Oceans as a Source of Immunotherapy


Department of Molecular Science and Technology, Ajou University, Suwon 16499, Korea
*
Author to whom correspondence should be addressed.
Mar. Drugs 2019, 17(5), 282; https://doi.org/10.3390/md17050282
Submission received: 30 March 2019
/
Revised: 3 May 2019
/
Accepted: 6 May 2019
/
Published: 10 May 2019
(This article belongs to the Special Issue Marine Immunomodulators)
Abstract
:Marine flora is taxonomically diverse, biologically active, and chemically unique. It is an excellent resource, which offers great opportunities for the discovery of new biopharmaceuticals such as immunomodulators and drugs targeting cancerous, inflammatory, microbial, and fungal diseases. The ability of some marine molecules to mediate specific inhibitory activities has been demonstrated in a range of cellular processes, including apoptosis, angiogenesis, and cell migration and adhesion. Immunomodulators have been shown to have significant therapeutic effects on immune-mediated diseases, but the search for safe and effective immunotherapies for other diseases such as sinusitis, atopic dermatitis, rheumatoid arthritis, asthma and allergies is ongoing. This review focuses on the marine-originated bioactive molecules with immunomodulatory potential, with a particular focus on the molecular mechanisms of specific agents with respect to their targets. It also addresses the commercial utilization of these compounds for possible drug improvement using metabolic engineering and genomics.
1. Introduction
Immune system dysfunction leads to the development of allergies, autoimmune and chronic inflammatory diseases, and cancers. Inflammation has been suggested to be the principal cause of chronic illnesses such as obesity, diabetes, cancer, rheumatoid arthritis (RA), neurodegenerative, and autoimmune diseases (ADs). The hallmark of autoimmunity is chronic inflammation that leads to the release of pro-inflammatory cytokines and other mediators, known as danger-associated molecular patterns (DAMPS), that activate pathogen recognition receptors (PRR) expressed by immune cells. Autoantibodies recognize these DAMPs and activate myeloid immune cells with an enhanced inflammatory response, leading to exacerbation of the condition. This self-perpetuating cycle continues, in order to assist with injury repair [1,2,3].
Recent estimates suggest that 7.6–9.4% of the world’s population is affected by immune-mediated diseases. Such diseases include inflammatory bowel disease (IBD), type 1 diabetes mellitus (TIDM), and RA. Women are up to ten times as likely to be sufferers as men [4]. ADs are among the ten leading causes of death for women, and affect them disproportionately in every age group up to 75 years of age [5]. According to the NIH report, around 23.5 million Americans have ADs, while the American Autoimmune Related Diseases Association (AARDA) puts this figure at 50 million. More than 80 different ADs have been identified and 40 additional diseases are suspected as being ADs [6,7]. The holy grail of immunotherapy is the reprograming of the immune system to maintain or restore homeostasis, and there is an urgent need to develop such drugs.
The search for novel immunomodulators is challenging, despite the existence of considerable amounts of demographic and epidemiological data about ADs. Questions about how autoimmunity is triggered and how self-tolerance is broken down remain to be fully answered. Nevertheless, progression in our understanding of the pathophysiology of ADs has led to the possibility of identifying new drug targets and new effective compounds with significant therapeutic potential. Several drug discovery and development programs are focused on the search for bioactive compounds obtained from natural sources. The study of nature’s enormous arsenal of new bioactive compounds and natural metabolites has historically led to immense benefits with respect to drug discovery [8]. The bioactivity of marine-derived natural products is significantly higher than that of compounds of terrestrial origin. For instance, in a National Cancer Institute preclinical cytotoxicity screen, approximately 1% of the marine samples tested showed anti-tumor potential in comparison with 0.1% of the terrestrial samples tested [9].
Oceans cover three quarters of the Earth’s surface, and contain the world’s greatest biodiversity, with at least 300,000 species of animals and plants described to date [10,11]. The diverse environments of oceanic zones provide a rich source of organisms. The hostile and competitive oceanic environment, with its high salt concentration and variations in hydrostatic pressure, produces microorganisms which are resistant to various kinds of stress, leading to the production of unique biomolecules. This enormous source of marine microorganisms has been exploited as a source of medicine since ancient times. The oceans are an exceptional storehouse of structurally and chemically novel bioactive compounds with unique biological features that are not generally found in terrestrial natural products. Over 60% of the active compounds of marketed formulations are natural products or their synthetic derivatives or mimics [12]. Nearly 3000 new compounds from marine sources have been identified over the last few years, and some have entered clinical trials [13]. Due to improved technologies for deep-sea sample collection and large-scale drug production through aquaculture and drug synthesis, the extent of this activity has been increasing steadily. Advanced engineering and the use of new technologies such as scuba diving techniques, remotely operated vehicles (ROVs) and manned submersibles have opened up the scientific exploration of the marine environment [14,15,16].
Marine compounds have been shown to have significant activity in antiviral, analgesic, antitumor, immunomodulatory, and anti-inflammatory assays [17,18]. Currently, six out of nine drugs in the market for cancer treatment are of marine origin, while several more are under clinical trials [18]. Undoubtedly, in the area of cancer, the marine metabolic arsenal plays a dominant role.
2. Oceanic Sources of Immunotherapy
Immunotherapies induce, enhance or suppress an immune response to ameliorate a pathological condition. Immune response activation therapies are used in vaccines for conditions such as cancer, while immune response suppression therapies can be used to prevent graft rejection and treat autoimmune conditions and allergies. The metabolic products of microbes, phytoplankton, and zooplanktons obtained from marine environments offer a great diversity of physical and chemical attributes. Exploitation of marine microorganisms, which are the source of new genes, has led to the discovery of new drugs and targets.
2.1. Oceanic Bacteria
Marine bacteria synthesize a wide range of valuable compounds with potential pharmaceutical applications [19,20]. Bacteria have yielded novel anti-inflammatory agents such as pseudopterosins, topsentins, scytonemin, and manoalide, anticancer and immunomodulatory agents including sarcodictylin eleutherobin, bryostatins and discodermolide, and antibiotics like marinone. Lactobacilli stimulate and modulate the mucosal immune system by reducing the production of pro-inflammatory cytokines through their actions on nuclear factor-κB (NF-κB) pathways. This effect is mediated by the production of anti-inflammatory cytokine interleukin-10 (IL-10) and host defense peptide β–defensin 2. The production of immunoglobulin A (IgA), dendritic cell maturation, and cell apoptosis is enhanced in response to short chain fatty acids [21]. Three diketopiperazine derivatives, cyclo(L-Pro-D-Val), cyclo(L-Pro-L-Tyr), and cyclo(L-pro-D-Leu), have been derived from two marine bacteria, Bacillus sp. HC001 and Piscicoccus sp. 12L081. These compounds show anti-inflammatory effects by suppressing polyphosphate mediated septic responses and hyper-permeability through the inhibition of p38 mitogen-activated protein kinases (MAPKs) activation. Downregulation of tumor necrosis factor (TNF-α), IL-6, NF-κB and ERK1/2 was also observed after administration of these three compounds [22]. A novel exopolysaccharide (EPS) EPS1-T14, a water-soluble non-toxic exopolymer obtained from the marine bacterium Bacillus licheniformis, is able to stimulate an immune response. EPS1-T14 exhibits antiviral activity, as it inhibits the replication of HSV-2 in human peripheral blood mononuclear cells (hPBMCs). EPS1-T14 also stimulates the Th1 cell-mediated immune response [23]. Another EPS, TA-1, isolated from the thermophilic marine bacterium, Thermus aquaticus, is the strongest candidate for the EPS-binding receptor such as toll-like receptors (TLRs). TA-1 stimulates the release of the proinflammatory cytokines TNF-α and IL-6 from murine macrophages via a TLR-2 mediated pathway [24]. Prodigiosin (Figure 1, 1) derived from marine bacteria such as Pseudoalteromonas denitrificans, Vibro psychroerythrus, and Serratia marcescens, has a strong inhibitory effect on many protozoan, fungal, and bacterial species, and induces apoptosis in cancer cell lines, as observed by the development of characteristic DNA laddering and apoptotic bodies [25,26]. Cycloprodigisin, a stable analog of prodigiosin (Figure 1, 1) from Pseudoalteromonas dentrificans inhibits TNF-α induced NF-κB activation, as determined through luciferase assay. This NF-κB-inhibitory effect of cycloprodigiosin was retained under multiple stimuli in HeLa, U373, and COS7 cell lines [27,28,29]. Some representative immunomodulatory and anti-inflammatory chemical constituents isolated from marine bacteria are listed in Table 1.
2.2. Cyanobacteria
Cyanobacteria are a fascinating and versatile, biologically important group of bacteria. Cyanobacteria unquestionably constitute one of the richest sources of novel and existing bioactive compounds, including toxins with a wide range of pharmaceutical applications. Cyanobacteria-derived bioactive molecules have a broad spectrum of activities, including antibacterial, antiviral, protease inhibition, anticancer and immunomodulatory activity (Table 2) [44]. Marine cyanobacterial lipopolysaccharide (LPS) has the potential to act as a TLR4 antagonist [45]. A cyanobacterial LPS (CyP), isolated from Oscillatoria planktothrix FP1 shows antagonism to TLR4 activation and exhibits protective effects in inflammatory conditions. CyP acts as a potential inhibitor of the LPS-induced inflammatory response in human and mouse dendritic cells, inhibiting both the MyD88-dependent and MyD88-independent TLR4 signaling pathways. CyP completely inhibits LPS-induced IL-1β, TNF-α, and IL-8 production [46,47]. CyP, when co-incubated with Porphyromonas gingivalis LPS (Pg-LPS) inhibited IL-1β, IL-8, and TNF-α expression more efficiently than LPS. CyP can modulate the Pg-LPS-induced pro-inflammatory response, by blocking TLR4-MD2, and also by preserving miR-146a expression [48]. Malyngamides, a class of compounds derived from the marine cyanobacterium Lyngbya majuscula has potent anti-inflammatory activity. One compound of this class, Malyngamide F (Figure 1, 2) acetate, can inhibit the production of nitric oxide (NO) and other inflammatory biomarkers in RAW 264.7 cells. It selectively inhibits the MyD88-dependent pathway, because LPS stimulation decreases IL-1β and increases TNF-α transcription in MyD88 knockout mice via an MyD88-independent pathway [49]. Polysaccharides that have been extracted from Arthrospira platensis (Spirulina) have been reported to exhibit immune modulation. Immulina is one of these polysaccharides, and has been reported to decrease levels of TNF-α and IL-4 in FcεRI-activated RBL-2H3 cells [50,51]. Potent grassystatin A-C were obtained from the marine cyanobacterium Lyngbya confervoides. In response to exogenous antigen, grassystatin A (Figure 1, 3) significantly reduces the proliferation of T cells. Grassystatin A (Figure 1, 3) inhibit upregulation of IL-17 and interferon-γ (INF-γ) in response to antigen presentation [52]. Some representative immunomodulatory and anti-inflammatory chemical constituents isolated from marine cyanobacteria are listed in Table 2.
2.3. Sponges
Sponges are currently the most important source of biologically active natural marine products and are considered to be a treasure trove of drugs [53,54]. Due to their lack of physical defenses, they have evolved a wide suite of defensive chemicals to deter predators [55]. New biomolecules discovered from marine sponges have strong immunosuppressive activities (Table 3). Didemnins, members of a depsipeptide class of compounds isolated from the Caribbean tunicate Trididemnum solidum, exhibit a variety of biological activities [56]. In particular, didemnin B (Figure 1, 4) is characterized as immunosuppressive, and inhibits lymphocyte activation [57,58]. Target deconvolution studies, which aim to identify the molecular targets of active hits, have revealed that didemnin B (Figure 1, 4) binds to the eukaryotic elongation factor 1α and palmitoyl-protein thioesterase 1. Large amounts of didemnin B (Figure 1, 4) were taken up by proliferating cells, so this compound appears to be a promising drug for cancer treatment or the suppression of activation of the immune system [59].
Dysidea sp. has contributed significantly to biomolecule production [18,60,61]. Polyoxygenated sterols derived from Dysidea sp., have been shown to have strong selective immunosuppressive capability, blocking the interaction between IL-8 and its receptor [62]. Pateamine A (Figure 1, 5) derived from Mycale sp., selectively inhibits the production of IL-2 in the T and B cells that produce the secondary immune response [63,64]. Discodermolide (Figure 1, 6), a unique immunosuppressive and cytotoxic agent, is isolated from Discodermia dissolute, a deep water sponge [65]. An evaluation by the Longley group found that (+)-discodermolide has immunosuppressive properties at low concentrations, both in vivo and in vitro [66]. (+)-discodermolide was found to suppress the two-way mixed lymphocyte reaction in hPBMCs and murine splenocytes.

{kind=link}
{kind=link}
Table 2.
Marine cyanobacteria and their therapeutic chemical constituents.
| Cyanobacteria Species | Chemical(s) | Immunomodulatory Activity | Ref(s) |
|---|---|---|---|
| Oscillatoria planktothrix | CyP | CyP modulates pro-inflammatory effect and inhibits TNF-α, IL-1β and IL-8. | [48] |
| Lyngbya majuscula | Microcolin-A (Peptides) | Suppresses murine splenocytes and inhibits LFA-1 and ICAM-1 mediated cell adhesion. | [49,67] |
| Lyngbya sordida | Malyngamide 2 (lipopeptide) | Inhibits production of NO in LPS-primed RAW 264.7 cells. | [68] |
| Arthrospira platensis | Immolina (Polysaccharide) | Reduces TNF-α and IL-4 levels in RBL-2H3 FcεRI-activated cells. | [50,51] |
| Trichodesmium Erythraeum | Aqueous extract | Anti-inflammatory effects in carrageenan-induced inflammation in rats. | [69] |
| Lyngbya cf. confervoides | Grassystatins A-C | Inhibits presentation of T cell antigen and expression of Cathepsin E, IL-17, and IFN- γ. | [52] |
CyP, cyanobacterial LPS; ICAM-1, intercellular cell adhesion molecule-1; IFN, interferon; IL, interleukin; LFA-1, lymphocyte function-associated molecule-1; TNF, tumour necrosis factor.
Table 3.
Marine sponges and their therapeutic chemical constituents.
| Sponge Species | Chemical(s) | Immunomodulatory Activity | Ref(s) |
|---|---|---|---|
| Plakortis simplex | Simplexides Glycolopids | Inhibits T cell proliferation and induces cytokines and chemokines in a CD1d-dependent manner. | [70,71] |
| Dysidea sp. | Dendroceratida & bolinaquinone (Polyoxygenated sterols) | Inhibits neutrophilic infiltration and IL-1, IL-8, PGE2, COX-2 expression in vivo. | [62,72] |
| Petrosia contignata | Contignasterol (Oxygenated sterol) | Inhibits histamine release in mast cells. | [73] |
| Petrosia sp. | Petrocortyne A (polyacetylenic alcohols) | Inhibits macrophages, reduces the production of TNF-α and the expression of phlogistic infiltration cell factors. | [27,74] |
| Mycale sp. | Pateamine (Thiazole macrolide) | Specifically targets translation initiation factors. Inhibits eIF4A-eIF4G association and promotes stable ternary complex formation between eIF4A and eIF4B. IL-2 inhibitor. | [75,76] |
| Callyspongia sp. | Callyspongidiol (Polyketide) | Dendritic cell activation with enhanced IL-4 and IL-10 production. | [77] |
| Ianthella quadrangulata | Iso-iantheran (Polyketide) | Has implication in tumor or autoimmune diseases. Ionotropic P2Y11 receptor activation. | [78] |
| Xestospongia bergquisita | Xestobergsterol (Polyhidroxylated steroid) | Inhibits the generation of IP3 and PLC activity and intracellular Ca2 + mobilization. | [79] |
| Clathria lissosclera | Clathriols (Polyoxygenated steroids) | Inhibits superoxide production from neutrophils of hPBMCs. | [80] |
| Hyritos sponge | Heteronemin (Sesterterpene) | Inhibits TNF-α induced NF-κB activation and induces caspase-dependent apoptosis in K562 cells. | [81] |
| Xestospongia testudinaria | Methanolic extract | Exhibits anti-inflammatory activity against carrageen-induced paw inflammation. | [82] |
| Plakortis angulospiculatus | Plakortide P | NO inhibition in LPS stimulated macrophages. | [83] |
| Geodia cydonium | Methanolic and Chloroform extraction | Reduces IL-8, CXCL10 and VEGF levels and increases IL-4 and IL-10 levels. | [84,85] |
| Coscinoderma mathewsi | Coscinolactams A-B (Terpenes) & suvanine | PGE2 and NO inhibition in RAW 264.7 cells stimulated by LPS. | [86] |
| Lobophytum crassum | Lobocrassin B | Inhibits LPS-induced BMDC activation by inhibiting TNF-α production. | [87] |
| Petrosaspongia nigra | Petrosaspongiolide | Inhibits chronic inflammation by lowering the production of eicosanoids and TNF-α. | [88] |
| Hyrrios erecta | Puupehedione, dipuupehedione, bispuupehenone | Exhibits cytotoxic and immunomodulatory potential against A-549 human cancer cell line. | [89] |
| Gelliodes fibrosa | Terpenes, steroids and lipids | Ethyl acetate extracts from Gelliodes Fibrosa and Tedania anhelans on in vivo carbon clearance tests showed a moderate immunostimulant effect. | [90] |
| Ircinia variabilis | Fasciculatin (Sesterterpenes) | Exhibits moderate cytotoxicity and no selectivity in the cancer cell lines. | [91] |
| Dendrilla nigra | Lipopolysaccharides & neolamellarins | Dendrilla exhibits enhanced phagocytosis against Escherichia coli. Neolamellarins inhibits HIF-1 activation and VEGF secretion in T47D cells. | [92,93] |
| Theonella swinhoei | Solomonsterol A, perthamides C & D (Peptides) | Theonella peptolides show mild immunosuppressive activity, inhibition of murine hind paw oedema. | [94,95] |
| Discodermia spp | Discodermolide (Polyhydroxylated lactone) | Inhibits murine T cell proliferation and causes cell cycle arrest in gap2 or mitosis phase of human and murine cell lines. | [96] |
| Reniera spp | Cyclic Tripeptide (Renieramide) | In preliminary tests renieramide showed immunomodulating activity. | [97] |
| Trididemnum solidum | Didemnins depsipeptides | Inhibits viral replication in vitro and P388 leukemia in vivo. | |
| Pseudoaxinyssa cantharella | Girolline | Inhibits of IL-8, NF-κB and AP-1 in macrophages derived from THP1. Reduction of IL-8 and IL-6 in primary mononuclear human cells. | [98] |
| Callyspongia siphonella | Callysterol (Sterol) | Potentially inhibits rat hind paw oedema, reduced release of TXB2 from LPS-activated rat brain microglia. | [99] |
| Axinella verrucosa, Acanthella aurantica and Stylissa massa | Alkaloids | Inhibits expression of NF-κB and production of IL-8, IL-2 and TNF-α. | [100] |
| Tedania ignis | Tedanol (Diterpenoid) | Potent anti-inflammatory action to reduce carrageenan-induced mouse paw oedema. Strong inhibition of COX-2 and iNOS expressions. | [101,102] |
| Haliclona sp. | Halipeptins (Depsipeptide) | Strong anti-inflammatory activity, in vivo and in vitro. | [103] |
| Cacospongia mollior | Sesterterpenoid | Suppresses the production of LPS-induced PGE2. | [104] |
| Fascaplysinopsis Bergquist sp. | Fascaplysin (Indole alkaloid) | CDK 4 inhibitor, potential to elicit anti-neuroinflammatory or neuroprotective responses in neuroinflammatory disease models. | [105] |
| Terpios sp. | Terpioside B (Glycolipid) | Inhibits macrophage iNOS expression. | [106] |
AP-1, activator protein; CDK, cyclin-dependent kinase 4; HIF-1, Hypoxia-inducible factor-1; IFN, interferon; IL, interleukin; iNKT, Natural killer T cells with an invariant T cell receptor alpha chain; iNOS, inducible nitric oxide synthase; NF-κB, nuclear factor-κB; PGE2, prostaglandin E2; PLC, phospholipase C; TNF, tumour necrosis factor; TXB2, thromboxane B2; VEGF, Vascular endothelial growth factor.
