel antimicrobial and anti-staphylococcal biofilm agents. AMB Express 2013, 3, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Raftery, M.J.; Waugh, R.J.; Bowie, J.H.; Wallace, J.C.; Tyler, M.J. The structures of the frenatin peptides from the skin secretion of the giant tree frog Litoria infrafrenata. J. Pept. Sci. 1996, 2, 117–124. [Google Scholar] [CrossRef]
- Buchanan, R.L.; Gorris, L.G.M.; Hayman, M.M.; Jackson, T.C.; Whiting, R.C. A review of Listeria monocytogenes: An update on outbreaks, virulence, dose-response, ecology, and risk assessments. Food Control 2017, 75, 1–13. [Google Scholar] [CrossRef]
- Etter, A.J.; Hammons, S.R.; Roof, S.; Simmons, C.; Wu, T.; Cook, P.W.; Katubig, A.; Stasiewicz, M.J.; Wright, E.; Warchocki, S.; et al. Enhanced sanitation standard operating procedures have limited lmpact on Listeria monocytogenes Prevalence in Retail Delis. J. Food Prot. 2017, 15, 1903–1912. [Google Scholar] [CrossRef] [PubMed]
- Gaulin, C.; Currie, A.; Gravel, G.; Hamel, M.; Leblanc, M.A.; Ramsay, D.; Bekal, S. Summary of 11 years of enteric outbreak investigations and criteria to initiate an investigation, Province of Quebec, 2002 through 2012. J. Food. Prot. 2014, 77, 1563–1570. [Google Scholar] [CrossRef] [PubMed]
- Chae, M.S.; Schraft, H.; Truelstrup Hansen, L.; Mackereth, R. Effects of physicochemical surface characteristics of Listeria monocytogenes strains on attachment to glass. Food Microbiol. 2006, 23, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Lal, K.; Pollock, J.J. Histatins 2 and 4 are autoproteolytic degradation products of human parotid saliva. Oral Microbiol. Immunol. 1992, 7, 127–128. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Singla, D.; Rashid, M.; Raghava, G.P. Designing of peptides with desired half-life in intestine-like environment. BMC Bioinformatics 2014, 15, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Lamiable, A.; Thevenet, P.; Rey, J.; Vavrusa, M.; Derreumaux, P.; Tuffery, P. PEP-FOLD3: Faster de novo structure prediction for linear peptides in solution and in complex. Nucleic Acids Res. 2016, 44, W449–W454. [Google Scholar] [CrossRef] [PubMed]
- Camproux, A.C.; Gautier, R.; Tuffery, P. A hidden markov model derived structural alphabet for proteins. J. Mol. Biol. 2004, 339, 591–605. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yang, Y.; Zhan, J.; Dai, L.; Zhou, Y. Energy functions in de novo protein design: Current challenges and future prospects. Annu. Rev. Biophys. 2013, 42, 315–335. [Google Scholar] [CrossRef] [PubMed]
- Poyart-Salmeron, C.; Carlier, C.; Trieu-Cuot, P.; Courtieu, A.L.; Courvalin, P. Transferable plasmid-mediated antibiotic resistance in Listeria monocytogenes. Lancet 1990, 335, 1422–1426. [Google Scholar] [CrossRef]
- Qu, X.D.; Lehrer, R.I. Secretory phospholipase A2 is the principal bactericide for staphylococci and other gram-positive bacteria in human tears. Infect. Immun. 1998, 66, 2791–2797. [Google Scholar] [PubMed]
- Charpentier, E.; Gerbaud, G.; Jacquet, C.; Rocourt, J.; Courvalin, P. Incidence of antibiotic resistance in Listeria species. J. Infect. Dis. 1995, 172, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Aureli, P.; Ferrini, A.M.; Mannoni, V.; Hodzic, S.; Wedell-Weergaard, C.; Oliva, B. Susceptibility of Listeria monocytogenes isolated from food in Italy to antibiotics. Int. J. Food Microbiol. 2003, 83, 325–330. [Google Scholar] [CrossRef]
- Li, Q.; Sherwood, J.S.; Logue, C.M. Antimicrobial resistance of Listeria spp. recovered from processed bison. Lett. Appl. Microbiol. 2007, 44, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Spinello, A.; Cusimano, M.G.; Schillaci, D.; Inguglia, L.; Barone, G.; Arizza, V. Antimicrobial and Antibiofilm Activity of a Recombinant Fragment of beta-Thymosin of Sea Urchin Paracentrotus lividus. Mar. Drugs 2018, 16, 366. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.G.; Hahm, K.S.; Park, Y.; Kim, H.Y.; Lee, W.; Lim, S.C.; Seo, Y.K.; Choi, C.H. Functional and structural characteristics of anticancer peptide Pep27 analogues. Cancer Cell Int. 2005, 5, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Rydengard, V.; Shannon, O.; Lundqvist, K.; Kacprzyk, L.; Chalupka, A.; Olsson, A.K.; Morgelin, M.; Jahnen-Dechent, W.; Malmsten, M.; Schmidtchen, A. Histidine-rich glycoprotein protects from systemic Candida infection. PLoS Pathog. 2008, 4, e1000116. [Google Scholar] [CrossRef] [PubMed]
- Mahlapuu, M.; Hakansson, J.; Ringstad, L.; Bjorn, C. Antimicrobial Peptides: An Emerging Category of Therapeutic Agents. Front. Cell Infect. Microbiol. 2016, 6, 194–208. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.D.; Won, H.S.; Kim, J.H.; Mishig-Ochir, T.; Lee, B.J. Antimicrobial peptides for therapeutic applications: A review. Molecules 2012, 17, 12276–12286. [Google Scholar] [CrossRef] [PubMed]
- Sinde, E.; Carballo, J. Attachment of Salmonella spp. and Listeria monocytogenes to stainless steel, rubber and polytetrafluorethylene: The influence of free energy and the effect of commercial sanitizers. Food Microbiol. 2000, 17, 439–447. [Google Scholar] [CrossRef]
- Leriche, V.; Carpentier, B. Limitation of adhesion and growth of Listeria monocytogenes on stainless steel surfaces by Staphylococcus sciuri biofilms. J. Appl. Microbiol. 2000, 88, 594–605. [Google Scholar] [CrossRef] [PubMed]
- Chae, M.S.; Schraft, H. Cell viability of Listeria monocytogenes biofilms. Food Microbiol. 2001, 18, 103–112. [Google Scholar] [CrossRef]
- Chae, M.S.; Schraft, H. Comparative evaluation of adhesion and biofilm formation of different Listeria monocytogenes strains. Int. J. Food Microbiol. 2000, 62, 103–111. [Google Scholar] [CrossRef]
- Wong, A.C. Biofilms in food processing environments. J. Dairy Sci. 1998, 81, 2765–2770. [Google Scholar] [CrossRef]
- Hood, S.K.; Zottola, E.A. Adherence to stainless steel by foodborne microorganisms during growth in model food systems. Int. J. Food Microbiol. 1997, 37, 145–153. [Google Scholar] [CrossRef]
- Joseph, B.; Otta, S.K.; Karunasagar, I.; Karunasagar, I. Biofilm formation by Salmonella spp. on food contact surfaces and their sensitivity to sanitizers. Int. J. Food Microbiol. 2001, 64, 367–372. [Google Scholar] [CrossRef]
- Bower, C.K.; Daeschel, M.A. Resistance responses of microorganisms in food environments. Int. J. Food Microbiol. 1999, 50, 33–44. [Google Scholar] [CrossRef]
- Lee, I.H.; Zhao, C.; Cho, Y.; Harwig, S.S.; Cooper, E.L.; Lehrer, R.I. Clavanins, alpha-helical antimicrobial peptides from tunicate hemocytes. FEBS Lett. 1997, 400, 158–162. [Google Scholar] [CrossRef]
- Iijima, R.; Kisugi, J.; Yamazaki, M. A novel antimicrobial peptide from the sea hare Dolabella auricularia. Dev. Comp. Immunol. 2003, 27, 305–311. [Google Scholar] [CrossRef]
- Maupetit, J.; Derreumaux, P.; Tuffery, P. A fast method for large-scale de novo peptide and miniprotein structure prediction. J. Comput. Chem. 2010, 31, 726–738. [Google Scholar] [PubMed]
- Schillaci, D.; Cusimano, M.G.; Spinello, A.; Barone, G.; Russo, D.; Vitale, M.; Parrinello, D.; Arizza, V. Paracentrin 1, a synthetic antimicrobial peptide from the sea-urchin Paracentrotus lividus, interferes with staphylococcal and Pseudomonas aeruginosa biofilm formation. AMB Express 2014, 4, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Schillaci, D.; Spinello, A.; Cusimano, M.G.; Cascioferro, S.; Barone, G.; Vitale, M.; Arizza, V. A peptide from human beta thymosin as a platform for the development of new anti-biofilm agents for Staphylococcus spp. and Pseudomonas aeruginosa. World J. Microbiol. Biotechnol. 2016, 32, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Case, D.A.; Brozell, S.R.; Cerutti, D.S.; Cheatham, T.E.; Cruzeiro, V.W.D.; Darden, T.A.; Duke, R.E.; Ghoreishi, D.; Gilson, M.K.; Gohlke, H.; et al. AMBER 2018; University of California: San Francisco, CA, USA, 2018. [Google Scholar]