In several other non-lymphoid cell lines (+)-discodermolide exhibited antiproliferative effects by arresting the cell cycle at G2 and M phase due to microtubule network stabilization [107,108]. Petrosaspongiolide M (Figure 1, 7) isolated from the Caledonian marine sponge Petrosa spongia nigra significantly inhibits chronic inflammation in rats and mice by diminishing eicosanoids and TNF-α production [88]. Petrosaspongiolide M (Figure 1, 7) decreases the NF-κB-DNA binding in response to zymosan in mouse peritoneal macrophages [109]. A marine sesterterpene, heteronemin (Figure 1, 8), isolated from Hyritos sponge species has been found to affect cellular processes including cell cycle and apoptosis, and inhibits TNF-α-induced NF-κB activation [81]. Methanolic extracts of Xestospongia testudinaria, the Red Sea marine sponge, prevent carrageenan-induced acute local inflammation in rats. Malondialdehyde and NO in inflamed rat paws was decreased by this extract, while glutathione, glutathione peroxidase, and catalase activities were increased. It appears to have antioxidant, anti-inflammatory, and immunomodulatory effects [82]. Some representative immunomodulatory and anti-inflammatory chemical constituents isolated from marine sponges are listed in Table 3.
2.4. Algae
Marine algae are rich sources of vitamins, minerals, essential amino acids, lipids, fatty acids, dietary fiber, and polysaccharides [110,111,112,113,114]. Bioactivity studies of marine algae have revealed numerous health-promoting effects including anticoagulant, antibacterial, anti-hypolipidemic, anti-hypertensive, antioxidant, anticancer, and immunomodulatory activities [115]. Fucoidans (Figure 1, 9) from the brown algae Laminaria cichorioides, Laminaria japonica and Fucus evanescens specifically interact with TLRs in vitro, causing the activation of NF-κB via the MyD88 and TRIF-signaling pathways [116]. Brown algae-derived fucoidans activate genes that are responsible for cytokine synthesis, exhibiting pronounced immunotropic activity ex vivo and promoting defense against various pathogens [117]. Sugariura et al. showed that a diet including dried Eisenia arborea powder reduced serum IgE levels and shifted the Th1/Th2 balance by suppressing the release of Th2-type cytokines IL-4 and IL-10 and enhancing the expression of Th1 and IFN-γ in rat spleen and mesenteric lymph node-derived lymphocytes [118]. The red alga Gracilaria verrucosa has anticancer and antioxidant properties. The two enone fatty acids (E)-9-Oxooctadec-10-enoic-acid (Figure 1, 10) and (E)-10-Oxooctadec-8-enoic-acid (Figure 1, 11) isolated from Gracilaria verrucosa inhibit the production of inflammatory biomarkers including NO, IL-6 and TNF-α by suppressing the nuclear translocation of NF-κB and phosphorylation of STAT1 in LPS-stimulated RAW 264.7 cells [119]. Lectins from marine algae Solieria filiformis, Caulerpa cupressoides and Pterocladiella capillacea demonstrate anti-inflammatory effects by enhancing IL-10 and IL-6 formation without affecting the IFN-γ and IL-12 production in murine splenocytes [120]. Some representative immunomodulatory and anti-inflammatory chemical constituents isolated from marine algae are listed in Table 4.
2.5. Marine Fungi
Recent developments in marine mycology have led to a large amount of research into natural products from substrate-insulated fungi in various marine habitats [121]. The discovery rate of novel marine-derived natural products from fungi increased exponentially over the period from 1970 to 2010 [121]. Cyclosporins (Figure 1, 12) are produced by species of fungi including Tolypocladium inflatum gams [122], Neocosmospora vasinfecta, Verticillium spp. and Microdochium nivale [123,124]. Owing to their potent immune-modulation properties, cyclosporins (Figure 1, 12) are used in patients with organ transplants. The agent specifically binds to cyclophilin expressed in T lymphocytes. The production of IL-2, IL-3, IL-4, granulocyte colony-stimulating factor (G-CSF) and TNF-α is reduced by cyclosporine in T-lymphocytes [125,126]. Sirolimus (Figure 1, 13) (Rapamycin) a macrocyclic lactone immunosuppressive drug was also derived from the fungus Streptomyces hygroscopicus [127]. It binds to FK-bound protein 12 and serine threonine kinase, mTOR, inhibiting the transduction of IL-2R and other cytokine signals relevant to allograft rejection [128,129].
Table 4.
Marine algal flora and their therapeutic chemical constituents.
| Algal Species | Chemical | Immunomodulatory Activity | Ref(s) |
|---|---|---|---|
| Eisenia arborea | Phlorotannin | Inhibits IgE and exhibits anti-degranulation effects; changes Th1/Th2 balance in Brown Norway rat strain. | [118] |
| Endarachne binghamiae | Polysaccharides (Sodium alginate, alginic) | Stimulates concentration-dependent proliferation of T cells and significant induction of the production of TNF-α and nitric oxide in macrophages and IFN-γ in T cells. | [130] |
| Caulerpa cupressoides, Pterocladiella capillacea and Solieria demonstrate | Lectins | Improves the IL-10 induction and induces the immune response of Th2 in mouse splenocytes. | [120] |
| Gracilaria verrucosa | Enone fatty acids | Inhibits the production of NO, TNF-α, and IL-6 inflammatory biomarkers. | [119] |
| Sargassum ilicifolium | Terpenes, steroids and lipids | Demonstrate chemotactic, phagocytic and intracellular killing of human neutrophils, and show a significant immunostimulatory effect in vivo. | [90,131] |
| Laminaria japonica | Laminarin oligosaccharides & polysaccharides | Apoptotic cell death protein was significantly reduced by laminarin oligosaccharides. | [132] |
| Nannochloropsis oceanica | Ethanol extract | Inhibits NO generation and downregulates NF-κB and β-secretase activities in BV-2 cells. | [133] |
| Monostroma nitidum | Sulfated polysaccharides | RAW 264.7 cells were stimulated by polysaccharides, which produced considerable NO, and PGE2 induces strong immunomodulation. | [134] |
| Hijikia fusiforme | Polysaccharides | Enhanced activity for the proliferative response of spleen cells in endotoxin nonrespondent C3H / HeJ mice. | [135] |
| Gyrodinium impudicum | Polysaccharides | Gyrodinium impudicum show immunostimulatory effects and enhance the tumoricidal activities of macrophages and NK cells in vivo. | [136] |
| Ulva fasciata | Lipopolysaccharides | Ulva in the diet significantly increases defense factors such as haemogram, agglutination index, phagocytic rate, bacterial clearance and serum bactericidal activity. | [92] |
| Sargassum thunbergii | Fucoidan | Fucoidan enhances phagocytosis and macrophage chemiluminescence. | [137] |
| Meristotheca papulosa | Polysaccharides | Extracts of M. papulosa significantly stimulated the proliferation of human lymphocytes. | [138] |
| Focellatus | Carrageenan | λ-carrageenan showed antitumor activity and lymphocyte activation in mice transplanted tumor. | [139] |
| Chlorella stigmatophora | Polysaccharides | Chlorella stigmatophora extract shows anti-inflammatory effect in paw oedema test and immunomodulatory effects in delayed hypersensitivity test. | [140] |
| Spirulina fusiformis | Polysaccharides & β -carotene | Spirulina fusiformis suppresses adjuvant-induced arthritis in mice. | [141,142] |
| Ceratodictyon spongiosum | trans-ceratospongamide (Peptide) | Potent inhibition of sPLA2 expression in an anti-inflammatory cell model. | [143] |
| Eisenia bicyclis | Phlorotannins Dieckol, Eckol | Inhibits LPS-induced NO production, iNOS and COX-2 protein levels and t-BHP-induced ROS generation in RAW 264.7 cells. | [144,145] |
| Eckolonia cava | Fucodiphloroethol | Degranulation in RBL-2H3 cells induced by IgE. | [146,147] |
| Rhipocephalus phoenix | Rhipocephalin (Sesquiterpene) | Bee venom sPLA2 inhibitory activity. | [148] |
| Crypthecodinium cohnii | Exopolysaccharide EPCP1-2 | Regulates the expression of TLR-4, MAPK and NF-κB signaling pathways | [149] |
| Gyrodinium impudium | Sulphated polysaccharide P-KG103. | Activates NO production in a JNK-dependent manner and stimulates cytokines IL-1, IL-6, and TNF-α production in macrophages. | [136,150] |
| Ishige okamurae | Diphlorethohydroxycarmal-ol (Phlorotannin) | Inhibits the IL-6 production and expression of NF-κB in murine macrophage RAW 264.7 cells. | [151] |
| Fucus distichus | Phlorotannin subfraction | Reduces TNF-α, IL-10, MCP-1 and COX-2 expression. | [152] |
| Dinoflagellates (Protoceratium reticulatum, Lingulodinium polyedrum, Gonyaulax spinifera) | Yessotoxin (Polyketide) | Inhibits macrophage phagocytosis and TNF-α, MIP-1α & MIP-2 expression. | [153,154] |
| Laurencia claviformis, Laurencia filiformis, Laurencia tasmanica, Laurencia undulata | Pacifenol (Terpenoid) | Anti-inflammatory activity, reduces the production of leukotriene B4 (LTB4) and thromboxane B2 (TXB2). | [155,156] |
| Stypopodium flabelliforme | Epitaondiol (Terpenoid) | Anti-inflammatory effects, inhibits the release and modulation of the COX pathway eicosanoids (LTB4 and TXB2). | [157,158] |
| Lobophora variegata | Lobophorins (Macrolides) | Anti-inflammatory properties. | [159] |
| Cymopolia barbata | Bromohydroquinones cymopol and cyclocymopol | Bee venom sPLA2 inhibitory activity. | [160] |
| Stypoposium flabelliforme | Meroterpene epitaondiol | Potent anti-inflammatory agent with strong activity on TPA induced ear oedema in mice and human neutrophils. | [161] |
| Vidilia obtusaloba | Bromophenols vidalols | Bee venom sPLA2 inhibitory activity. | [162] |
COX, cyclooxygenase; IFN, interferon; IL, interleukin; JNK, c-Jun NH2-terminal kinase; IgE, immunoglobulin E; iNOS, inducible nitric oxide synthase; LKB4; leukotriene B4; LPS, lipopolysaccharide; MAPK; mitogen-activated protein kinase; MCP-1, monocyte chemoattractive protein-1; MIP, macrophage inflammatory protein; NF-kB, nuclear factor-κB; NO, nitric oxide; PGE2, prostaglandin E2; PLA2, phospholipase A; t-BHP, tert-butylhydroperoxide; TLR, toll-like receptor; TNF, tumour necrosis factor; TPA, 12-O-tetradecanoylphorbol13-acetate; TXB2, thromboxane B2.
Semivioxanthin (Figure 1, 14) from marine derived fungi are found to regulate the production of TNF-α and upregulate the expression of MHC II, CD80, and CD86 [163]. Brevicompanine E (Figure 1, 15) isolated from the oceanic fungus Penicillium sp. is potentially useful for modulating neuroinflammation by attenuating NF-κB and activator protein-1 (AP-1) activity in LPS-induced microglia [164]. Brevicompanine E (Figure 1, 15) inhibits LPS-induced IκBα degradation and NF-κB nuclear translocation, and represses phosphorylation of c-Jun NH2-terminal kinase (JNK) and Akt (serine/threonine-specific protein kinase) [164]. Azonazine (Figure 1, 16) hexacyclic dipeptide obtained from the Hawaiian marine sedimentary fungus Aspergillus insulicola exhibits anti-inflammatory activity by inhibiting the production of NF-κB [165]. Three isocoumarins, (Figure 1, 17) dichlorodiaportintone, desmethyldichlorodiaportintone, and desmethyldichlorodiaportinol, from the marine mangrove endophytic fungus, Ascomycota sp. CYSK-4, produce anti-inflammatory activity by inhibiting LPS-induced NO production in RAW 264.7 cells [166]. An anthraquinone derivative, questinol (Figure 1, 18), isolated from the fungus Eurotium amstelodami exhibits an anti-inflammatory effect by significantly inhibiting prostaglandin E2 (PGE2) and NO production in LPS-stimulated RAW 264.7 cells. The production of pro-inflammatory cytokines, including IL-1, IL-6 and TNF-α is inhibited and inducible nitric oxide synthase (iNOS) expression levels suppressed in a dose-dependent manner [167]. Some representative immunomodulatory and anti-inflammatory chemical constituents isolated from marine fungi are listed in Table 5.
Apart from their economic importance, fungi have been utilized as food, usually collected from their fruiting bodies, mushrooms. Some mushrooms can stimulate the immune system, modulate cellular and humoral immunity, and potentiate anti-tumorigenic activity, and potentially rejuvenate immune systems weakened by the chemotherapy and radiotherapy used for cancer treatment. This ability of mushrooms therefore qualifies them as candidates for immunotherapy in cancer and other diseases [168].
2.6. Mangroves and Other Higher Plants
Partially submerged in the ocean, mangroves form a tangled network of above-ground roots, which creates a unique and complex habitat for all sorts of marine life. Mangroves have long been used in fisher-folk medicine to treat disease [169,170]. Some mangroves, like Rhizophora mangle and R. mucronata, have been screened for their anti-ulcer, anti-viral, and anti-inflammatory activities [171,172,173]. Leaf extract from Rhizophora apiculata has been shown to inhibit HIV-1 or HIV-2 or SIV viruses in various cell cultures [174]. Extract of Rhizophora apiculata has shown anti-inflammatory and anti-tumor activity against B16F10 melanoma cells in BALB/c mice. Rhizophora apiculata substantially reduces acute inflammation in mice induced by carrageenan, as well as inflammatory oedema induced by formalin [175]. Extract of rhizome from Acorus calamus inhibited the growth of many human and mouse cell lines. In hPBMCs the production of IL-2, NO, and TNF-α was inhibited, IFN-γ and cell-surface markers CD16 and HLA-DR were not affected, but CD25 was downregulated [176]. Some representative immunomodulatory and anti-inflammatory chemical constituents isolated from marine mangroves are listed in Table 6.
2.7. Marine Animals and Others
Entire marine animals, and their parts, contribute to the triggering of several biomedical mechanisms involved in inflammatory/allergic cascades [177]. An extract from the Caribbean Gorgonian Pseudopterogorgia elisabethae shows anti-inflammatory activity due to the presence of unusual diterpene glycosides, and is now used in cosmetic skin products as an anti-allergic factor [178]. Stichodactyla toxin (ShK)-186, a peptide toxin from sea anemones, blocks Kv1.3 potassium channels with a high degree of specificity. Kv1.3 potassium channels play a critical role in regulating the function of effector-memory T cells and class-switched memory B cells that are implicated in ADs [179]. Whole-body extracts of the marine prawn Nematopaleamon tenuipes (PEP), two gastropods, Euchelus asper (EAE) and Hemifusus pulgilinus (HPE), produced immunosuppression on Swiss albino mice in a concentration dependent manner [180]. An α-d-Glucan called MP-A, isolated from Mytilus coruscus (hard-shelled mussel), has shown anti-inflammatory activity in THP-1 human macrophage cells. MP-A suppresses LPS-induced TNF-α, NO, and PEG2 production via the TLR4 pathway [181]. Fatty acid extract from the tunicate Halocynthia aurantium significantly and dose-dependently increases NO and PGE2 production in RAW264.7 cells, producing immune enhancement without cytotoxicity. These fatty acids also regulate the transcription of immune-associated genes, including iNOS, IL-1β, IL-6, COX-2, and TNF-α [182]. Some representative immunomodulatory and anti-inflammatory chemical constituents isolated from marine creatures are listed in Table 6.
Table 5.
Marine fungi and their therapeutic chemical constituents.
| Marine Fungi | Chemical(s) | Immunomodulatory Activity | Ref(s) |
|---|---|---|---|
| Neocosmospora vasinfecta | cyclosporine | Calcineurine complex inhibition with cyclophilines. | [183] |
| Streptomyces hygroscopicus | Sirolimus macrocyclic lactone | Inhibits IL-2R signal transduction and other cytokine signals. | [127,128] |
| CTD-13C | Semivioxanthin | Regulates expression of TNF-α, CD80, CD86 and MHC II in RAW 264.7 cells. | [163] |
| Penicillium sp. | Brevicompanine E | Reduces the production of proinflammatory cytokines induced by LPS. | [164] |
| Toxicocladosporium sp. SF-5699. | Citreohybridonol | Suppresses neuroinflammatory enzymes and cytokines associated with NF-кB and MAPK in BV2 cells stimulated by LPS. | [184] |
| Aspergillus insulicola | Azonazine (Dipeptide) | Inhibits the production of NF-κB luciferase and nitrite. | [165] |
| Aspergillus sp. SF-5921 | Aurantiamide acetate | Exhibits NF-κB, JNK, and p38 inhibition in BV2 microglia cells. | [185] |
| Ascomycota sp. CYSK-4 | Isocoumarins | Inhibits the production of NO in LPS-induced RAW 264.7 cells | [166] |
| Xylaria sp. 2508 | Xyloketal | Exhibits neuroprotective effect on neonatal hypoxic-ischemic brain injury both in vivo and in vitro. | [186] |
| Eurotium amstelodami | Questinol (Anthraquinone) | Inhibits NO and PGE2 production in LPS-stimulated RAW 264.7 cells. | [167] |
| Eurotium sp. SF-5989 | Neoechinulins A and B (Diketopiperazine) | PGE2 and NO generation as well as iNOS and COX2 expression are downregulated. Diminishes IL-1 and TNF-α secretion. | [187] |
| Chaetomium globosum | Chaetoglobosin Fex | Suppresses LPS-stimulated IL-6, monocyte chemotactic protein-1, and TNF-α in peritoneal macrophages and mouse macrophage cells. | [188] |
| Penicillium paxilli Ma(G)K | Pyrenocine A | Inhibits gene expression in LPS-stimulated macrophages due to NF-κB-mediated signal transduction. | [189] |
| Ecklonia stolonifera | Phlorofucofuroeckol (Phlorotannin) | Inhibits NO and PGE2 production by the suppressing iNOS and COX-2 protein expression. | [190] |
CD, cluster of differentiation; COX, cyclooxygenase; IL, interleukin; JNK, c-Jun NH2-terminal kinase; iNOS, inducible nitric oxide synthase; LKB4; leukotriene B4; MAPK; mitogen-activated protein kinase; MHC, major histocompatibility complex; NF-kB, nuclear factor-κB; NO, nitric oxide; PGE2, prostaglandin E2; PLA2, phospholipase A2; TLR, toll-like receptor; TNF, tumor necrosis factor.
Table 6.
Mangroves, corals and other marine creatures and their therapeutic chemical constituents.
| Species | Chemical(s) | Immunomodulatory Activity | Ref(s) |
|---|---|---|---|
| Ecteinascidia turbinate | Yondelis (Trabectedin) | Reduces the proliferation of monocytes and the differentiation of ex vivo macrophages. | [191] |
| Rhizophora apiculata | Leaf extract | Inhibits HIV-1 or HIV-2 and reduces acute inflammation. | [174,175] |
| Acorus calamus | Rhizome extract | Inhibits cell proliferation and IL-2, NO, and TNF-α production is encouraged. | [176] |
| Pseudopterogorgia elisabethae | Diterpene glycosides | Inhibits TPA induced oedema in mouse, MPO release in human PMNs and, NO production in J774 macrophages. | [178] |
| Stichodactyla helianthus | Peptide ShK | Regulates the function of effector-memory T cells and class-switched memory B cells. | [179] |
| Mytilus coruscus | D-Glucan | Suppresses the production of LPS-induced TNF-α, NO, and PEG2. | |
| Halocynthia aurantium | Fatty acid | Increases production of NO and PGE2 in RAW 264.7 cells. | [182] |
| Lepeophtheirus salmonis | Trypsins | Causes an inhibitory effect on central inflammatory gene (IL-1β) | [192] |
| Litopenaeus vannamei | Polysaccharides | Exhibits immunomodulatory action of superoxide dismutase and its possible use as an indicator of immune responses. | [193] |
| Nematopaleamon tenuipes Hemifusus pugilinus Euchelus asper & Rastrelliger kanagurta | Fractions of Petroleum ether:ethyl acetate (1:1) | Exhibits immunosuppressive activity in the plaque forming cell assay. | [180,194] |
| Crenomytilus grayanus | Mytilan (Bioglycan) | Mytilan isolated from the mussel mantle Crenomytilus grayanus is highly immunomodulating. | [195] |
| Bryozoans | Convolutamydine A (Oxindole alkaloid) | Inhibits COX-2, iNOS, IL-6, PGE2 and TNF-α production. | [196] |
| Seleronephthya gracillimum | Pregnane-type steroids (Sclerosteroid) | Inhibits the expression of both iNOS and COX-2 proteins in LPS induced macrophages. | [197] |
| Marthasterias glacialis | Ergosta-7,22-dien-3-ol | Anti-inflammatory. Effective against iNOS, CHOP and IκB-α expression. | [198] |
| Astropecten polyacanthus | Steroids | Inhibits pro-inflammatory cytokine secretion, including IL-12, p40, IL-6 and TNF-α. | [199] |
| Lobophytum micchaelae | Michosterols (Polyoxygenated steroids) | Suppresses the generation of superoxide anion and elastase release in human neutrophils stimulated by N-formyl-methionyl-leucyl-phenylalanine /cytochaslasine B. | [200] |
| Paralemnalia thyrsoides | Isoparalemmone (Sesquiterpenoid) | Inhibits iNOS protein expression in activated RAW 264.7 cells. | [201] |
| Cladiella hirsuta | Hirsutalins (Diterpenes) | Inhibits LPS-stimulated iNOS protein production. | [202] |
| Lobophytum leavigatum | Laevigatol | Inhibitory effects on NF-κB-induced transcriptional activity in Hep-G2 cells. | [203] |
| Sinularia gibberosa | Gibberoketosterol (Steroids) | Inhibits the production of iNOS and COX-2 proteins in LPS-stimulated RWA 264.7 cells. | [204] |
| Pseudopterogorgia elisabethae | Pseudopterosins (Diterpene glycosides) | Blocks zymosan-induced eicosanoid release in RAW 264.7 cells. | [178] |
| Eunicea fusca | Fucosides (Diterpene arabinose glycosides) | Inhibits inflammation in the oedema model induced by 12-O- tetradecanoylphorbol-13-acetate. | [205,206] |
| Hexaplex trunculus, Charonia tritonis | Flesh and ashes of burned shell | Strengthens body’s immune system; sore and wound healing property. | [207] |
| Potamididae | Shell and flesh | Inhibits the inflammation of the mouth, recurrent aphthous ulcer, and gingivitis. | [208] |
| Eudistoma toealensis | Staurosporine & Enzastaurin | Ameliorates neuroinflammation by reducing demyelination and axonal damage. | [31] |
| Haliotis discus hannai | Extracts fermented with C. militaris mycelia (HFCM-5) | Inhibits the production of NO in RAW 264.7 cells. | [209] |
| Capnella imbricate | Capnellene | Inhibits iNOS and COX-2 in IFN-γ-stimulated microglial cells. | [210] |
| Haliotis diversicolor | Shell powder | Decreases iNOS expression and enhances the function of macrophages. | [211] |
| Filopaludina bengalensis | Footpad lipid extract | Inhibits ROS, TNF-α, and NO production. | [212] |
| Dicathais orbita Gmelin | Chloroform extract of the hypobranchial gland | Inhibits the production of NO, downregulated the production of TNFα in RAW 264.7. | [213] |
| Perna canaliculus Gmelin | Novel omega 3 polyunsaturated fatty acids | Inhibits the biosynthesis of cholesterol, COX-2, TNF-α and PGE. Inhibits TNF-α and IL-12p40 production in THP-I. | [214] |
| Anadara kagoshimensis | Polypeptide fraction | Inhibits NO in LPS-stimulated macrophage RAW 264.7cells. Inhibit IL-6, TNF-α, and IL-8 in human cervical cancer HeLa cells. | [215] |
| Fissurella Latimarginata Sowerby | Hemocyanin | Increases IFN-γ and higher numbers of tumor-infiltrating CD4+ lymphocytes. The generation of IL-6, IL-12, IL-23 and TNF-α in dendritic cells increases rapidly. | [216] |
| Perna canaliculu Mytilus unguiculatus s, | Lipid extract | Reduces the swelling of paw oedema. Inflammatory mediators (LTB4, PGE 2, and TXB2) and pro-inflammatory cytokines (IL-1, IL-6, INF-γ, and TNF-α) have been suppressed. | [217] |
| Sepiella inermis | Zhikang Capsule | Suppresses TNF-α, IFN-γ, IL-1β, and IL-12. Anti-inflammatory mediators (IL-4 and IL-10) have been promoted. | [218] |
| Oily fishes | Marine n-3 polyunsaturated fatty acids | Decreases human T cell spread, slows onset of arthritis, reduces paw swelling, reduces knee joint pathology, modulates a range of immunological reactions associated with RA. | [219] |
| Sinularia kavarattiensis | Sinuleptolide | IL-1β, IL-6, IL-8, IL-18, and TNF-α inhibition. | [220] |
| Carijoa sp. | Steroid glycoside carijoside | Neutrophil superoxide and elastase inhibition. | [221] |
| Sinularia gyrosa | Terpene gyrosanolides B & C | Inhibits iNOS expression in macrophages. | [222] |
| Sinularia flexibilis | 11-Dehydrosinulariolide | Attenuates 6-OHDA-induced downregulation of TH-immunoreactivity and 6-OHDA-induced upregulation of DJ-1 protein in rat and zebrafish models. | [223] |
| Klyxum simplex | Klysimplexin sulfoxide (Terpene) & simplexin E | Inhibits expression of COX-2 and iNOS in macrophages. | [224,225] |
| Lobophytum crassum | Diterpenes | Inhibits NO release and iNOS expression in macrophages. | [226] |
| Nephthea chabroli | Nebrosteroid I (Steroid) | Inhibits iNOS expression in macrophages. | [227] |
| Hyriopsis cumingii lea | Polysaccharide | Activates adaptive immune response including T and B cells. | [227] |
| Styela plicata | Dermatan sulfate (Polysaccharide) | Lymphocyte and macrophage, as well as TNF-α, TGF-β and VEGF, have significantly decreased in inflamed colon of the rats. | [228] |
| Lobophytum durum | Durumhemiketalolide (Terpene) | Inhibits expression of macrophage COX-2 and iNOS. | [229] |
| Lemnalia cervicorni | Lemnalol | Inhibits spinal TNF-α in microglial cells and astrocytes in neurophathic rats. | [230] |
| Sarcophyton ehrenbergi | Glycolipid & sarcoehrenosides | Inhibits iNOS expression in macrophages. | [229] |
| Sarcophyton crassocaule | Sarcocrassocolides A & B (Terpene) | Inhibits iNOS expression in macrophages. | [231] |
| Aplidium species | Rossinones A & B (Terpene) | Inhibits neutrophil superoxide. | [232] |
| Nephthea erecta & Nephthea chabroli | Chabrosterol (Steroid) | Inhibits iNOS and COX-2 expression in macrophages. | [233] |
| Mastigias papua | Symbiopolyol (Polyketide sulfate) | Inhibited expression of inducible vascular cell adhesion molecule-1, which binds to leukocytes in early inflammation stages. | [234] |
| Shellfish & finfish sp. | Docosahexaenoic acid | Inhibits carrageenan-induced microglial activation, p38 MAPK phosphorylation, and TNF- α and IL-1β mRNA expression in spinal cord. | [235] |
CHOP, C/EBP homologous protein; COX, cyclooxygenase; HIV, human immunodeficiency virus; IgE, immunoglobulin E; IL, interleukin; iNOS, inducible nitric oxide synthase; LKB4; leukotriene B4; MAPK; mitogen-activated protein kinase; MHC, major histocompatibility complex; NF-kB, nuclear factor-κB; NO, nitric oxide; PGE2, prostaglandin E2; PMNs, polymorphonuclear neutrophils; PLA2, phospholipase A; TLR, toll-like receptor; TNF, tumor necrosis factor; TPA, 12-O-tetradecanoylphorbol13-acetate; TXB2, thromboxane B2.
3. Anti-inflammatory and Immunomodulatory Effects of the Chemical Constituents of Marine Flora
Marine organisms are not only adapted to life in water with high salt concentrations, but have incorporated halogens into their chemical constituents, since ocean water contains chloride, bromide and iodide [194]. The extensive utilization of halogen ions by various marine organisms has important consequences for their overall composition. A plethora of chemical compounds have been discovered from this source. The chemical uniqueness of marine organism-derived compounds has accelerated drug discovery from those marine sources which have the highest probability of having novel molecules and interesting biological activity [236]. Marine flora is a prolific source of bioactive constituents including polysaccharides, oligosaccharides, terpenoids, steroids, alkaloids, polyphenols and antioxidants.
3.1. Polysaccharides
The most abundant and chemically complex organic molecules in the oceans are polysaccharides (Figure 1, 19) [237]. Polysaccharides (Figure 1, 19) of marine origin are a class of biochemical compounds that has been shown to have valuable therapeutic properties. These compounds are considered to be biocompatible and to have little or no toxicity. Marine algae and bacteria possess an extensive and valuable chemical library of unique polysaccharides. Sulphated polysaccharides can enhance the innate immune response by promoting the tumoricidal activities of macrophages and natural killer cells [136,238,239,240]. Polysaccharides (Figure 1, 19) from macro- and microalgae have anti-inflammatory and immunomodulatory properties [241,242]. Among the marine algal polysaccharides, fucoidans, which are fucose-containing sulphated polysaccharides from brown seaweeds, have immunomodulatory effects [243,244]. EPCP1-2, a marine EPS extracted from Crypthecodinium cohnii, exhibits anti-inflammatory activity achieved by regulating the TLR4 pathway [245]. P-KG03 sulphated polysaccharide, derived from marine microalgae Gyrodinium impudium strain KG03, activates the production of NO in a JNK-dependent manner and stimulates the production of cytokines IL-1β and 6 and TNF-α in macrophages, and prevents the growth of tumor cells in vitro and in vivo [136,150]. Alginic acid, a colloidal polysaccharide from brown seaweed, inhibits the secretion of TNF-α and IL-1 [246]. Several bacteria found in the deep-sea, in shallow hydrothermal vents, the Antarctic, and hypersaline lakes produce EPS [247,248,249,250,251]. EPS1 from a haloalkaliphilic, thermophilic strain of Bacillus licheniformis T14 hinders HSV-2 replication in hPBMCs. The non-cytotoxic exopolymer EPS1-T14 can stimulate the immune response and thus contribute to host defense against viruses [23].
3.2. Alkaloids
Alkaloids, a structurally diverse group of secondary metabolites containing nitrogen, have a range of biological activities. Alkaloids are mostly found in higher plants, but many marine organisms also contain alkaloids [252,253]. Alkaloids from the marine sponges Axinella verrucosa and Acanthella aurantiaca have been characterized as NF-κB-specific inhibitors [100]. An oxindole alkaloid, convolutamydine A (Figure 1, 20) and its two analogs, ISA003 and ISA147 from marine bryozoans inhibits the formalin-induced licking behavior significantly in mice, migration of leucocytes, and expression of COX-2, PGE2, iNOS, IL-6, and TNF-α in RAW 264.7 cells [196]. Neoechinulins A (Figure 1, 21) and B, two diketopiperazine indole alkaloids from marine fungus Eurotium sp. SF-5989 exert in vitro anti-inflammatory activity on LPS-stimulated RAW 264.7 cells. Neoechinulin A (Figure 1, 21) was considered safe in vitro using a cell viability assay, but neoechinulin B exhibited toxicity. Neoechinulin A (Figure 1, 21) derived from Microsporum sp. downregulates the formation or expression of COX-2, PGE2, NO, ROS, iNOS, IL-1, IL-6, and TNF-α in oligomeric amyloid-β activated BV-2 microglial cells. This compound also downregulates apoptosis mediated by activated microglia in pheochromocytoma PC-12 cells and reduces the nuclear translocation of NF-κB p50 and p56 subunits. Neoechinulin A (Figure 1, 21) can also inhibit neuroinflammation in Alzheimer’s disease [254]. Cytochalasan-based alkaloid chaetoglobosin Fex (Cha Fex) (Figure 1, 22), isolated from the fungus Chaetomium globosum, suppresses IL-6, TNF-α and monocyte chemotactic protein-1 in LPS-stimulated peritoneal macrophages and RAW 264.7 cells. Cytokine mRNA expression is lowered, entry of the p65 subunit of NF-κB into the nucleus and LPS-elicited breakdown of IκBα is impaired, and the levels of the extracellular-signal-related kinase (ERK1/2), p38, and c-Jun is reduced by Cha Fex (Figure 1, 22) alkaloid. In addition, the upregulation of membrane-associated CD-14 expression induced by LPS on RAW 264.7 cells and human monocytes was suppressed [188]. Alkaloids have been used by humans for a variety of purposes for more than 4000 years. Alkaloids and alkaloid-containing taxa will undoubtedly continue to play an important role in modern drug development [255].
3.3. Polyphenols
More than 8000 polyphenolic compounds are found in marine flora, including phlorotannins, flavonoids, anthocyanins, tannins, lignin, epigallocatechin, epicatechin, catechin, and hydroxylated polybrominated diphenyl ethers [256,257,258]. Polyphenols with multiple phenolic structural units are bioactive and are widely distributed in plants [257] and have a wide range of biological activities including antioxidant [259], cardiovascular protective, anti-cancer [260,261], anti-inflammatory and immune-modulatory effects [262,263]. Modern molecular and cellular biology techniques have led to a greater understanding of the benefits arising from polyphenols [264,265,266,267]. Cellular signaling and regulation of gene expression by polyphenols through modulation of NF-κB has a significant impact on cancer and chronic inflammation. For instance, resveratrol acts on the NF-κB pathway at multiple levels and is able to down-regulate its expression, phosphorylation and transcription activity [268,269,270,271]. Diphlorethohydroxycarmalol (DPHC) (Figure 1, 23), a phlorotannin from Ishige okamuarae, exerts an anti-inflammatory effect by strongly inhibiting IL-6 production in LPS-stimulated RAW264.7 cells. In addition, DPHC inhibits the expression of signal transducer and activator of transcription 5 (STAT5) signaling and increases the production of suppressor of cytokine signaling 1 (SOCS1) [151]. A phlorotannin sub-fraction isolated from Fucus distichus reduces TNF-α, IL-10, MCP-1 and COX-2 expression. The sub-fraction also lowers downstream TLR activation and expression of inflammatory biomarkers. In view of the potential cellular signaling capabilities of polyphenols, phlorotannin is beneficial because it acts as a free radical scavenger, and can also modulate inflammatory signaling receptors such as TLRs and downstream protein pathways, including NF-κB, JNK and p38 MAPKs [152].
3.4. Steroids/Sterols
Steroids are lipophilic compounds derived from cholesterol, and have a variety of marine, terrestrial, and synthetic sources. Steroids and their metabolites play an important role in the physiology and biochemistry of living organisms. For example they are used as hormone antagonists [272], contraceptives [273], cardiovascular therapeutic agents [274], anti-cancer agents [275], osteoporosis treatments [276], anesthetics, antibiotics, anti-asthmatics, and anti-inflammatories [277].
Steroidal compounds isolated from sponges that modulate the pregnane X receptors (PXRs) are effective in reducing intestinal inflammation by manipulating NF-κB activity [278,279]. Solomonsterol A (Figure 1, 24) from the marine sponge Theonella swinhoei is a selective PXR agonist and has been shown to mitigate systemic inflammation and immune system disturbances in a RA mouse model. Solomonsterol A (Figure 1, 24) inhibits the development of arthritis caused by anti-collagen antibodies in transgenic mice harboring a humanized PXR2. Solomonsterol A (Figure 1, 24) reduces the expression of the inflammatory markers TNF-α, IFN-γ and IL-17 and chemokines MIP1-α and RANTES, which reduces the inflammatory response [280]. Pregnane-type steroids derived from the soft coral Seleronephthya gracillimum show in vitro anti-inflammatory activity by reducing the accumulation of the pro-inflammatory proteins iNOS and COX-2. Sclerosteroid K (Figure 1, 25) and M (Figure 1, 27) reduce iNOS accumulation, while LPS-stimulated COX-2 accumulation in RAW 264.7 macrophages is prevented by these sclerosteriods [197]. Ergosta-7, 22-dien-3-ol (Figure 1, 28), an unsaturated sterol from the spiny sea star Marthasterias glacialis has anti-inflammatory activity on RAW 264.7 cells in vitro. The inflammatory markers NF-κB, iNOS, IL-6, and COX-2 are downregulated. Sterol ergosta-7, 22-dien-3-ol is most effective, but a potentially synergistic effect was obtained when this compound was administered with other compounds [198]. Steroids isolated from the starfish Astropecten polyacanthus, downregulate the generation of IL-6, IL-12 p40, and TNF-α in LPS-stimulated bone marrow-derived dendritic cells [199]. Polyoxygenated steroids michosterols A-C extracted from the soft coral Lobophytum micchaelae exhibit potent anti-inflammatory activity in stimulated neutrophils by suppressing the superoxide anion generation and elastase release in N-formyl-methionyl-leucyl-phenylalanine/cytochaslasine B (fMLP/CB) [200].
3.5. Miscellaneous Compounds with Anti-oxidant Activities
Antioxidants are agents which modulate the levels of highly reactive oxygen species (ROS), which cause damage by binding to biomolecules such as DNA. Antioxidants act by neutralizing ROS produced during biochemical reactions. In diseases such as Alzheimer’s, Parkinson’s, atherosclerosis, stroke, cancer, diabetes, RA, and IBD the antioxidant potential of bio-modulators has been studied and the underlying prophylactic and therapeutic aspects investigated [281,282,283,284,285,286]. Antioxidants can function as immune modulators and are used in conjunction with mainstream therapy in some diseases. Antioxidants have been used in disease prevention, where they serve as free radical scavengers. Tocopherols in humans and mice suppress PGE2 synthesis and enhance cell-mediated immunity [287,288,289]. Selenium augments the phagocytic abilities of macrophages and prevents CD8+ T lymphocyte damage [290]. Astaxanthin and fucoxanthin, marine carotenoids, appear to be biologically more effective than terrestrial carotenoids [291,292,293,294,295]. Astaxanthin decreases the production of NO, iNOS activity, and the production of PGE2 and TNF-α in RAW 264.7 cells in a dose-dependent manner [296]. Fucoxanthin prevents inflammation by inhibiting inflammatory cytokines TNF-α and IL-1, and limits the expression of COX-2 and iNOS, as well as eliminating excess ROS [297].
Vitamin C is an electron donor and acts as potent water-soluble antioxidant helping to prevent protein, lipid and DNA oxidation. Supplementation with vitamin C reduces IgE and histamine levels by increasing IFN-γ and deceasing IL-4, an observation which indicates the suppression of immune response of type Th2-type cytokines [298]. Trace elements such as iron, copper, selenium and antioxidants are found in eight species of red (Hypnea spinella, Gracilaria textorii, Gracilaria vermicullophyla), green (Caulerpa sertularioides, Codium simulans, Codium amplivesiculatum Ulva lactuca) and brown (Dictyota flabellata) microalgae [299]. Due to environmental conditions such as temperature and high levels of irradiation, microalgae and seaweeds have high levels of ROS, which seaweeds deactivate with their high intracellular amounts of antioxidant compounds such as polyphenols, phycobilins, carotenoids and vitamins [300].