- Lindorff-Larsen, K.; Piana, S.; Palmo, K.; Maragakis, P.; Klepeis, J.L.; Dror, R.O.; Shaw, D.E. Improved side-chain torsion potentials for the Amber ff99SB protein force field. Proteins 2010, 78, 1950–1958. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Davidchack, R.L.; Ouldridge, T.E.; Tretyakov, M.V. New Langevin and gradient thermostats for rigid body dynamics. J. Chem. Phys. 2015, 142, 144114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berendsen, H.J.C.; Postma, J.P.M.; Vangunsteren, W.F.; Dinola, A.; Haak, J.R. Molecular-Dynamics with coupling to an external Bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Dolinsky, T.J.; Nielsen, J.E.; McCammon, J.A.; Baker, N.A. PDB2PQR: An automated pipeline for the setup of Poisson-Boltzmann electrostatics calculations. Nucleic Acids Res. 2004, 32, W665–W667. [Google Scholar] [CrossRef] [PubMed]
- Mauro, N.; Schillaci, D.; Varvara, P.; Cusimano, M.G.; Geraci, D.M.; Giuffre, M.; Cavallaro, G.; Maida, C.M.; Giammona, G. Branched high molecular weight glycopolypeptide with broad-spectrum antimicrobial activity for the treatment of biofilm related infections. ACS Appl. Mater. Interfaces 2018, 10, 318–331. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Peptide | Sequence |
|---|---|
| H2 | ASHLGHHALDHLLK |
| Tag | MRGSHHHHHHGSSGENLYFQSL |
| H2d | MRGSHHHHHHGSSGENLYFQSLASHLGHHALDHLLK |
| Chemical Physical Properties | Peptides | ||
|---|---|---|---|
| H2 | Tag | H2d | |
| Theoretical mass (Da) | 1548.7 | 2555.7 | 4086.5 |
| Isoelectric point | 6.68 | 6.37 | 6.41 |
| Hydrophobic ratio | 42% | 18% | 27% |
| Protein-binding potential Boman index (kcal/mol) | 0.86 | 2.61 | 1.93 |
| Half-life (s) | 1.13 | 0.73 | 0.64 |
| Minimum Inhibitory Concentration (MIC)mg/mL and [mM] | |||||||
|---|---|---|---|---|---|---|---|
| Peptides | L. monocytogenes Strains | ||||||
| ATCC19114 | ATCC19115 | ATCC7644 | NCTC18890 | 89 | 101 | 103 | |
| H2 | >5 [3.2] | 5 [3.2] | >5 [3.2] | >5 [3.2] | >5 [3.2] | >5 [3.2] | >5 [3.2] |
| Tag | >5 [2.0] | >5 [2.0] | >5 [2.0] | >5 [2.0] | >5 [2.0] | >5 [2.0] | >5 [2.0] |
| H2d | 2.5 [0.6] | 2.5 [0.6] | 2.5 [0.6] | 5 [1.2] | 1.2 [0.3] | 2.5 [0.6] | 2.5 [0.6] |
| MIC mg/mL and [mM] | |||||||
|---|---|---|---|---|---|---|---|
| Peptides | L. monocytogenes Strains | ||||||
| ATCC19114 | ATCC19115 | ATCC7644 | NCTC18890 | 89 | 101 | 103 | |
| H2 | >5 [3.2] | 5 [3.2] | >5 [3.2] | >5 [3.2] | >5 [3.2] | >5 [3.2] | >5 [3.2] |
| Tag | >5 [2.0] | >5 [2.0] | >5 [2.0] | >5 [2.0] | >5 [2.0] | >5 [2.0] | >5 [2.0] |
| H2d | 2.5 [0.6] | 1.2 [0.3] | 2.5 [0.6] | 5 [1.2] | 2.5 [0.6] | 2.5 [0.6] | 2.5 [0.6] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cusimano, M.G.; Spinello, A.; Barone, G.; Schillaci, D.; Cascioferro, S.; Magistrato, A.; Parrino, B.; Arizza, V.; Vitale, M. A Synthetic Derivative of Antimicrobial Peptide Holothuroidin 2 from Mediterranean Sea Cucumber (Holothuria tubulosa) in the Control of Listeria monocytogenes. Mar. Drugs 2019, 17, 159. https://doi.org/10.3390/md17030159
Cusimano MG, Spinello A, Barone G, Schillaci D, Cascioferro S, Magistrato A, Parrino B, Arizza V, Vitale M. A Synthetic Derivative of Antimicrobial Peptide Holothuroidin 2 from Mediterranean Sea Cucumber (Holothuria tubulosa) in the Control of Listeria monocytogenes. Marine Drugs. 2019; 17(3):159. https://doi.org/10.3390/md17030159
Chicago/Turabian StyleCusimano, Maria Grazia, Angelo Spinello, Giampaolo Barone, Domenico Schillaci, Stella Cascioferro, Alessandra Magistrato, Barbara Parrino, Vincenzo Arizza, and Maria Vitale. 2019. "A Synthetic Derivative of Antimicrobial Peptide Holothuroidin 2 from Mediterranean Sea Cucumber (Holothuria tubulosa) in the Control of Listeria monocytogenes" Marine Drugs 17, no. 3: 159. https://doi.org/10.3390/md17030159