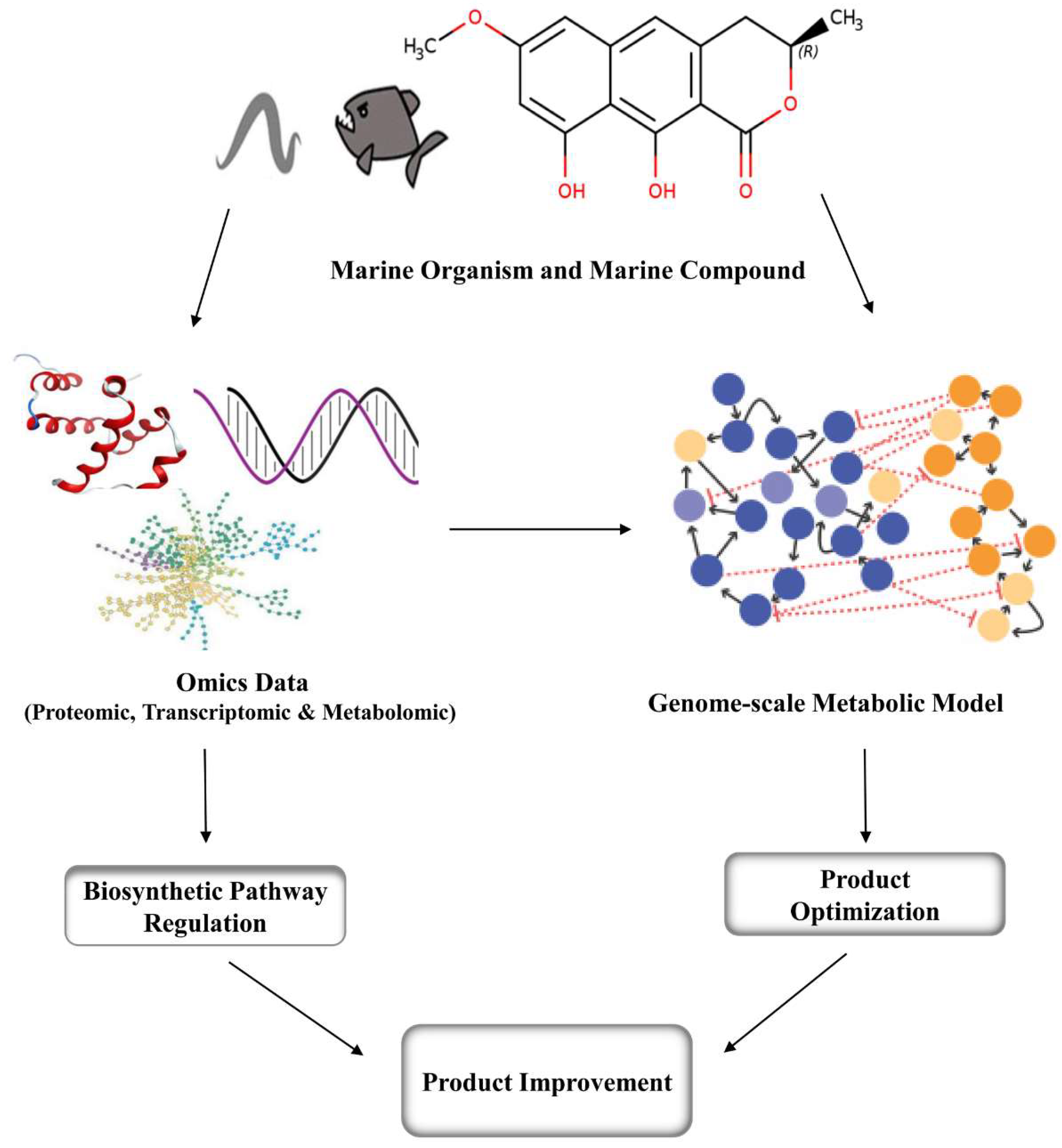
4. Metabolic Engineering and Genomic Approaches for Marine Compounds
Marine compounds that are used as drugs in microorganisms, plants and animals are synthesized in small amounts and are difficult to obtain in large quantities. This is where metabolic engineering comes into play. Using metabolic engineering, we can model organisms using existing metabolic reconstructions to discover gene knockouts which could improve the yield of products of interest. Genome-scale metabolic constraint-based flux models have been constructed for Streptomyces coelicolor A3 strains with the aim of improving and optimizing the production of antibiotics [301]. The deletion of the gonCP gene in marine actinobacterium Streptomyces coniferous resulted in improved antitumor activity of two derivatives, PM100117 and PM100118 [302]. By combining metabolic engineering and mutagenesis, it was possible to produce a level of astaxanthin in Xanthophyllomyces dendrorhous that was 89 times higher than that of the wild-type strain [303]. Advances in the understanding of microbial metabolic pathways, together with the use of new combinatorial techniques and random mutagenesis, can increase product yield and enhance efficacy [304]. Genome-scale metabolic models have been developed for several non-marine organisms and several are currently used in industrial settings. Using genomic proteomic transcriptomic and other omics data across various conditions from in-vivo experiments and literature of marine organisms the genome-scale metabolic model can be developed to produce overproducing strains of a target organism (Figure 2) [305]. In many cases, the chemical structures of biomolecules can be predicted to a certain extent, based on the analysis and biosynthetic logic of the enzymes encoded in a biosynthetic gene cluster, and their similarity to known counterparts [306]. Genomic analysis provides new insights into marine biodiversity and can reveal new drug sources. Retrieval of genomic information from marine microorganisms can also be used for the discovery of new drug molecules from microorganisms that are yet to be cultivated [307]. The CRISPR-Cas9 gene editing technology has been used in large number of microorganisms for genome modulation [308]. Researchers assembled a multifunctional metabolic engineering system based on CRISPR, which makes use of an RNA-guided nuclease. This system allows metabolic engineers to modify microorganisms via gene modifications and substitutions, and provide a practical means to reduce metabolic flux through redundant metabolic pathways and direct energy towards production of the target compound [309].
5. Conclusions
Chemical substances derived from marine organisms have proven to be a very effective in the prevention and treatment of disease. The recent development of new marine-derived treatments for cancer, inflammation and infectious diseases suggest that a focus on the development of marine medicines could be very valuable. The discovery of novel chemicals with therapeutic potential from marine sources requires the exploration of unique habitats such as deep-sea environments, as well as isolation and culturing of marine microorganisms. The traditional pharmaceutical fit of marine microorganisms as model natural product drug sources makes them more attractive. The production of bulk quantities of microbe-derived drugs is still challenging and can be addressed using metabolic engineering, in order to meet the growing need for a wide range of pharmaceuticals. Marine organisms both known and as yet undiscovered, may hold answers to some of our most pressing medical problems.
Author Contributions
B.A. and S.C. conceived the idea. B.A. and M.S. collected the data and literature. B.A. and M.S. wrote the manuscript. S.C. coordinated the project and wrote the manuscript. All authors have given their approval to the final version of the manuscript.
Funding
This work was supported by the National Research Foundation of Korea (2019R1H1A2039674) and the Commercializations Promotion Agency for R&D Outcomes funded by the Ministry of Science and ICT (2018K000369).
Conflicts of Interest
The authors declare that there are no conflict of interest.
References
- Franks, A.L.; Slansky, J.E. Multiple associations between a broad spectrum of autoimmune diseases, chronic inflammatory diseases and cancer. Anticancer Res. 2012, 32, 1119–1136. [Google Scholar] [PubMed]
- Suurmond, J.; Diamond, B. Autoantibodies in systemic autoimmune diseases: Specificity and pathogenicity. J. Clin. Invest. 2015, 125, 2194–2202. [Google Scholar] [CrossRef]
- Smedby, K.E.; Askling, J.; Mariette, X.; Baecklund, E. Autoimmune and inflammatory disorders and risk of malignant lymphomas—An update. J. Intern. Med. 2008, 264, 514–527. [Google Scholar] [CrossRef] [PubMed]
- Rubtsova, K.; Rubtsov, A.V.; Thurman, J.M.; Mennona, J.M.; Kappler, J.W.; Marrack, P. B cells expressing the transcription factor T-bet drive lupus-like autoimmunity. J. Clin. Invest. 2017, 127, 1392–1404. [Google Scholar] [CrossRef]
- Walsh, S.J.; Rau, L.M. Autoimmune diseases: A leading cause of death among young and middle-aged women in the United States. Am. J. Public Health 2000, 90, 1463–1466. [Google Scholar] [PubMed]
- Lerner, A.; Jeremias, P.; Matthias, T. The world incidence and prevalence of autoimmune diseases is increasing. Int. J. Celiac Dis. 2015, 3, 151–155. [Google Scholar] [CrossRef]
- Lohi, S.; Mustalahti, K.; Kaukinen, K.; Laurila, K.; Collin, P.; Rissanen, H.; Lohi, O.; Bravi, E.; Gasparin, M.; Reunanen, A.; et al. Increasing prevalence of coeliac disease over time. Aliment. Pharmacol. Ther. 2007, 26, 1217–1225. [Google Scholar] [CrossRef]
- Li, J.W.; Vederas, J.C. Drug discovery and natural products: End of era or an endless frontier. Biomed Khim 2011, 57, 148–160. [Google Scholar] [CrossRef]
- Munro, M.H.; Blunt, J.W.; Dumdei, E.J.; Hickford, S.J.; Lill, R.E.; Li, S.; Battershill, C.N.; Duckworth, A.R. The discovery and development of marine compounds with pharmaceutical potential. J. Biotechnol. 1999, 70, 15–25. [Google Scholar] [CrossRef]
- Boeuf, G. Marine biodiversity characteristics. C. R. Biol. 2011, 334, 435–440. [Google Scholar] [CrossRef]
- Bowler, C.; Karl, D.M.; Colwell, R.R. Microbial oceanography in a sea of opportunity. Nature 2009, 459, 180–184. [Google Scholar] [CrossRef]
- Cragg, G.M.; Newman, D.J. Natural products: A continuing source of novel drug leads. Biochim. Biophys. Acta 2013, 1830, 3670–3695. [Google Scholar] [CrossRef] [PubMed]
- Cragg, G.M.; Newman, D.J.; Weiss, R.B. Coral reefs, forests, and thermal vents: The worldwide exploration of nature for novel antitumor agents. Semin. Oncol. 1997, 24, 156–163. [Google Scholar] [PubMed]
- Antonelli, G. Underwater Robots, 4th ed.; Springer International Publishing: Berlin, Germany, 2018; p. 350. [Google Scholar]
- Elvander, J.; Hawkes, G. ROVs and AUVs in support of marine renewable technologies. In Proceedings of the 2012 Oceans, Hampton Roads, VA, USA, 14–19 October 2012; p. 1. [Google Scholar]
- Ridolfi, A.; Costanzi, R.; Fanelli, F.; Monni, N.; Allotta, B.; Bianchi, S.; Conti, R.; Gelli, J.; Gori, L.; Pugi, L.; et al. FeelHippo: A low-cost autonomous underwater vehicle for subsea monitoring and inspection. In Proceedings of the 2016 IEEE 16th International Conference on Environment and Electrical Engineering (EEEIC), Florence, Italy, 7–10 June 2016. [Google Scholar]
- Mayer, A.M.; Gustafson, K.R. Marine pharmacology in 2005-2006: Antitumour and cytotoxic compounds. Eur. J. Cancer 2008, 44, 2357–2387. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M.; Rodriguez, A.D.; Berlinck, R.G.; Hamann, M.T. Marine pharmacology in 2005-6: Marine compounds with anthelmintic, antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiprotozoal, antituberculosis, and antiviral activities; affecting the cardiovascular, immune and nervous systems, and other miscellaneous mechanisms of action. Biochim. Biophys. Acta 2009, 1790, 283–308. [Google Scholar] [PubMed]
- Gulder, T.A.; Moore, B.S. Chasing the treasures of the sea—Bacterial marine natural products. Curr. Opin. Microbiol. 2009, 12, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Waters, A.L.; Hill, R.T.; Place, A.R.; Hamann, M.T. The expanding role of marine microbes in pharmaceutical development. Curr. Opin. Biotechnol. 2010, 21, 780–786. [Google Scholar] [CrossRef] [PubMed]
- Devine, D.A.; Marsh, P.D. Prospects for the development of probiotics and prebiotics for oral applications. J. Oral Microbiol. 2009, 1, 1949. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.; Ku, S.K.; Min, G.; Choi, H.; Park, D.H.; Bae, J.S. Suppressive effects of three diketopiperazines from marine-derived bacteria on polyphosphate-mediated septic responses. Chem. Biol. Interact. 2016, 257, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Gugliandolo, C.; Spano, A.; Lentini, V.; Arena, A.; Maugeri, T.L. Antiviral and immunomodulatory effects of a novel bacterial exopolysaccharide of shallow marine vent origin. J. Appl. Microbiol. 2014, 116, 1028–1034. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.H.; Yang, Y.L.; Chen, Y.P.; Hua, K.F.; Lu, C.P.; Sheu, F.; Lin, G.H.; Tsay, S.S.; Liang, S.M.; Wu, S.H. A novel exopolysaccharide from the biofilm of Thermus aquaticus YT-1 induces the immune response through Toll-like receptor 2. J. Biol. Chem. 2011, 286, 17736–17745. [Google Scholar] [CrossRef]
- Patil, C.D.; Patil, S.V.; Salunke, B.K.; Salunkhe, R.B. Prodigiosin produced by Serratia marcescens NMCC46 as a mosquito larvicidal agent against Aedes aegypti and Anopheles stephensi. Parasitol. Res. 2011, 109, 1179–1187. [Google Scholar] [CrossRef]
- Sundaramoorthy, N.; Yogesh, P.; Dhandapani, R. Production of prodigiosin from Serratia marcescens isolated from soil. Indian J. Sci. Technol. 2009, 2, 32–34. [Google Scholar]
- Kim, H.S.; Hayashi, M.; Shibata, Y.; Wataya, Y.; Mitamura, T.; Horii, T.; Kawauchi, K.; Hirata, H.; Tsuboi, S.; Moriyama, Y. Cycloprodigiosin hydrochloride obtained from Pseudoalteromonas denitrificans is a potent antimalarial agent. Biol. Pharm. Bull. 1999, 22, 532–534. [Google Scholar] [CrossRef]
- Kawauchi, K.; Okamoto, S.; Oka, S.-I.; Kamata, H.; Yagisawa, H.; Hirata, H. Cycloprodigiosin hydrocloride suppresses tumor necrosis factor (TNF) α-induced transcriptional activation by NF-κB. Febs Lett. 2001, 507, 74–80. [Google Scholar]
- Terracciano, S.; Aquino, M.; Rodriquez, M.; Monti, M.C.; Casapullo, A.; Riccio, R.; Gomez-Paloma, L. Chemistry and biology of anti-inflammatory marine natural products: Molecules interfering with cyclooxygenase, NF-kappaB and other unidentified targets. Curr. Med. Chem. 2006, 13, 1947–1969. [Google Scholar] [CrossRef]
- Mitsiades, C.S.; Ocio, E.M.; Pandiella, A.; Maiso, P.; Gajate, C.; Garayoa, M.; Vilanova, D.; Carlos Montero, J.; Mitsiades, N.; McMullan, C.; et al. Aplidin, a marine organism-derived compound with potent antimyeloma activity in vitro and in vivo. Cancer Res. 2008, 68, 5216–5225. [Google Scholar] [CrossRef]
- Ning, C.; Wang, H.-M.D.; Gao, R.; Chang, Y.-C.; Hu, F.; Meng, X.; Huang, S.-Y. Marine-derived protein kinase inhibitors for neuroinflammatory diseases. Biomed. Eng. Online 2018, 17, 46. [Google Scholar] [CrossRef]
- Kalechman, Y.; Albeck, M.; Sredni, B. In vivo synergistic effect of the immunomodulator AS101 and the PKC inducer bryostatin. Cell. Immunol. 1992, 143, 143–153. [Google Scholar] [CrossRef]
- Philip, P.A.; Rea, D.; Thavasu, P.; Carmichael, J.; Stuart, N.S.; Rockett, H.; Talbot, D.C.; Ganesan, T.; Pettit, G.R.; Balkwill, F.; et al. Phase I study of bryostatin 1: Assessment of interleukin 6 and tumor necrosis factor alpha induction in vivo. The Cancer Research Campaign Phase I Committee. J. Natl. Cancer Inst. 1993, 85, 1812–1818. [Google Scholar] [CrossRef]
- Vaeck, M.; Grooten, J.; Hamers, R.; De Baetselier, P. The immunomodulatory effect of anti-Micrococcus luteus antibodies. I. Effect on in vitro rabbit T cell functions. Eur. J. Immunol. 1983, 13, 772–778. [Google Scholar] [CrossRef]
- Kijjoa, A.; Sawangwong, P. Drugs and Cosmetics from the Sea. Mar. Drugs 2004, 2, 73–82. [Google Scholar] [CrossRef]
- Ankisetty, S.; Khan, S.I.; Avula, B.; Gochfeld, D.; Khan, I.A.; Slattery, M. Chlorinated didemnins from the tunicate Trididemnum solidum. Mar. Drugs 2013, 11, 4478–4486. [Google Scholar] [CrossRef]
- Fürstner, A. Chemistry and Biology of Roseophilin and the Prodigiosin Alkaloids: A Survey of the Last 2500 Years. Angew. Chem. Int. Ed. 2003, 42, 3582–3603. [Google Scholar] [CrossRef]
- Asolkar, R.N.; Freel, K.C.; Jensen, P.R.; Fenical, W.; Kondratyuk, T.P.; Park, E.J.; Pezzuto, J.M. Arenamides A-C, cytotoxic NFkappaB inhibitors from the marine actinomycete Salinispora arenicola. J. Nat. Prod. 2009, 72, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Lin, X.; Wu, N.; Liu, M.; Zheng, Y.; Sheng, J.; Ji, X.; Sun, M. Targeting cellular apoptotic pathway with peptides from marine organisms. Biochim. Biophys. Acta 2013, 1836, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Bae, M.-A.; Yamada, K.; Ijuin, Y.; Tsuji, T.; Yazawa, K.; Uemura, D. Aburatubolactam A, a novel inhibitor of superoxide anion generation from a marine microorganism. Heterocycl. Commun. 1996, 2, 315. [Google Scholar] [CrossRef]
- Renner, M.K.; Shen, Y.-C.; Cheng, X.-C.; Jensen, P.R.; Frankmoelle, W.; Kauffman, C.A.; Fenical, W.; Lobkovsky, E.; Clardy, J. Cyclomarins A−C, New Antiinflammatory Cyclic Peptides Produced by a Marine Bacterium (Streptomyces sp.). J. Am. Chem. Soc. 1999, 121, 11273–11276. [Google Scholar] [CrossRef]
- Trischman, J.A.; Tapiolas, D.M.; Jensen, P.R.; Dwight, R.; Fenical, W.; McKee, T.C.; Ireland, C.M.; Stout, T.J.; Clardy, J. Salinamides A and B: Anti-inflammatory depsipeptides from a marine streptomycete. J. Am. Chem. Soc. 1994, 116, 757–758. [Google Scholar] [CrossRef]
- Strangman, W.K.; Kwon, H.C.; Broide, D.; Jensen, P.R.; Fenical, W. Potent inhibitors of pro-inflammatory cytokine production produced by a marine-derived bacterium. J. Med. Chem. 2009, 52, 2317–2327. [Google Scholar] [CrossRef]
- Singh, R.K.; Tiwari, S.P.; Rai, A.K.; Mohapatra, T.M. Cyanobacteria: An emerging source for drug discovery. J. Antibiot. (Tokyo) 2011, 64, 401–412. [Google Scholar] [CrossRef]
- Durai, P.; Batool, M.; Choi, S. Structure and effects of cyanobacterial lipopolysaccharides. Mar. Drugs 2015, 13, 4217–4230. [Google Scholar] [CrossRef] [PubMed]
- Thorgersen, E.B.; Macagno, A.; Rossetti, C.; Mollnes, T.E. Cyanobacterial LPS antagonist (CyP)-a novel and efficient inhibitor of Escherichia coli LPS-induced cytokine response in the pig. Mol. Immunol. 2008, 45, 3553–3557. [Google Scholar] [CrossRef] [PubMed]
- Macagno, A.; Molteni, M.; Rinaldi, A.; Bertoni, F.; Lanzavecchia, A.; Rossetti, C.; Sallusto, F. A cyanobacterial LPS antagonist prevents endotoxin shock and blocks sustained TLR4 stimulation required for cytokine expression. J. Exp. Med. 2006, 203, 1481–1492. [Google Scholar] [CrossRef] [PubMed]
- Molteni, M.; Bosi, A.; Rossetti, C. The effect of cyanobacterial LPS antagonist (CyP) on cytokines and micro-RNA expression Induced by Porphyromonas gingivalis LPS. Toxins (Basel) 2018, 10, 290. [Google Scholar] [CrossRef] [PubMed]
- Villa, F.A.; Lieske, K.; Gerwick, L. Selective MyD88-dependent pathway inhibition by the cyanobacterial natural product malyngamide F acetate. Eur. J. Pharmacol. 2010, 629, 140–146. [Google Scholar] [CrossRef]
- Appel, K.; Munoz, E.; Navarrete, C.; Cruz-Teno, C.; Biller, A.; Thiemann, E. Immunomodulatory and inhibitory effect of immulina((R)), and immunloges((R)) in the Ig-E mediated activation of RBL-2H3 Cells. A new role in allergic inflammatory responses. Plants (Basel) 2018, 7, 13. [Google Scholar] [CrossRef]
- Yang, H.N.; Lee, E.H.; Kim, H.M. Spirulina platensis inhibits anaphylactic reaction. Life Sci. 1997, 61, 1237–1244. [Google Scholar] [CrossRef]
- Kwan, J.C.; Eksioglu, E.A.; Liu, C.; Paul, V.J.; Luesch, H. Grassystatins A-C from marine cyanobacteria, potent cathepsin E inhibitors that reduce antigen presentation. J. Med. Chem. 2009, 52, 5732–5747. [Google Scholar] [CrossRef]
- Blunt, J.W.; Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2018, 35, 8–53. [Google Scholar] [CrossRef]
- Anjum, K.; Abbas, S.Q.; Shah, S.A.; Akhter, N.; Batool, S.; Hassan, S.S. Marine sponges as a drug treasure. Biomol. Ther. (Seoul) 2016, 24, 347–362. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.R.; Kavlekar, D.P.; LokaBharathi, P.A. Marine drugs from sponge-microbe association—A review. Mar. Drugs 2010, 8, 1417–1468. [Google Scholar] [CrossRef]
- Rinehart, K.L.; Gloer, J.B.; Cook, J.C.; Mizsak, S.A.; Scahill, T.A. Structures of the didemnins, antiviral and cytotoxic depsipeptides from a Caribbean tunicate. J. Am. Chem. Soc. 1981, 103, 1857–1859. [Google Scholar] [CrossRef]
- Nuijen, B.; Bouma, M.; Manada, C.; Jimeno, J.M.; Schellens, J.H.; Bult, A.; Beijnen, J.H. Pharmaceutical development of anticancer agents derived from marine sources. Anticancer Drugs 2000, 11, 793–811. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Currano, J.N.; Carroll, P.J.; Joullie, M.M. Didemnins, tamandarins and related natural products. Nat. Prod. Rep. 2012, 29, 404–424. [Google Scholar] [CrossRef] [PubMed]
- Vera, M.D.; Joullie, M.M. Natural products as probes of cell biology: 20 years of didemnin research. Med. Res. Rev. 2002, 22, 102–145. [Google Scholar] [CrossRef]
- Mayer, A.M.; Hamann, M.T. Marine pharmacology in 2000: Marine compounds with antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiplatelet, antituberculosis, and antiviral activities; affecting the cardiovascular, immune, and nervous systems and other miscellaneous mechanisms of action. Mar. Biotechnol. (N. Y.) 2004, 6, 37–52. [Google Scholar]
- Mayer, A.M.; Rodriguez, A.D.; Berlinck, R.G.; Fusetani, N. Marine pharmacology in 2007-8: Marine compounds with antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiprotozoal, antituberculosis, and antiviral activities; affecting the immune and nervous system, and other miscellaneous mechanisms of action. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2011, 153, 191–222. [Google Scholar] [PubMed]
- De Almeida Leone, P.; Redburn, J.; Hooper, J.; Quinn, R. Polyoxygenated Dysidea sterols that inhibit the binding of [I125] IL-8 to the human recombinant IL-8 receptor type A. J. Nat. Prod. 2000, 63, 694–697. [Google Scholar] [CrossRef] [PubMed]
- Romo, D.; Rzasa, R.M.; Shea, H.A.; Park, K.; Langenhan, J.M.; Sun, L.; Akhiezer, A.; Liu, J.O. Total Synthesis and Immunosuppressive Activity of (−)-Pateamine A and Related Compounds: Implementation of a β-Lactam-Based Macrocyclization. J. Am. Chem. Soc. 1998, 120, 12237–12254. [Google Scholar] [CrossRef]
- Pattenden, G.; Critcher, D.; Remuiñán, M. Total synthesis of (-)-pateamine A, a novel immunosuppressive agent from Mycale sp. Can. J. Chem. 2011, 82, 353–365. [Google Scholar] [CrossRef]
- Gunasekera, S.P.; Gunasekera, M.; Longley, R.E.; Schulte, G.K. Discodermolide: A new bioactive polyhydroxylated lactone from the marine sponge Discodermia dissoluta [Erratum to document cited in CA113(9):75187b]. J. Org. Chem. 1991, 56, 1346. [Google Scholar] [CrossRef]
- Arefolov, A.; Panek, J.S. Studies directed toward the total synthesis of discodermolide: Asymmetric synthesis of the C1-C14 fragment. Org. Lett. 2002, 4, 2397–2400. [Google Scholar] [CrossRef] [PubMed]
- Takamatsu, S.; Nagle, D.G.; Gerwick, W.H. Secondary metabolites from marine cyanobacteria and algae inhibit LFA-1/ICAM-1 mediated cell adhesion. Planta Med. 2004, 70, 127–131. [Google Scholar]
- Malloy, K.L.; Villa, F.A.; Engene, N.; Matainaho, T.; Gerwick, L.; Gerwick, W.H. Malyngamide 2, an oxidized lipopeptide with nitric oxide inhibiting activity from a Papua New Guinea marine cyanobacterium. J. Nat. Prod. 2011, 74, 95–98. [Google Scholar] [CrossRef]
- Silambarasan, G.; Ramanathan, T.; Nabeel, M.A.; Kalaichelvan, V.K.; Kathiresan, K.; Balasubramanian, T. Anti-Inflammatory Activity of the Marine Cyanobacterium Trichodesmium Erythraeum against Carrageenan-Induced Paw Oedema in Wistar Albino Rats. Eur. J. Inflam. 2011, 9, 53–56. [Google Scholar] [CrossRef]
- Costantino, V.; Fattorusso, E.; Mangoni, A.; Di Rosa, M.; Ianaro, A. Glycolipids from sponges. VII.1 simplexides, novel immunosuppressive glycolipids from the caribbean sponge Plakortis simplex. Bioorg. Med. Chem. Lett. 1999, 9, 271–276. [Google Scholar] [CrossRef]
- Loffredo, S.; Staiano, R.I.; Granata, F.; Costantino, V.; Borriello, F.; Frattini, A.; Lepore, M.T.; Mangoni, A.; Marone, G.; Triggiani, M. Simplexide induces CD1d-dependent cytokine and chemokine production from human monocytes. PLoS ONE 2014, 9, e111326. [Google Scholar] [CrossRef]
- Busserolles, J.; Paya, M.; D’Auria, M.V.; Gomez-Paloma, L.; Alcaraz, M.J. Protection against 2,4,6-trinitrobenzenesulphonic acid-induced colonic inflammation in mice by the marine products bolinaquinone and petrosaspongiolide M. Biochem. Pharmacol. 2005, 69, 1433–1440. [Google Scholar] [CrossRef]
- Takei, M.; Burgoyne, D.L.; Andersen, R.J. Effect of contignasterol on histamine release induced by anti-immunoglobulin E from rat peritoneal mast cells. J. Pharm. Sci. 1994, 83, 1234–1235. [Google Scholar] [CrossRef]
- Hong, S.; Kim, S.H.; Rhee, M.H.; Kim, A.R.; Jung, J.H.; Chun, T.; Yoo, E.S.; Cho, J.Y. In vitro anti-inflammatory and pro-aggregative effects of a lipid compound, petrocortyne A, from marine sponges. Naunyn Schmiedebergs Arch. Pharmacol. 2003, 368, 448–456. [Google Scholar] [CrossRef]
- Northcote, P.T.; Blunt, J.W.; Munro, M.H.G. Pateamine: A potent cytotoxin from the New Zealand Marine sponge, mycale sp. Tetrahedron Lett. 1991, 32, 6411–6414. [Google Scholar] [CrossRef]
- Low, W.K.; Dang, Y.; Schneider-Poetsch, T.; Shi, Z.; Choi, N.S.; Merrick, W.C.; Romo, D.; Liu, J.O. Inhibition of eukaryotic translation initiation by the marine natural product pateamine A. Mol. Cell 2005, 20, 709–722. [Google Scholar] [CrossRef]
- Takei, M.; Umeyama, A.; Shoji, N.; Hashimoto, T. Polyacetylenediols regulate the function of human monocyte-derived dendritic cells. Int. Immunopharmacol. 2010, 10, 913–921. [Google Scholar] [CrossRef]
- Greve, H.; Meis, S.; Kassack, M.U.; Kehraus, S.; Krick, A.; Wright, A.D.; Konig, G.M. New iantherans from the marine sponge Ianthella quadrangulata: Novel agonists of the P2Y(11) receptor. J. Med. Chem. 2007, 50, 5600–5607. [Google Scholar] [CrossRef]
- Jung, M.E.; Johnson, T.W. First total synthesis of xestobergsterol A and active structural analogues of the xestobergsterols. Org. Lett. 1999, 1, 1671–1674. [Google Scholar] [CrossRef]
- Keyzers, R.A.; Northcote, P.T.; Berridge, M.V. Clathriol B, a New 14β Marine Sterol from the New Zealand Sponge Clathria lissosclera. ChemInform 2003, 34, 279–282. [Google Scholar] [CrossRef]
- Schumacher, M.; Cerella, C.; Eifes, S.; Chateauvieux, S.; Morceau, F.; Jaspars, M.; Dicato, M.; Diederich, M. Heteronemin, a spongean sesterterpene, inhibits TNFα-induced NF-κB activation through proteasome inhibition and induces apoptotic cell death. Biochem. Pharmacol. 2010, 79, 610–622. [Google Scholar] [CrossRef]
- El-Shitany, N.A.; Shaala, L.A.; Abbas, A.T.; Abdel-Dayem, U.A.; Azhar, E.I.; Ali, S.S.; van Soest, R.W.; Youssef, D.T. Evaluation of the anti-Inflammatory, antioxidant and immunomodulatory effects of the organic extract of the red Sea marine sponge Xestospongia testudinaria against carrageenan induced rat paw inflammation. PLoS ONE 2015, 10, e0138917. [Google Scholar] [CrossRef]
- Kossuga, M.; Nascimento, A.; Reimão, J.; Tempone, A.; Taniwaki, N.; Veloso, K.; Ferreira, A.; Cavalcanti, B.; Pessoa, C.; Moraes, M.; et al. Antiparasitic, Antineuroinflammatory, and Cytotoxic Polyketides from the Marine Sponge Plakortis angulospiculatus Collected in Brazil. J. Nat. Prod. 2008, 71, 334–339. [Google Scholar] [CrossRef]
- Costantini, S.; Romano, G.; Rusolo, F.; Capone, F.; Guerriero, E.; Colonna, G.; Ianora, A.; Ciliberto, G.; Costantini, M. Anti-Inflammatory effects of a methanol extract from the marine sponge Geodia cydonium on the human breast cancer MCF-7 cell line. Mediators Inflamm. 2015, 2015, 204975. [Google Scholar] [CrossRef] [PubMed]
- Costantini, S.; Guerriero, E.; Teta, R.; Capone, F.; Caso, A.; Sorice, A.; Romano, G.; Ianora, A.; Ruocco, N.; Budillon, A.; et al. Evaluating the effects of an organic extract from the Mediterranean sponge Geodia cydonium on human breast cancer cell lines. Int. J. Mol. Sci. 2017, 18, 2112. [Google Scholar] [CrossRef] [PubMed]
- De Marino, S.; Festa, C.; D’Auria, M.V.; Bourguet-Kondracki, M.-L.; Petek, S.; Debitus, C.; Andrés, R.M.; Terencio, M.C.; Payá, M.; Zampella, A. Coscinolactams A and B: New nitrogen-containing sesterterpenoids from the marine sponge Coscinoderma mathewsi exerting anti-inflammatory properties. Tetrahedron 2009, 65, 2905–2909. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Lu, M.-C.; Su, J.-H.; Chu, C.-L.; Shiuan, D.; Weng, C.-F.; Sung, P.-J.; Huang, K.-J. Immunomodulatory effect of marine cembrane-type diterpenoids on dendritic cells. Mar. Drugs 2013, 11, 1336–1350. [Google Scholar] [CrossRef]
- Garcia-Pastor, P.; Randazzo, A.; Gomez-Paloma, L.; Alcaraz, M.J.; Paya, M. Effects of petrosaspongiolide M, a novel phospholipase A2 inhibitor, on acute and chronic inflammation. J. Pharmacol. Exp. Ther. 1999, 289, 166. [Google Scholar]
- Bourguet-Kondracki, M.-L.; Debitus, C.; Guyot, M. Dipuupehedione, a cytotoxic new red dimer from a new caledonian marine sponge hyrtios sp. Tetrahedron Lett. 1996, 37, 3861–3864. [Google Scholar] [CrossRef]
- Chandraraj, S.; Prakash, B.; Navanath, K. Immunomodulatory activities of ethyl acetate extracts of two marine sponges Gelliodes fibrosa and Tedania anhelans and brown algae Sargassum ilicifolium with reference to phagocytosis. Res. J. Pharm. Biol. Chem. Sci. 2010, 1, 302–307. [Google Scholar]
- Rifai, S.; Fassouane, A.; Pinho, P.M.; Kijjoa, A.; Nazareth, N.; Nascimento, M.S.J.; Herz, W. Cytotoxicity and inhibition of lymphocyte proliferation of fasciculatin, a linear furanosesterterpene isolated from Ircinia variabilis collected from the Atlantic Coast of Morocco. Mar. Drugs 2005, 3, 15–21. [Google Scholar] [CrossRef]
- Selvin, J.; Huxley, A.J.; Lipton, A.P. Immunomodulatory potential of marine secondary metabolites against bacterial diseases of shrimp. Aquaculture 2004, 230, 241–248. [Google Scholar] [CrossRef]
- Liu, R.; Liu, Y.; Zhou, Y.D.; Nagle, D.G. Molecular-targeted antitumor agents. 15. Neolamellarins from the marine sponge Dendrilla nigra inhibit hypoxia-inducible factor-1 activation and secreted vascular endothelial growth factor production in breast tumor cells. J. Nat. Prod. 2007, 70, 1741–1745. [Google Scholar] [CrossRef]
- Festa, C.; De Marino, S.; Sepe, V.; Monti, M.C.; Luciano, P.; D’Auria, M.V.; Débitus, C.; Bucci, M.; Vellecco, V.; Zampella, A. Perthamides C and D, two new potent anti-inflammatory cyclopeptides from a Solomon Lithistid sponge Theonella swinhoei. Tetrahedron 2009, 65, 10424–10429. [Google Scholar] [CrossRef]
- Roy, M.C.; Ohtani, I.I.; Ichiba, T.; Tanaka, J.; Satari, R.; Higa, T. New Cyclic Peptides from the Indonesian Sponge Theonellaswinhoei. Tetrahedron 2000, 56, 9079–9092. [Google Scholar] [CrossRef]
- Gunasekera, S.P.; Gunasekera, M.; Longley, R.E.; Schulte, G.K. Discodermolide: A new bioactive polyhydroxylated lactone from the marine sponge Discodermia dissoluta. J. Org. Chem. 1990, 55, 4912–4915. [Google Scholar] [CrossRef]
- Ciasullo, L.; Casapullo, A.; Cutignano, A.; Bifulco, G.; Debitus, C.; Hooper, J.; Gomez-Paloma, L.; Riccio, R. Renieramide, a cyclic tripeptide from the Vanuatu sponge Reniera n. sp. J. Nat. Prod. 2002, 65, 407–410. [Google Scholar] [CrossRef] [PubMed]
- Fung, S.-Y.; Sofiyev, V.; Schneiderman, J.; Hirschfeld, A.F.; Victor, R.E.; Woods, K.; Piotrowski, J.S.; Deshpande, R.; Li, S.C.; de Voogd, N.J.; et al. Unbiased screening of marine sponge extracts for anti-inflammatory agents combined with chemical genomics identifies girolline as an inhibitor of protein synthesis. ACS Chem. Biol. 2014, 9, 247–257. [Google Scholar] [CrossRef]
- Youssef, D.T.; Ibrahim, A.K.; Khalifa, S.I.; Mesbah, M.K.; Mayer, A.M.; van Soest, R.W. New anti-inflammatory sterols from the Red Sea sponges Scalarispongia aqabaensis and Callyspongia siphonella. Nat. Prod. Commun. 2010, 5, 27–31. [Google Scholar]
- Cimino, G.; De Rosa, S.; De Stefano, S.; Mazzarella, L.; Puliti, R.; Sodano, G. Isolation and X-ray crystal structure of a novel bromo-compound from two marine sponges. Tetrahedron Lett. 1982, 23, 767–768. [Google Scholar] [CrossRef]
- Costantino, V.; Fattorusso, E.; Mangoni, A.; Perinu, C.; Cirino, G.; De Gruttola, L.; Roviezzo, F. Tedanol: A potent anti-inflammatory ent-pimarane diterpene from the Caribbean Sponge Tedania ignis. Biorg. Med. Chem. 2009, 17, 7542–7547. [Google Scholar] [CrossRef]
- Castrillo, A.; de Las Heras, B.; Hortelano, S.; Rodriguez, B.; Villar, A.; Bosca, L. Inhibition of the nuclear factor kappa B (NF-kappa B) pathway by tetracyclic kaurene diterpenes in macrophages. Specific effects on NF-kappa B-inducing kinase activity and on the coordinate activation of ERK and p38 MAPK. J. Biol. Chem. 2001, 276, 15854–15860. [Google Scholar] [CrossRef]
- Randazzo, A.; Bifulco, G.; Giannini, C.; Bucci, M.; Debitus, C.; Cirino, G.; Gomez-Paloma, L. Halipeptins A and B: Two novel potent anti-inflammatory cyclic depsipeptides from the Vanuatu marine sponge Haliclona species. J. Am. Chem. Soc. 2001, 123, 10870–10876. [Google Scholar] [CrossRef]
- Faulkner, D.J. Marine natural products. Nat. Prod. Rep. 2001, 18, 1R–49R. [Google Scholar] [CrossRef]
- Leitch, A.E.; Haslett, C.; Rossi, A.G. Cyclin-dependent kinase inhibitor drugs as potential novel anti-inflammatory and pro-resolution agents. Br. J. Pharmacol. 2009, 158, 1004–1016. [Google Scholar] [CrossRef] [PubMed]
- Costantino, V.; Fattorusso, E.; Mangoni, A.; Teta, R.; Panza, E.; Ianaro, A. Terpioside B, a difucosyl GSL from the marine sponge Terpios sp. is a potent inhibitor of NO release. Bioorg. Med. Chem. 2010, 18, 5310–5315. [Google Scholar] [CrossRef] [PubMed]
- Ter Haar, E.; Kowalski, R.J.; Hamel, E.; Lin, C.M.; Longley, R.E.; Gunasekera, S.P.; Rosenkranz, H.S.; Day, B.W. Discodermolide, a cytotoxic marine agent that stabilizes microtubules more potently than taxol. Biochemistry 1996, 35, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Hung, D.T.; Chen, J.; Schreiber, S.L. (+)-Discodermolide binds to microtubules in stoichiometric ratio to tubulin dimers, blocks taxol binding and results in mitotic arrest. Chem. Biol. 1996, 3, 287–293. [Google Scholar] [CrossRef]
- Posadas, I.; Terencio, M.C.; Randazzo, A.; Gomez-Paloma, L.; Paya, M.; Alcaraz, M.J. Inhibition of the NF-kappaB signaling pathway mediates the anti-inflammatory effects of petrosaspongiolide M. Biochem. Pharmacol. 2003, 65, 887–895. [Google Scholar] [CrossRef]
- MacArtain, P.; Gill, C.I.; Brooks, M.; Campbell, R.; Rowland, I.R. Nutritional value of edible seaweeds. Nutr. Rev. 2007, 65, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Černá, M. Seaweed Proteins and Amino Acids as Nutraceuticals. In Advances in Food and Nutrition Research; Academic Press: Cambridge, MA, USA, 2011; Volume 64, pp. 297–312. [Google Scholar]
- Misurcova, L.; Skrovankova, S.; Samek, D.; Ambrozova, J.; Machu, L. Health benefits of algal polysaccharides in human nutrition. Adv. Food Nutr. Res. 2012, 66, 75–145. [Google Scholar]
- Tabarsa, M.; Rezaei, M.; Ramezanpour, Z.; Waaland, J.R. Chemical compositions of the marine algae Gracilaria salicornia (Rhodophyta) and Ulva lactuca (Chlorophyta) as a potential food source. J. Sci. Food Agric. 2012, 92, 2500–2506. [Google Scholar] [CrossRef]
- Rajapakse, N.; Kim, S.K. Nutritional and digestive health benefits of seaweed. Adv. Food Nutr. Res. 2011, 64, 17–28. [Google Scholar]
- Lee, J.C.; Hou, M.F.; Huang, H.W.; Chang, F.R.; Yeh, C.C.; Tang, J.Y.; Chang, H.W. Marine algal natural products with anti-oxidative, anti-inflammatory, and anti-cancer properties. Cancer Cell Int. 2013, 13, 55. [Google Scholar] [CrossRef] [PubMed]
- Makarenkova, I.D.; Logunov, D.; Tukhvatulin, A.I.; Semenova, I.B.; Zviagintheva, T.N.; Gorbach, V.I.; Ermakova, S.P.; Besednova, N.N. Sulfated polysaccharides of brown seaweeds-ligands of toll-like receptors. Biomed Khim 2012, 58, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Besednova, N.N.; Zaporozhets, T.S.; Somova, L.M.; Kuznetsova, T.A. Review: Prospects for the use of extracts and polysaccharides from marine algae to prevent and treat the diseases caused by Helicobacter pylori. Helicobacter 2015, 20, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, Y.; Matsuda, K.; Okamoto, T.; Kakinuma, M.; Amano, H. Anti-allergic effects of the brown alga Eisenia arborea on Brown Norway rats. Fish. Sci. 2008, 74, 180–186. [Google Scholar] [CrossRef]
- Lee, H.J.; Dang, H.T.; Kang, G.J.; Yang, E.J.; Park, S.S.; Yoon, W.J.; Jung, J.H.; Kang, H.K.; Yoo, E.S. Two enone fatty acids isolated from Gracilaria verrucosa suppress the production of inflammatory mediators by down-regulating NF-kappaB and STAT1 activity in lipopolysaccharide-stimulated RAW 264.7 cells. Arch. Pharm. Res. 2009, 32, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Monteiro Abreu, T.; Castelo Melo Silva, L.M.; Vanderlei, E.S.; de Melo, C.M.; Pereira, V.R.; Barros Benevides, N.M. Cytokine production induced by marine algae lectins in BALB/c mice splenocytes. Protein Pept. Lett. 2012, 19, 975–981. [Google Scholar] [CrossRef]
- Overy, D.P.; Bayman, P.; Kerr, R.G.; Bills, G.F. An assessment of natural product discovery from marine (sensu strictu) and marine-derived fungi. Mycology 2014, 5, 145–167. [Google Scholar] [CrossRef]
- Dreyfuss, M.; Härri, E.; Hofmann, H.; Kobel, H.; Pache, W.; Tscherter, H. Cyclosporin A and C. Eur. J. Appl. Microbiol. Biotechnol. 1976, 3, 125–133. [Google Scholar] [CrossRef]
- Dong, H.; Jiang, J.; Yan, T.; Zhao, J. Optimization of cyclosporin A production by Beauveria nivea in continuous fed-batch fermentation. Arch. Biol. Sci. (Serbia) 2011, 63, 907–914. [Google Scholar] [CrossRef]
- Bhosale, S.H.; Patil, K.B.; Parameswaran, P.S.; Naik, C.G.; Jagtap, T.G. Active pharmaceutical ingredient (api) from an estuarine fungus, Microdochium nivale (Fr.). J. Environ. Biol. 2011, 32, 653–658. [Google Scholar]
- Borel, J.F.; Feurer, C.; Magnee, C.; Stahelin, H. Effects of the new anti-lymphocytic peptide cyclosporin A in animals. Immunology 1977, 32, 1017–1025. [Google Scholar]
- Wiederrecht, G.; Lam, E.; Hung, S.; Martin, M.; Sigal, N. The Mechanism of Action of FK-506 and Cyclosporin A. Ann. N. Y. Acad. Sci. 1993, 696, 9–19. [Google Scholar] [CrossRef]
- Sehgal, S.N. Sirolimus: Its discovery, biological properties, and mechanism of action. Transplant. Proc. 2003, 35, 7S–14S. [Google Scholar] [CrossRef]
- Sehgal, S.N. Rapamune (Sirolimus, rapamycin): An overview and mechanism of action. Ther. Drug Monit. 1995, 17, 660–665. [Google Scholar] [CrossRef]
- Brown, E.J.; Albers, M.W.; Shin, T.B.; Ichikawa, K.; Keith, C.T.; Lane, W.S.; Schreiber, S.L. A mammalian protein targeted by G1-arresting rapamycin-receptor complex. Nature 1994, 369, 756–758. [Google Scholar] [CrossRef]
- Huang, R.; Lee, H.-T. Immunological properties of the marine brown alga endarachne binghamiae (Phaeophyceae). Int. J. Appl. Sci. Eng. 2005, 3, 167–173. [Google Scholar]
- Yende, S.R.; Harle, U.N.; Chaugule, B.B. Therapeutic potential and health benefits of Sargassum species. Pharmacogn Rev. 2014, 8, 1–7. [Google Scholar] [CrossRef]
- Kim, K.H.; Kim, Y.W.; Kim, H.B.; Lee, B.J.; Lee, D.S. Anti-apoptotic activity of laminarin polysaccharides and their enzymatically hydrolyzed oligosaccharides from Laminaria japonica. Biotechnol. Lett. 2006, 28, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.Y.; Hwang, C.J.; Lee, H.P.; Kim, H.S.; Han, S.B.; Hong, J.T. Inhibitory effect of ethanol extract of Nannochloropsis oceanica on lipopolysaccharide-induced neuroinflammation, oxidative stress, amyloidogenesis and memory impairment. Oncotarget 2017, 8, 45517–45530. [Google Scholar] [CrossRef] [PubMed]
- Karnjanapratum, S.; You, S. Molecular characteristics of sulfated polysaccharides from Monostroma nitidum and their in vitro anticancer and immunomodulatory activities. Int. J. Biol. Macromol. 2011, 48, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Okai, Y.; Higashi-Okai, K.; Ishizaka, S.; Ohtani, K.; Matsui-Yuasa, I.; Yamashita, U. Possible immunodulating activities in an extract of edible brown alga, Hijikia fusiforme (Hijiki). J. Sci. Food Agric. 1998, 76, 56–62. [Google Scholar] [CrossRef]
- Yim, J.H.; Son, E.; Pyo, S.; Lee, H.K. Novel sulfated polysaccharide derived from red-tide microalga Gyrodinium impudicum strain KG03 with immunostimulating activity in vivo. Mar. Biotechnol. (N. Y.) 2005, 7, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Itoh, H.; Noda, H.; Amano, H.; Zhuaug, C.; Mizuno, T.; Ito, H. Antitumor activity and immunological properties of marine algal polysaccharides, especially fucoidan, prepared from Sargassum thunbergii of Phaeophyceae. Anticancer Res. 1993, 13, 2045–2052. [Google Scholar]
- Shan, B.; Yoshida, Y.; Kuroda, E.; Yamashita, U. Brief communication immunomodulating activity of seaweed extract on human lymphocytes in vitro. Int. J. Immunopharmacol. 1999, 21, 59–70. [Google Scholar] [CrossRef]
- Zhou, G.; Sun, Y.; Xin, H.; Zhang, Y.; Li, Z.; Xu, Z. In vivo antitumor and immunomodulation activities of different molecular weight lambda-carrageenans from Chondrus ocellatus. Pharmacol. Res. 2004, 50, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Guzman, S.; Gato, A.; Lamela, M.; Freire-Garabal, M.; Calleja, J.M. Anti-inflammatory and immunomodulatory activities of polysaccharide from Chlorella stigmatophora and Phaeodactylum tricornutum. Phytother. Res. 2003, 17, 665–670. [Google Scholar] [CrossRef] [PubMed]
- Rasool, M.; Sabina, E.P. Appraisal of immunomodulatory potential of Spirulina fusiformis: An in vivo and in vitro study. J. Nat. Med. 2009, 63, 169–175. [Google Scholar] [CrossRef]
- Rasool, M.; Sabina, E.P.; Lavanya, B. Anti-inflammatory effect of Spirulina fusiformis on adjuvant-induced arthritis in mice. Biol. Pharm. Bull. 2006, 29, 2483–2487. [Google Scholar] [CrossRef]
- Tan, L.T.; Williamson, R.T.; Gerwick, W.H.; Watts, K.S.; McGough, K.; Jacobs, R. cis,cis- and trans,trans-ceratospongamide, new bioactive cyclic heptapeptides from the Indonesian red alga Ceratodictyon spongiosum and symbiotic sponge Sigmadocia symbiotica. J. Org. Chem. 2000, 65, 419–425. [Google Scholar] [CrossRef]
- Shibata, T.; Nagayama, K.; Tanaka, R.; Yamaguchi, K.; Nakamura, T. Inhibitory effects of brown algal phlorotannins on secretory phospholipase A2s, lipoxygenases and cyclooxygenases. J. Appl. Phycol. 2003, 15, 61–66. [Google Scholar] [CrossRef]
- Jung, H.A.; Jin, S.E.; Ahn, B.R.; Lee, C.M.; Choi, J.S. Anti-inflammatory activity of edible brown alga Eisenia bicyclis and its constituents fucosterol and phlorotannins in LPS-stimulated RAW264.7 macrophages. Food Chem. Toxicol. 2013, 59, 199–206. [Google Scholar] [CrossRef]
- Li, Y.; Lee, S.-H.; Le, Q.-T.; Kim, M.-M.; Kim, S.J. Anti-allergic effects of phlorotannins on histamine release via binding inhibition between IgE and Fc epsilonRI. J. Agric. Food Chem. 2008, 56, 12073–12080. [Google Scholar] [CrossRef]
- Le, Q.-T.; Li, Y.; Qian, Z.-J.; Kim, M.-M.; Kim, S.-K. Inhibitory effects of polyphenols isolated from marine alga Ecklonia cava on histamine release. Process Biochem. 2009, 44, 168–176. [Google Scholar] [CrossRef]
- Mayer, A.M.S.; Paul, V.J.; Fenical, W.; Norris, J.N.; de Carvalho, M.S.; Jacobs, R.S. Phospholipase A2 inhibitors from marine algae. Hydrobiologia 1993, 260, 521–529. [Google Scholar] [CrossRef]
- Ma, X.; Xie, B.; Du, J.; Zhang, A.; Hao, J.; Wang, S.; Wang, J.; Cao, G. The Anti-Inflammatory Effect and Structure of EPCP1-2 from Crypthecodinium cohnii via Modulation of TLR4-NF-κB Pathways in LPS-Induced RAW 264.7 Cells. Mar. Drugs 2017, 15, 376. [Google Scholar] [CrossRef]
- Bae, S.Y.; Yim, J.H.; Lee, H.K.; Pyo, S. Activation of murine peritoneal macrophages by sulfated exopolysaccharide from marine microalga Gyrodinium impudicum (strain KG03): Involvement of the NF-kappa B and JNK pathway. Int. Immunopharmacol. 2006, 6, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Kang, N.J.; Han, S.C.; Kang, G.J.; Koo, D.H.; Koh, Y.S.; Hyun, J.W.; Lee, N.H.; Ko, M.H.; Kang, H.K.; Yoo, E.S. Diphlorethohydroxycarmalol inhibits interleukin-6 production by regulating NF-kappaB, STAT5 and SOCS1 in lipopolysaccharide-stimulated RAW264.7 cells. Mar. Drugs 2015, 13, 2141–2157. [Google Scholar] [CrossRef] [PubMed]
- Kellogg, J.; Esposito, D.; Grace, M.; Komarnytsky, S.; Ann Lila, M. Alaskan seaweeds lower inflammation in RAW 264.7 macrophages and decrease lipid accumulation in 3T3-L1 adipocytes. J. Funct. Foods 2015, 15, 396–407. [Google Scholar] [CrossRef]
- Orsi, C.F.; Colombari, B.; Callegari, F.; Todaro, A.M.; Ardizzoni, A.; Rossini, G.P.; Blasi, E.; Peppoloni, S. Yessotoxin inhibits phagocytic activity of macrophages. Toxicon 2010, 55, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Alfonso, A.; Vieytes, M.R.; Botana, L.M. Yessotoxin, a Promising Therapeutic Tool. Mar. Drugs 2016, 14, 30. [Google Scholar] [CrossRef] [PubMed]
- San-Martin, A.; Rovirosa, J.; Astudillo, L.; Sepulveda, B.; Ruiz, D.; San-Martin, C. Biotransformation of the marine sesquiterpene pacifenol by a facultative marine fungus. Nat. Prod. Res. 2008, 22, 1627–1632. [Google Scholar] [CrossRef]
- Li, Y.X.; Li, Y.; Lee, S.H.; Qian, Z.J.; Kim, S.K. Inhibitors of oxidation and matrix metalloproteinases, floridoside, and D-isofloridoside from marine red alga Laurencia undulata. J. Agric. Food Chem. 2010, 58, 578–586. [Google Scholar] [CrossRef] [PubMed]
- Gil, B.; Ferrandiz, M.L.; Sanz, M.J.; Terencio, M.C.; Ubeda, A.; Rovirosa, J.; San-Martin, A.; Alcaraz, M.J.; Paya, M. Inhibition of inflammatory responses by epitaondiol and other marine natural products. Life Sci. 1995, 57, PL25–30. [Google Scholar] [CrossRef]
- Llanio, M.; Fernández, M.D.; Cabrera, B.; Bermejo, P.; Abad, M.; Payá, M.; Alcaraz, M.-J. The marine plant thalassia testudinum possesses anti-inflammatory and analgesic properties. Pharmacologyonline 2006, 3, 594–600. [Google Scholar]
- Jacobson, P.B.; Jacobs, R.S. Fuscoside: An anti-inflammatory marine natural product which selectively inhibits 5-lipoxygenase. Part I: Physiological and biochemical studies in murine inflammatory models. J. Pharmacol. Exp. Ther. 1992, 262, 866–873. [Google Scholar]
- Folmer, F.; Jaspars, M.; Schumacher, M.; Dicato, M.; Diederich, M. Marine natural products targeting phospholipases A2. Biochem. Pharmacol. 2010, 80, 1793–1800. [Google Scholar] [CrossRef] [PubMed]
- Keyzers, R.A.; Davies-Coleman, M.T. Anti-inflammatory metabolites from marine sponges. Chem. Soc. Rev. 2005, 34, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Potts, B.C.; Faulkner, D.J.; Jacobs, R.S. Phospholipase A2 inhibitors from marine organisms. J. Nat. Prod. 1992, 55, 1701–1717. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.-Y.; Cai, S.-X.; Zhang, W.-J.; Tang, X.-L.; Shin, H.-Y.; Lee, J.-Y.; Gu, Q.-Q.; Park, H. Semi-vioxanthin Isolated from marine-derived fungus regulates tumor necrosis factor-α, cluster of differentiation (CD) 80, CD86, and major histocompatibility complex class II expression in RAW264.7 cells via nuclear factor-kappa B and mitogen-activated protein kinase signaling pathways. Biol. Pharm. Bull. 2008, 31, 2228–2233. [Google Scholar] [PubMed]
- Yang, X.; Du, L.; Tang, X.; Jung, S.Y.; Zheng, B.; Soh, B.Y.; Kim, S.Y.; Gu, Q.; Park, H. Brevicompanine E reduces lipopolysaccharide-induced production of proinflammatory cytokines and enzymes in microglia by inhibiting activation of activator protein-1 and nuclear factor-kappaB. J. Neuroimmunol. 2009, 216, 32–38. [Google Scholar] [CrossRef]
- Wu, Q.X.; Crews, M.S.; Draskovic, M.; Sohn, J.; Johnson, T.A.; Tenney, K.; Valeriote, F.A.; Yao, X.J.; Bjeldanes, L.F.; Crews, P. Azonazine, a novel dipeptide from a Hawaiian marine sediment-derived fungus, Aspergillus insulicola. Org. Lett. 2010, 12, 4458–4461. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, Z.; Liu, H.; Pan, Y.; Li, J.; Liu, L.; She, Z. Dichloroisocoumarins with potential anti-Inflammatory activity from the mangrove endophytic fungus Ascomycota sp. CYSK-4. Mar. Drugs 2018, 16, 54. [Google Scholar] [CrossRef]
- Yang, X.; Kang, M.C.; Li, Y.; Kim, E.A.; Kang, S.M.; Jeon, Y.J. Anti-inflammatory activity of questinol isolated from marine-derived fungus Eurotium amstelodami in lipopolysaccharide-stimulated RAW 264.7 macrophages. J. Microbiol. Biotechnol. 2014, 24, 1346–1353. [Google Scholar] [CrossRef]
- Ayeka, P.A. Potential of mushroom compounds as immunomodulators in cancer immunotherapy: A review. Evid. Based Complement. Alternat. Med. 2018, 2018, 9. [Google Scholar] [CrossRef]
- Bandaranayake, W.M. Traditional and medicinal uses of mangroves. Mangroves Salt Marshes 1998, 2, 133–148. [Google Scholar] [CrossRef]
- Kathiresan, K. A review of studies on Pichavaram mangrove, southeast India. Hydrobiologia 2000, 430, 185–205. [Google Scholar] [CrossRef]
- Rahim, A.A.; Rocca, E.; Steinmetz, J.; Jain Kassim, M.; Sani Ibrahim, M.; Osman, H. Antioxidant activities of mangrove Rhizophora apiculata bark extracts. Food Chem. 2008, 107, 200–207. [Google Scholar] [CrossRef]
- Rahim, A.A.; Rocca, E.; Steinmetz, J.; Kassim, M.J.; Adnan, R.; Sani Ibrahim, M. Mangrove tannins and their flavanoid monomers as alternative steel corrosion inhibitors in acidic medium. Corros. Sci. 2007, 49, 402–417. [Google Scholar] [CrossRef]
- Agoramoorthy, G.; Chen, F.-A.; Venkatesalu, V.; Kuo, D.-H.; Shea, P.-C. Evaluation of antioxidant Polyphenols from selected mangrove plants of India. Asian J. Chem. 2008, 20, 1311. [Google Scholar]
- Premanathan, M.; Arakaki, R.; Izumi, H.; Kathiresan, K.; Nakano, M.; Yamamoto, N.; Nakashima, H. Antiviral properties of a mangrove plant, Rhizophora apiculata Blume, against human immunodeficiency virus. Antiviral Res. 1999, 44, 113–122. [Google Scholar] [CrossRef]
- Prabhu, V.V.; Guruvayoorappan, C. Anti-inflammatory and anti-tumor activity of the marine mangrove Rhizophora apiculata. J. Immunotoxicol. 2012, 9, 341–352. [Google Scholar] [CrossRef]
- Mehrotra, S.; Mishra, K.P.; Maurya, R.; Srimal, R.C.; Yadav, V.S.; Pandey, R.; Singh, V.K. Anticellular and immunosuppressive properties of ethanolic extract of Acorus calamus rhizome. Int. Immunopharmacol. 2003, 3, 53–61. [Google Scholar] [CrossRef]
- Alves, R.R.; Leo Neto, N.A.; Brooks, S.E.; Albuquerque, U.P. Commercialization of animal-derived remedies as complementary medicine in the semi-arid region of Northeastern Brazil. J. Ethnopharmacol. 2009, 124, 600–608. [Google Scholar] [CrossRef]
- Mayer, A.M.; Jacobson, P.B.; Fenical, W.; Jacobs, R.S.; Glaser, K.B. Pharmacological characterization of the pseudopterosins: Novel anti-inflammatory natural products isolated from the Caribbean soft coral, Pseudopterogorgia elisabethae. Life Sci. 1998, 62, PL401-7. [Google Scholar] [CrossRef]
- Chi, V.; Pennington, M.W.; Norton, R.S.; Tarcha, E.J.; Londono, L.M.; Sims-Fahey, B.; Upadhyay, S.K.; Lakey, J.T.; Iadonato, S.; Wulff, H.; et al. Development of a sea anemone toxin as an immunomodulator for therapy of autoimmune diseases. Toxicon 2012, 59, 529–546. [Google Scholar] [CrossRef]
- Ponkshe, C.A.; Indap, M.M. In vivo and in vitro evaluation for immunomodulatory activity of three marine animal extracts with reference to phagocytosis. Indian J. Exp. Biol. 2002, 40, 1399–1402. [Google Scholar]
- Liu, F.; Zhang, X.; Li, Y.; Chen, Q.; Liu, F.; Zhu, X.; Mei, L.; Song, X.; Liu, X.; Song, Z.; et al. Anti-Inflammatory effects of a Mytilus coruscus alpha-d-glucan (MP-A) in activated macrophage cells via TLR4/NF-kappaB/MAPK pathway Inhibition. Mar. Drugs 2017, 15, 294. [Google Scholar] [CrossRef]
- Monmai, C.; Go, S.H.; Shin, I.S.; You, S.G.; Lee, H.; Kang, S.B.; Park, W.J. Immune-enhancement and anti-Inflammatory activities of fatty acids extracted from Halocynthia aurantium tunic in RAW264.7 cells. Mar. Drugs 2018, 16, 309. [Google Scholar] [CrossRef]
- Nakajima, H.; Hamasaki, T.; Nishimura, K.; Kondo, T.; Kimura, Y.; Udagawa, S.-I.; Sato, S. Isolation of 2-acetylamino-3-hydroxy-4-methyloct-6-enoic acid, a derivative of the “C9-amino acid” residue of cyclosporins, produced by the fungus Neocosmospora vasinfecta E. F. Smith. Agric. Biol. Chem. 1988, 52, 1621–1623. [Google Scholar] [CrossRef]
- Cho, K.H.; Kim, D.C.; Yoon, C.S.; Ko, W.M.; Lee, S.J.; Sohn, J.H.; Jang, J.H.; Ahn, J.S.; Kim, Y.C.; Oh, H. Anti-neuroinflammatory effects of citreohybridonol involving TLR4-MyD88-mediated inhibition of NF-small ka, CyrillicB and MAPK signaling pathways in lipopolysaccharide-stimulated BV2 cells. Neurochem. Int. 2016, 95, 55–62. [Google Scholar] [CrossRef]
- Yoon, C.S.; Kim, D.C.; Lee, D.S.; Kim, K.S.; Ko, W.; Sohn, J.H.; Yim, J.H.; Kim, Y.C.; Oh, H. Anti-neuroinflammatory effect of aurantiamide acetate from the marine fungus Aspergillus sp. SF-5921: Inhibition of NF-kappaB and MAPK pathways in lipopolysaccharide-induced mouse BV2 microglial cells. Int. Immunopharmacol. 2014, 23, 568–574. [Google Scholar] [CrossRef]
- Xiao, A.J.; Chen, W.; Xu, B.; Liu, R.; Turlova, E.; Barszczyk, A.; Sun, C.L.; Liu, L.; Deurloo, M.; Wang, G.L.; et al. Marine compound xyloketal B reduces neonatal hypoxic-ischemic brain injury. Mar. Drugs 2014, 13, 29–47. [Google Scholar] [CrossRef]
- Kim, K.S.; Cui, X.; Lee, D.S.; Sohn, J.H.; Yim, J.H.; Kim, Y.C.; Oh, H. Anti-inflammatory effect of neoechinulin a from the marine fungus Eurotium sp. SF-5989 through the suppression of NF-small ka, CyrillicB and p38 MAPK Pathways in lipopolysaccharide-stimulated RAW264.7 macrophages. Molecules 2013, 18, 13245–13259. [Google Scholar] [CrossRef]
- Dou, H.; Song, Y.; Liu, X.; Gong, W.; Li, E.; Tan, R.; Hou, Y. Chaetoglobosin Fex from the marine-derived endophytic fungus inhibits induction of inflammatory mediators via Toll-like receptor 4 signaling in macrophages. Biol. Pharm. Bull. 2011, 34, 1864–1873. [Google Scholar] [CrossRef]
- Toledo, T.R.; Dejani, N.N.; Monnazzi, L.G.; Kossuga, M.H.; Berlinck, R.G.; Sette, L.D.; Medeiros, A.I. Potent anti-inflammatory activity of pyrenocine A isolated from the marine-derived fungus Penicillium paxilli Ma(G)K. Mediators Inflamm. 2014, 2014, 767061. [Google Scholar] [CrossRef]
- Kim, A.R.; Shin, T.S.; Lee, M.S.; Park, J.Y.; Park, K.E.; Yoon, N.Y.; Kim, J.S.; Choi, J.S.; Jang, B.C.; Byun, D.S.; et al. Isolation and identification of phlorotannins from Ecklonia stolonifera with antioxidant and anti-inflammatory properties. J. Agric. Food Chem. 2009, 57, 3483–3489. [Google Scholar] [CrossRef]
- Allavena, P.; Signorelli, M.; Chieppa, M.; Erba, E.; Bianchi, G.; Marchesi, F.; Olimpio, C.O.; Bonardi, C.; Garbi, A.; Lissoni, A.; et al. Anti-inflammatory properties of the novel antitumor agent yondelis (trabectedin): Inhibition of macrophage differentiation and cytokine production. Cancer Res. 2005, 65, 2964–2971. [Google Scholar] [CrossRef]
- Fast, M.D.; Johnson, S.C.; Eddy, T.D.; Pinto, D.; Ross, N.W. Lepeophtheirus salmonis secretory/excretory products and their effects on Atlantic salmon immune gene regulation. Parasite Immunol. 2007, 29, 179–189. [Google Scholar] [CrossRef]
- Campa-Cordova, A.I.; Hernandez-Saavedra, N.Y.; Ascencio, F. Superoxide dismutase as modulator of immune function in American white shrimp (Litopenaeus vannamei). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2002, 133, 557–565. [Google Scholar] [CrossRef]
- Akerkar, A.; Ponkshe, C.; Indap, M. Evaluation of immunomodulatory activity of extracts from marine animals. Indian J. Geo-Mar. Sci. 2009, 38, 22–27. [Google Scholar]
- Ovodova, R.G.; Glazkova, V.E.; Mikheyskaya, L.V.; Molchanova, V.I.; Isakov, V.V.; Ovodov, Y.S.; Fernandez Molina, L.E. The structure of mytilan, a bioglycan-immunomodulator isolated from the mussel Crenomytilus grayanus. Carbohydr. Res. 1992, 223, 221–226. [Google Scholar] [CrossRef]
- Fernandes, P.D.; Zardo, R.S.; Figueiredo, G.S.; Silva, B.V.; Pinto, A.C. Anti-inflammatory properties of convolutamydine A and two structural analogues. Life Sci. 2014, 116, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.Y.; Hsu, C.H.; Chao, C.H.; Wen, Z.H.; Wu, Y.C.; Dai, C.F.; Sheu, J.H. Cytotoxic and anti-inflammatory metabolites from the soft coral Scleronephthya gracillimum. Mar. Drugs 2013, 11, 1853–1865. [Google Scholar] [CrossRef] [PubMed]
- Pereira, D.M.; Correia-da-Silva, G.; Valentao, P.; Teixeira, N.; Andrade, P.B. Anti-inflammatory effect of unsaturated fatty acids and Ergosta-7,22-dien-3-ol from Marthasterias glacialis: Prevention of CHOP-mediated ER-stress and NF-kappaB activation. PLoS ONE 2014, 9, e88341. [Google Scholar] [CrossRef] [PubMed]
- Thao, N.P.; Cuong, N.X.; Luyen, B.T.; Quang, T.H.; Hanh, T.T.; Kim, S.; Koh, Y.S.; Nam, N.H.; Van Kiem, P.; Van Minh, C.; et al. Anti-inflammatory components of the starfish Astropecten polyacanthus. Mar. Drugs 2013, 11, 2917–2926. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.K.; Duh, C.Y. New cytotoxic cembranolides from the soft coral Lobophytum michaelae. Mar. Drugs 2012, 10, 306–318. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.C.; Wen, Z.-H.; Chao, C.H.; Ahmed, A.; Su, J.H.; Chiang, M.; Kuo, Y.-H.; Hsu, C.H.; Sheu, J.-H. Novel sesquiterenoids from the Formosan soft coral Paralemnalia thyrsoides. Tetrahedron Lett. 2006, 47, 8751–8755. [Google Scholar] [CrossRef]
- Chen, B.W.; Chang, S.M.; Huang, C.Y.; Chao, C.H.; Su, J.H.; Wen, Z.H.; Hsu, C.H.; Dai, C.F.; Wu, Y.C.; Sheu, J.H. Hirsutalins A-H, eunicellin-based diterpenoids from the soft coral Cladiella hirsuta. J. Nat. Prod. 2010, 73, 1785–1791. [Google Scholar] [CrossRef]
- Quang, T.H.; Ha, T.T.; Minh, C.V.; Kiem, P.V.; Huong, H.T.; Ngan, N.T.T.; Nhiem, N.X.; Tung, N.H.; Tai, B.H.; Thuy, D.T.T.; et al. Cytotoxic and anti-inflammatory cembranoids from the Vietnamese soft coral Lobophytum laevigatum. Biorg. Med. Chem. 2011, 19, 2625–2632. [Google Scholar] [CrossRef]
- Ahmed, A.; Hsieh, Y.-T.; Wen, Z.-H.; Wu, Y.-C.; Sheu, J.-H. Polyoxygenated Sterols from the Formosan Soft Coral Sinularia gibberosa. J. Nat. Prod. 2006, 69, 1275–1279. [Google Scholar] [CrossRef]
- Reina, E.; Puentes, C.; Rojas, J.; Garcia, J.; Ramos, F.A.; Castellanos, L.; Aragon, M.; Ospina, L.F. Fuscoside E: A strong anti-inflammatory diterpene from Caribbean octocoral Eunicea fusca. Bioorg. Med. Chem. Lett. 2011, 21, 5888–5891. [Google Scholar] [CrossRef]
- Marchbank, D.H.; Kerr, R.G. Semisynthesis of fuscoside B analogues and eunicosides, and analysis of anti-inflammatory activity. Tetrahedron 2011, 67, 3053–3061. [Google Scholar] [CrossRef]
- Voultsiadou, E. Therapeutic properties and uses of marine invertebrates in the ancient Greek world and early Byzantium. J. Ethnopharmacol. 2010, 130, 237–247. [Google Scholar] [CrossRef]
- Proksch, P. Chinese Marine Materia Medica. By Huashi Guan and Shuguang Wang. Shanghai Scientific and Technical Publishers, China Ocean Press, and Chemical Industry Press: Shanghai, Beijing, China, 2009; Hardback, 7064 pp; ¥ 2920; ISBN 978-7-5323-9958-1/R•2707; ISBN 978-7-5323-9973-4/R•2708; ISBN 978-7-1220-6012-9. Mar. Drugs 2014, 3, 193–195. [Google Scholar]
- Joung, H.-J.; Kim, Y.-S.; Hwang, J.-W.; Han, Y.-K.; Jeong, J.-H.; Lee, J.-S.; Moon, S.-H.; Jeon, B.-T.; Park, P.-J. Anti-inflammatory effects of extract from Haliotis discus hannai fermented with Cordyceps militaris mycelia in RAW264.7 macrophages through TRIF-dependent signaling pathway. Fish Shellfish Immunol. 2014, 38, 184–189. [Google Scholar] [CrossRef]
- Jean, Y.H.; Chen, W.F.; Sung, C.S.; Duh, C.Y.; Huang, S.Y.; Lin, C.S.; Tai, M.H.; Tzeng, S.F.; Wen, Z.H. Capnellene, a natural marine compound derived from soft coral, attenuates chronic constriction injury-induced neuropathic pain in rats. Br. J. Pharmacol. 2009, 158, 713–725. [Google Scholar] [CrossRef]
- Chen, Z.C.; Wu, S.S.; Su, W.Y.; Lin, Y.C.; Lee, Y.H.; Wu, W.H.; Chen, C.H.; Wen, Z.H. Anti-inflammatory and burn injury wound healing properties of the shell of Haliotis diversicolor. BMC Complement. Altern. Med. 2016, 16, 487. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Chakraborty, M.; Bose, M.; Mukherjee, D.; Roychoudhury, A.; Dhar, P.; Mishra, R. Indian freshwater edible snail Bellamya bengalensis lipid extract prevents T cell mediated hypersensitivity and inhibits LPS induced macrophage activation. J. Ethnopharmacol. 2014, 157, 320–329. [Google Scholar] [CrossRef]
- Ahmad, T.B.; Rudd, D.; Smith, J.; Kotiw, M.; Mouatt, P.; Seymour, L.M.; Liu, L.; Benkendorff, K. Anti-Inflammatory Activity and Structure-Activity Relationships of Brominated Indoles from a Marine Mollusc. Mar. Drugs 2017, 15, 133. [Google Scholar] [CrossRef]
- Lawson, B.R.; Belkowski, S.M.; Whitesides, J.F.; Davis, P.; Lawson, J.W. Immunomodulation of murine collagen-induced arthritis by N, N-dimethylglycine and a preparation of Perna canaliculus. BMC Complement. Altern. Med. 2007, 7, 20. [Google Scholar] [CrossRef]
- Wu, Y.; Hu, X.; Song, L.; Zhu, J.; Yu, R. The inhibitory effect of a novel polypeptide fraction from Arca subcrenata on cancer-related inflammation in human cervical cancer HeLa cells. ScientificWorldJournal 2014, 2014, 768938. [Google Scholar]
- Arancibia, S.; Espinoza, C.; Salazar, F.; Del Campo, M.; Tampe, R.; Zhong, T.Y.; De Ioannes, P.; Moltedo, B.; Ferreira, J.; Lavelle, E.C.; et al. A novel immunomodulatory hemocyanin from the limpet Fissurella latimarginata promotes potent anti-tumor activity in melanoma. PLoS ONE 2014, 9, e87240. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Fu, Y.; Zheng, J.; Li, D. Anti-inflammatory activity and mechanism of a lipid extract from hard-shelled mussel (Mytilus coruscus) on chronic arthritis in rats. Mar. Drugs 2014, 12, 568–588. [Google Scholar] [CrossRef] [PubMed]
- Fei, L.; Xu, K. Zhikang Capsule ameliorates dextran sodium sulfate-induced colitis by inhibition of inflammation, apoptosis, oxidative stress and MyD88-dependent TLR4 signaling pathway. J. Ethnopharmacol. 2016, 192, 236–247. [Google Scholar] [CrossRef] [PubMed]
- Miles, E.A.; Calder, P.C. Influence of marine n-3 polyunsaturated fatty acids on immune function and a systematic review of their effects on clinical outcomes in rheumatoid arthritis. Br. J. Nutr. 2012, 107, S171–S184. [Google Scholar] [CrossRef]
- Lillsunde, K.E.; Festa, C.; Adel, H.; De Marino, S.; Lombardi, V.; Tilvi, S.; Nawrot, D.A.; Zampella, A.; D’Souza, L.; DAuria, M.V.; et al. Bioactive cembrane derivatives from the Indian Ocean soft coral, Sinularia kavarattiensis. Mar. Drugs 2014, 12, 4045–4068. [Google Scholar] [CrossRef]
- Liu, C.Y.; Hwang, T.L.; Lin, M.R.; Chen, Y.H.; Chang, Y.C.; Fang, L.S.; Wang, W.H.; Wu, Y.C.; Sung, P.J. Carijoside A, a bioactive sterol glycoside from an octocoral Carijoa sp. (Clavulariidae). Mar. Drugs 2010, 8, 2014–2020. [Google Scholar] [CrossRef]
- Cheng, S.Y.; Chuang, C.T.; Wen, Z.H.; Wang, S.K.; Chiou, S.F.; Hsu, C.H.; Dai, C.F.; Duh, C.Y. Bioactive norditerpenoids from the soft coral Sinularia gyrosa. Bioorg. Med. Chem. 2010, 18, 3379–3386. [Google Scholar] [CrossRef]
- Chen, W.-F.; Chakraborty, C.; Sung, C.-S.; Feng, C.-W.; Jean, Y.-H.; Lin, Y.-Y.; Hung, H.-C.; Huang, T.-Y.; Huang, S.-Y.; Su, T.-M.; et al. Neuroprotection by marine-derived compound, 11-dehydrosinulariolide, in an in vitro Parkinson’s model: A promising candidate for the treatment of Parkinson’s disease. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2012, 385, 265–275. [Google Scholar] [CrossRef]
- Chen, B.W.; Chao, C.H.; Su, J.H.; Wen, Z.H.; Sung, P.J.; Sheu, J.H. Anti-inflammatory eunicellin-based diterpenoids from the cultured soft coral Klyxum simplex. Org. Biomol. Chem. 2010, 8, 2363–2366. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-L.; Su, J.-H.; Wen, Z.-H.; Hsu, C.-H.; Chen, B.-W.; Dai, C.-F.; Kuo, Y.-H.; Sheu, J.-H. Simplexins A-I, Eunicellin-based Diterpenoids from the soft coral klyxum simplex. J. Nat. Prod. 2009, 72, 994–1000. [Google Scholar] [CrossRef] [PubMed]
- Wanzola, M.; Furuta, T.; Kohno, Y.; Fukumitsu, S.; Yasukochi, S.; Watari, K.; Tanaka, C.; Higuchi, R.; Miyamoto, T. Four new cembrane diterpenes isolated from an Okinawan soft coral Lobophytum crassum with inhibitory effects on nitric oxide production. Chem. Pharm. Bull. (Tokyo) 2010, 58, 1203–1209. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.Y.; Wen, Z.H.; Wang, S.K.; Chiou, S.F.; Hsu, C.H.; Dai, C.F.; Chiang, M.Y.; Duh, C.Y. Unprecedented hemiketal cembranolides with anti-inflammatory activity from the soft coral Lobophytum durum. J. Nat. Prod. 2009, 72, 152–155. [Google Scholar] [CrossRef] [PubMed]
- Belmiro, C.L.; Castelo-Branco, M.T.; Melim, L.M.; Schanaider, A.; Elia, C.; Madi, K.; Pavao, M.S.; de Souza, H.S. Unfractionated heparin and new heparin analogues from ascidians (chordate-tunicate) ameliorate colitis in rats. J. Biol. Chem. 2009, 284, 11267–11278. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.Y.; Wen, Z.H.; Chiou, S.F.; Tsai, C.W.; Wang, S.K.; Hsu, C.H.; Dai, C.F.; Chiang, M.Y.; Wang, W.H.; Duh, C.Y. Ceramide and cerebrosides from the octocoral Sarcophyton ehrenbergi. J. Nat. Prod. 2009, 72, 465–468. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.C.; Huang, S.Y.; Jean, Y.H.; Chen, W.F.; Sung, C.S.; Kao, E.S.; Wang, H.M.; Chakraborty, C.; Duh, C.Y.; Wen, Z.H. Intrathecal lemnalol, a natural marine compound obtained from Formosan soft coral, attenuates nociceptive responses and the activity of spinal glial cells in neuropathic rats. Behav. Pharmacol. 2011, 22, 739–750. [Google Scholar] [CrossRef]
- Lin, W.Y.; Su, J.H.; Lu, Y.; Wen, Z.H.; Dai, C.F.; Kuo, Y.H.; Sheu, J.H. Cytotoxic and anti-inflammatory cembranoids from the Dongsha Atoll soft coral Sarcophyton crassocaule. Bioorg. Med. Chem. 2010, 18, 1936–1941. [Google Scholar] [CrossRef]
- Appleton, D.R.; Chuen, C.S.; Berridge, M.V.; Webb, V.L.; Copp, B.R. Rossinones A and B, biologically active meroterpenoids from the Antarctic ascidian, Aplidium species. J. Org. Chem. 2009, 74, 9195–9198. [Google Scholar] [CrossRef]
- Cheng, S.-Y.; Huang, Y.-C.; Wen, Z.-H.; Chiou, S.-F.; Wang, S.-K.; Hsu, C.-H.; Dai, C.-F.; Duh, C.-Y. Novel sesquiterpenes and norergosterol from the soft corals Nephthea erecta and Nephtheachabroli. Tetrahedron Lett. 2009, 50, 802–806. [Google Scholar] [CrossRef]
- Hanif, N.; Ohno, O.; Kitamura, M.; Yamada, K.; Uemura, D. Symbiopolyol, a VCAM-1 inhibitor from a symbiotic dinoflagellate of the jellyfish Mastigias papua. J. Nat. Prod. 2010, 73, 1318–1322. [Google Scholar] [CrossRef]
- Lu, Y.; Zhao, L.X.; Cao, D.L.; Gao, Y.J. Spinal injection of docosahexaenoic acid attenuates carrageenan-induced inflammatory pain through inhibition of microglia-mediated neuroinflammation in the spinal cord. Neuroscience 2013, 241, 22–31. [Google Scholar] [CrossRef]
- Abad, M.J.; Bermejo, P. Bioactive natural products from marine sources. Stud. Nat. Prod. Chem. 2001, 25, 683–755. [Google Scholar]
- Laurienzo, P. Marine polysaccharides in pharmaceutical applications: An overview. Mar. Drugs 2010, 8, 2435–2465. [Google Scholar] [CrossRef]
- Gorelik, E.; Bere, W.W.; Herberman, R.B. Role of NK cells in the antimetastatic effect of anticoagulant drugs. Int. J. Cancer 1984, 33, 87–94. [Google Scholar] [CrossRef]
- Gorelik, E. Augmentation of the antimetastatic effect of anticoagulant drugs by immunostimulation in mice. Cancer Res. 1987, 47, 809–815. [Google Scholar]
- Zhou, G.; Xin, H.; Sheng, W.; Sun, Y.; Li, Z.; Xu, Z. In vivo growth-inhibition of S180 tumor by mixture of 5-Fu and low molecular lambda-carrageenan from Chondrus ocellatus. Pharmacol. Res. 2005, 51, 153–157. [Google Scholar] [CrossRef]
- Cumashi, A.; Ushakova, N.A.; Preobrazhenskaya, M.E.; D’Incecco, A.; Piccoli, A.; Totani, L.; Tinari, N.; Morozevich, G.E.; Berman, A.E.; Bilan, M.I.; et al. Consorzio Interuniversitario Nazionale per la Bio-Oncologia, I. A comparative study of the anti-inflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology 2007, 17, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Luscher-Mattli, M. Polyanions—A lost chance in the fight against HIV and other virus diseases? Antivir. Chem. Chemother. 2000, 11, 249–259. [Google Scholar] [CrossRef]
- Li, B.; Lu, F.; Wei, X.; Zhao, R. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar] [CrossRef]
- Jiao, G.; Yu, G.; Zhang, J.; Ewart, H.S. Chemical structures and bioactivities of sulfated polysaccharides from marine algae. Mar. Drugs 2011, 9, 196–223. [Google Scholar] [CrossRef]
- Kinnel, R.B.; Esquenazi, E.; Leao, T.; Moss, N.; Mevers, E.; Pereira, A.R.; Monroe, E.A.; Korobeynikov, A.; Murray, T.F.; Sherman, D.; et al. A Maldiisotopic approach to discover natural products: Cryptomaldamide, a hybrid tripeptide from the marine cyanobacterium Moorea producens. J. Nat. Prod. 2017, 80, 1514–1521. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.H.; Lee, S.A.; Moon, P.D.; Na, H.J.; Park, R.K.; Um, J.; Kim, H.M.; Hong, S.H. Alginic acid has anti-anaphylactic effects and inhibits inflammatory cytokine expression via suppression of nuclear factor-κB activation. Clin. Exp. Allergy 2006, 36, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Nicolaus, B.; Kambourova, M.; Oner, E.T. Exopolysaccharides from extremophiles: From fundamentals to biotechnology. Environ. Technol. 2010, 31, 1145–1158. [Google Scholar] [CrossRef] [PubMed]
- Guezennec, J. From extreme environments to biologically active exopolysaccharides. Commun. Agric. Appl. Biol. Sci. 2003, 68, 227–234. [Google Scholar]
- Maugeri, T.L.; Gugliandolo, C.; Caccamo, D.; Stackebrandt, E. Three novel halotolerant and thermophilic Geobacillus strains from shallow marine vents. Syst. Appl. Microbiol. 2002, 25, 450–455. [Google Scholar] [CrossRef]
- Poli, A.; Anzelmo, G.; Nicolaus, B. Bacterial exopolysaccharides from extreme marine habitats: Production, characterization and biological activities. Mar. Drugs 2010, 8, 1779–1802. [Google Scholar] [CrossRef]
- Gugliandolo, C.; Lentini, V.; Spanò, A.; Maugeri, T.L. New bacilli from shallow hydrothermal vents of Panarea Island (Italy) and their biotechnological potential. J. Appl. Microbiol. 2012, 112, 1102–1112. [Google Scholar] [CrossRef]
- Dembitsky, V.M. Bromo- and iodo-containing alkaloids from marine microorganisms and sponges. Bioorg. Khim 2002, 28, 196–208. [Google Scholar]
- Guven, K.C.; Percot, A.; Sezik, E. Alkaloids in marine algae. Mar. Drugs 2010, 8, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Dewapriya, P.; Li, Y.X.; Himaya, S.W.; Pangestuti, R.; Kim, S.K. Neoechinulin A suppresses amyloid-beta oligomer-induced microglia activation and thereby protects PC-12 cells from inflammation-mediated toxicity. Neurotoxicology 2013, 35, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Bruhn, J.G.; Bruhn, C. Alkaloids and ethnobotany of Mexican peyote cacti and related species. Econ. Bot. 1973, 27, 241. [Google Scholar] [CrossRef]
- Harborne, J.B.; Williams, C.A. Advances in flavonoid research since 1992. Phytochemistry 2000, 55, 481–504. [Google Scholar] [CrossRef]
- Bravo, L. Polyphenols: Chemistry, dietary sources, metabolism, and nutritional significance. Nutr. Rev. 1998, 56, 317–333. [Google Scholar] [CrossRef]
- Cheynier, V. Polyphenols in food are more complex than often thought. Am. J. Clin. Nutr. 2005, 81, 223S–229S. [Google Scholar] [CrossRef]
- Heim, K.E.; Tagliaferro, A.R.; Bobilya, D.J. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar] [CrossRef]
- Andriantsitohaina, R.; Auger, C.; Chataigneau, T.; Etienne-Selloum, N.; Li, H.; Martinez, M.C.; Schini-Kerth, V.B.; Laher, I. Molecular mechanisms of the cardiovascular protective effects of polyphenols. Br. J. Nutr. 2012, 108, 1532–1549. [Google Scholar] [CrossRef]
- Middleton, E., Jr.; Kandaswami, C.; Theoharides, T.C. The effects of plant flavonoids on mammalian cells: Implications for inflammation, heart disease, and cancer. Pharmacol. Rev. 2000, 52, 673–751. [Google Scholar]
- Recio, M.C.; Andujar, I.; Rios, J.L. Anti-inflammatory agents from plants: Progress and potential. Curr. Med. Chem. 2012, 19, 2088–2103. [Google Scholar] [CrossRef]
- Puupponen-Pimia, R.; Nohynek, L.; Meier, C.; Kahkonen, M.; Heinonen, M.; Hopia, A.; Oksman-Caldentey, K.M. Antimicrobial properties of phenolic compounds from berries. J. Appl. Microbiol. 2001, 90, 494–507. [Google Scholar] [CrossRef]
- Giovannini, C.; Scazzocchio, B.; Vari, R.; Santangelo, C.; D’Archivio, M.; Masella, R. Apoptosis in cancer and atherosclerosis: Polyphenol activities. Ann. Ist. Super. Sanita 2007, 43, 406–416. [Google Scholar]
- Aquilano, K.; Baldelli, S.; Rotilio, G.; Ciriolo, M.R. Role of nitric oxide synthases in Parkinson’s disease: A review on the antioxidant and anti-inflammatory activity of polyphenols. Neurochem. Res. 2008, 33, 2416–2426. [Google Scholar] [CrossRef]
- Villegas, I.; Sanchez-Fidalgo, S.; Alarcon de la Lastra, C. New mechanisms and therapeutic potential of curcumin for colorectal cancer. Mol. Nutr. Food Res. 2008, 52, 1040–1061. [Google Scholar] [CrossRef]
- Shakibaei, M.; Harikumar, K.B.; Aggarwal, B.B. Resveratrol addiction: To die or not to die. Mol. Nutr. Food Res. 2009, 53, 115–128. [Google Scholar] [CrossRef]
- Sun, C.; Hu, Y.; Liu, X.; Wu, T.; Wang, Y.; He, W.; Wei, W. Resveratrol downregulates the constitutional activation of nuclear factor-kappaB in multiple myeloma cells, leading to suppression of proliferation and invasion, arrest of cell cycle, and induction of apoptosis. Cancer Genet. Cytogenet. 2006, 165, 9–19. [Google Scholar] [CrossRef]
- Holmes-McNary, M.; Baldwin, A.S., Jr. Chemopreventive properties of trans-resveratrol are associated with inhibition of activation of the IkappaB kinase. Cancer Res. 2000, 60, 3477–3483. [Google Scholar]
- Adhami, V.M.; Afaq, F.; Ahmad, N. Suppression of ultraviolet B exposure-mediated activation of NF-kappaB in normal human keratinocytes by resveratrol. Neoplasia 2003, 5, 74–82. [Google Scholar] [CrossRef]
- Manna, S.K.; Mukhopadhyay, A.; Aggarwal, B.B. Resveratrol suppresses TNF-induced activation of nuclear transcription factors NF-kappa B, activator protein-1, and apoptosis: Potential role of reactive oxygen intermediates and lipid peroxidation. J. Immunol. 2000, 164, 6509–6519. [Google Scholar] [CrossRef]
- Jovanović-Šanta, S.S.; Petri, E.T.; Klisurić, O.R.; Szécsi, M.; Kovačević, R.; Petrović, J.A. Antihormonal potential of selected D-homo and D-seco estratriene derivatives. Steroids 2015, 97, 45–53. [Google Scholar] [CrossRef]
- Lopez, L.M.; Grimes, D.A.; Schulz, K.F.; Curtis, K.M. Steroidal contraceptives: Effect on bone fractures in women. Cochrane Database Syst. Rev. 2011, 6, CD006033. [Google Scholar]
- Rattanasopa, C.; Phungphong, S.; Wattanapermpool, J.; Bupha-Intr, T. Significant role of estrogen in maintaining cardiac mitochondrial functions. J. Steroid Biochem. Mol. Biol. 2015, 147, 1–9. [Google Scholar] [CrossRef]
- Thao, N.P.; Luyen, B.T.T.; Kim, E.J.; Kang, J.I.; Kang, H.K.; Cuong, N.X.; Nam, N.H.; Kiem, P.V.; Minh, C.V.; Kim, Y.H. Steroidal Constituents from the Edible Sea Urchin Diadema savignyi Michelin Induce Apoptosis in Human Cancer Cells. J. Med. Food 2014, 18, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Cortet, B.; Biver, E.; Borg, S.; Chopin, F.; Hoppé, E.; Laroche, M.; Morel, G.; Razjbaum, G.; Roux, C.; Thomas, T.; et al. Management of male osteoporosis: Lessons for clinical practice. Joint Bone Spine 2011, 78, S208–S210. [Google Scholar] [CrossRef]
- Aav, R.; Kanger, T.; Pehk, T.; Lopp, M. Unexpected reactivity of ethyl 2-(Diethylphosphono)propionate toward 2,2-disubstituted-1,3-cyclopentanediones. Phosphorus Sulfur Silicon Related Elem. 2005, 180, 1739–1748. [Google Scholar] [CrossRef]
- Fiorucci, S.; Distrutti, E.; Bifulco, G.; D’Auria, M.V.; Zampella, A. Marine sponge steroids as nuclear receptor ligands. Trends Pharmacol. Sci. 2012, 33, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Sepe, V.; Ummarino, R.; D’Auria, M.V.; Mencarelli, A.; D’Amore, C.; Renga, B.; Zampella, A.; Fiorucci, S. Total synthesis and pharmacological characterization of solomonsterol A, a potent marine pregnane-X-receptor agonist endowed with anti-inflammatory activity. J. Med. Chem. 2011, 54, 4590–4599. [Google Scholar] [CrossRef] [PubMed]
- Mencarelli, A.; D’Amore, C.; Renga, B.; Cipriani, S.; Carino, A.; Sepe, V.; Perissutti, E.; D’Auria, M.V.; Zampella, A.; Distrutti, E.; et al. Solomonsterol A, a marine pregnane-X-receptor agonist, attenuates inflammation and immune dysfunction in a mouse model of arthritis. Mar. Drugs 2013, 12, 36–53. [Google Scholar] [CrossRef]
- Behl, C.; Moosmann, B. Antioxidant neuroprotection in Alzheimer’s disease as preventive and therapeutic approach22This article is part of a series of reviews on “Causes and Consequences of Oxidative Stress in Alzheimer’s Disease.” The full list of papers may be found on the homepage of the journal. Free Radical Biol. Med. 2002, 33, 182–191. [Google Scholar]
- Snow, B.J.; Rolfe, F.L.; Lockhart, M.M.; Frampton, C.M.; O’Sullivan, J.D.; Fung, V.; Smith, R.A.; Murphy, M.P.; Taylor, K.M.; Protect Study, G. A double-blind, placebo-controlled study to assess the mitochondria-targeted antioxidant MitoQ as a disease-modifying therapy in Parkinson’s disease. Mov. Disord. 2010, 25, 1670–1674. [Google Scholar] [CrossRef]
- Amaro, S.; Chamorro, A. Translational stroke research of the combination of thrombolysis and antioxidant therapy. Stroke 2011, 42, 1495–1499. [Google Scholar] [CrossRef]
- Fuchs-Tarlovsky, V. Role of antioxidants in cancer therapy. Nutrition 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Ishibashi, T. Molecular hydrogen: New antioxidant and anti-inflammatory therapy for rheumatoid arthritis and related diseases. Curr. Pharm. Des. 2013, 19, 6375–6381. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Davidson, B.P.; Yue, Q.; Belcik, T.; Xie, A.; Inaba, Y.; McCarty, O.J.; Tormoen, G.W.; Zhao, Y.; Ruggeri, Z.M.; et al. Molecular imaging of inflammation and platelet adhesion in advanced atherosclerosis effects of antioxidant therapy with NADPH oxidase inhibition. Circ. Cardiovasc. Imaging 2013, 6, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Meydani, S.N.; Meydani, M.; Verdon, C.P.; Shapiro, A.A.; Blumberg, J.B.; Hayes, K.C. Vitamin E supplementation suppresses prostaglandin E1(2) synthesis and enhances the immune response of aged mice. Mech. Ageing Dev. 1986, 34, 191–201. [Google Scholar] [CrossRef]
- Meydani, S.N.; Barklund, M.P.; Liu, S.; Meydani, M.; Miller, R.A.; Cannon, J.G.; Morrow, F.D.; Rocklin, R.; Blumberg, J.B. Vitamin E supplementation enhances cell-mediated immunity in healthy elderly subjects. Am. J. Clin. Nutr. 1990, 52, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Pekmezci, D. Vitamin E and immunity. Vitam. Horm. 2011, 86, 179–215. [Google Scholar] [PubMed]
- Salimian, J.; Arefpour, M.A.; Riazipour, M.; Poursasan, N. Immunomodulatory effects of selenium and vitamin E on alterations in T lymphocyte subsets induced by T-2 toxin. Immunopharmacol. Immunotoxicol. 2014, 36, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Guerin, M.; Huntley, M.E.; Olaizola, M. Haematococcus astaxanthin: Applications for human health and nutrition. Trends Biotechnol. 2003, 21, 210–216. [Google Scholar] [CrossRef]
- Hussein, G.; Sankawa, U.; Goto, H.; Matsumoto, K.; Watanabe, H. Astaxanthin, a carotenoid with potential in human health and nutrition. J. Nat. Prod. 2006, 69, 443–449. [Google Scholar] [CrossRef]
- Miyashita, K.A.H.M. Antiobesity Effect of Allenic Carotenoid, Fucoxanthin; Wiley-Blackwell: Hoboken, NJ, USA, 2009; pp. 145–160. [Google Scholar]
- Olaizola, M. The Production and Health Benefits of Astaxanthin. In Marine Nutraceuticals and Functional Foods; Barrow, C., Shahidi, F., Eds.; CRC Press: Boca Raton, FL, USA, 2007; p. 321. [Google Scholar]
- Miyashita, K.; Hosokawa, M. Beneficial Health Effects of Seaweed Carotenoid, Fucoxanthin. In Marine Nutraceuticals and Functional Foods; Barrow, C., Shahidi, F., Eds.; CRC Press: Boca Raton, FL, USA, 2008. [Google Scholar]
- Ohgami, K.; Shiratori, K.; Kotake, S.; Nishida, T.; Mizuki, N.; Yazawa, K.; Ohno, S. Effects of astaxanthin on lipopolysaccharide-induced inflammation in vitro and in vivo. Invest. Ophthalmol. Vis. Sci. 2003, 44, 2694–2701. [Google Scholar] [CrossRef]
- Tan, C.-P.; Hou, Y.-H. First Evidence for the Anti-inflammatory Activity of Fucoxanthin in High-Fat-Diet-Induced Obesity in Mice and the Antioxidant Functions in PC12 Cells. Inflammation 2014, 37, 443–450. [Google Scholar] [CrossRef]
- Sun, P.; Li, D.; Dong, B.; Qiao, S.; Ma, X.; Chen, X. Vitamin C: An immunomodulator that attenuates anaphylactic reactions to soybean glycinin hypersensitivity in a swine model. Food Chem. 2009, 113, 914–918. [Google Scholar] [CrossRef]
- Tenorio-Rodríguez, P.; Méndez-Rodrìguez, L.; Serviere-Zaragoza, E.; OHara, T.; Zenteno-Savín, T. Antioxidant substances and trace element content in macroalgae from a subtropical lagoon in the West Coast of the Baja California Peninsula. Vitam Trace Elem 2013, 2, 2167-0390. [Google Scholar]
- Cornish, M.; Garbary, D. Antioxidants from macroalgae: Potential applications in human health and nutrition. Algae 2010, 25, 155–171. [Google Scholar] [CrossRef]
- Mohite, O.S.; Weber, T.; Kim, H.U.; Lee, S.Y. Genome-Scale Metabolic Reconstruction of Actinomycetes for Antibiotics Production. Biotechnol. J. 2019, 14, 1800377. [Google Scholar] [CrossRef]
- Salcedo, R.G.; Olano, C.; Gómez, C.; Fernández, R.; Braña, A.F.; Méndez, C.; de la Calle, F.; Salas, J.A. Characterization and engineering of the biosynthesis gene cluster for antitumor macrolides PM100117 and PM100118 from a marine actinobacteria: Generation of a novel improved derivative. Microb. Cell Factories 2016, 15, 44. [Google Scholar] [CrossRef]
- Gassel, S.; Schewe, H.; Schmidt, I.; Schrader, J.; Sandmann, G. Multiple improvement of astaxanthin biosynthesis in Xanthophyllomyces dendrorhous by a combination of conventional mutagenesis and metabolic pathway engineering. Biotechnol. Lett. 2013, 35, 565–569. [Google Scholar] [CrossRef]
- Blazeck, J.; Alper, H. Systems metabolic engineering: Genome-scale models and beyond. Biotechnol. J. 2010, 5, 647–659. [Google Scholar] [CrossRef]
- Vijayakumar, S.; Conway, M.; Lió, P.; Angione, C. Optimization of Multi-Omic Genome-Scale Models: Methodologies, Hands-on Tutorial, and Perspectives. In Metabolic Network Reconstruction and Modeling: Methods and Protocols; Fondi, M., Ed.; Springer: New York, NY, USA, 2018; pp. 389–408. [Google Scholar]
- Medema, M.H.; Fischbach, M.A. Computational approaches to natural product discovery. Nat. Chem. Biol. 2015, 11, 639–648. [Google Scholar] [CrossRef]
- Zhao, X.Q. Genome-based studies of marine microorganisms to maximize the diversity of natural products discovery for medical treatments. Evid. Based Complement. Alternat. Med. 2011, 2011, 384572. [Google Scholar] [CrossRef]
- Li, Y.; Lin, Z.; Huang, C.; Zhang, Y.; Wang, Z.; Tang, Y.-J.; Chen, T.; Zhao, X. Metabolic engineering of Escherichia coli using CRISPR–Cas9 meditated genome editing. Metab. Eng. 2015, 31, 13–21. [Google Scholar] [CrossRef]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex genome engineering using CRISPR/Cas systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Structure of anti-inflammatory and immunomodulatory marine-derived compounds.

Figure 2.
Schematic diagram showing omics data analysis and genome-scale metabolic modelling for improvement of production.
Figure 2.
Schematic diagram showing omics data analysis and genome-scale metabolic modelling for improvement of production.

Table 1.
Marine bacteria and their therapeutic chemical constituents.
| Bacterial Species | Chemical(s) | Immunomodulatory Activity | Ref(s) |
|---|---|---|---|
| Aplidium albicus | Cyclic depsipeptide | In vivo activity in the plasmocytoma murine model of xenograft. Applidine exhibits antimyeloma activity in vivo. | [30] |
| Nocardiopsis sp. K-252, Nonomuraea longicatena | Lestaurtinib (Alkaloid) | Potent PKC and calmodulin inhibitor. Prevents myelin oligoglycoprotein induced encephalomyelitis in vivo. | [31] |
| Bryozoa neritina | Bryostatin polyketide | Exhibits antitumor activity against malignant melanoma. IL-6 and TNF-α levels rise in patients within 24 hours of the treatment. | [32,33] |
| Micrococcus luteus | Anti-Micrococcus luteus antibodies | Immunosuppressive potential through the expansion of immunoregulatory T cell subsets. | [34] |
| Trididemnum solidum | Didemnin B (Depsipeptides) | Exhibits strong anti-inflammatory and immunosuppressive activity. The expression of iNOS and NF-κB was inhibited in vitro. | [35,36] |
| Bacillus licheniformis | EPS 1-T14 | Stimulates Th1 cell-mediated immunity. | [23,24] |
| Thermus aquaticus | EPS TA-1 | Encourages the TLR2-dependent release of TNF-α and IL-6 in murine macrophages. | [24] |
| Serratia marcescens, Vibrio psychroerythrus, Pseudoalteromonas denitrificans & Zooshikella rubidus | Prodigiosin & Cycloprodigisin | Anti-inflammatory. Inhibits the activation of TNF-α induced NF-κB. | [27,28,37] |
| Bacillus sp. HC001, Piscicoccus sp. 12L081 | Diketopiperazines | Anti-inflammatory. Downregulates the release of TNF-α and IL-6 and suppress NF-κB expression. | [22] |
| Salinispora arenicola | Arenamides (Cyclic depsipeptide) | Blocks TNF-α in RAW 264.7 and human embryonic kidney cells. | [38,39] |
| Streptomyces sp. SCRC-A20 | Aburatubolactams | Antioxidant. Inhibits TPA-induced superoxide anion generation in human neutrophils. | [40] |
| Streptomyces sp. CNB-982 | Cyclomarins (Heptapeptides) | Anti-inflammatory. Inhibits oedema and pain in vivo. | [41] |
| Streptomyces sp. | Salinamides (Peptides) | Anti-inflammatory on phorbol ester-induced oedema mouse. | [42] |
| Streptomyces strain CNQ43 | Splenocin B | Anti-inflammatory. Potent inhibitors of pro-inflammatory cytokine IL-5, IL-13 and TNF-α. | [43] |
EPS, exopolysaccharide; iNOS, inducible nitric oxide synthase; IL, interleukin; NF-κB nuclear factor-κB; PKC, protein kinase C; TPA, 12-O-tetradecanoylphorbol-13-acetate; TLR, toll-like receptor; TNF, tumor necrosis factor.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ahmad, B.; Shah, M.; Choi, S. Oceans as a Source of Immunotherapy. Mar. Drugs 2019, 17, 282. https://doi.org/10.3390/md17050282
AMA Style
Ahmad B, Shah M, Choi S. Oceans as a Source of Immunotherapy. Marine Drugs. 2019; 17(5):282. https://doi.org/10.3390/md17050282
Chicago/Turabian StyleAhmad, Bilal, Masaud Shah, and Sangdun Choi. 2019. "Oceans as a Source of Immunotherapy" Marine Drugs 17, no. 5: 282. https://doi.org/10.3390/md17050282
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.