Oral Fucoidan Attenuates Lung Pathology and Clinical Signs in a Severe Influenza A Mouse Model

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
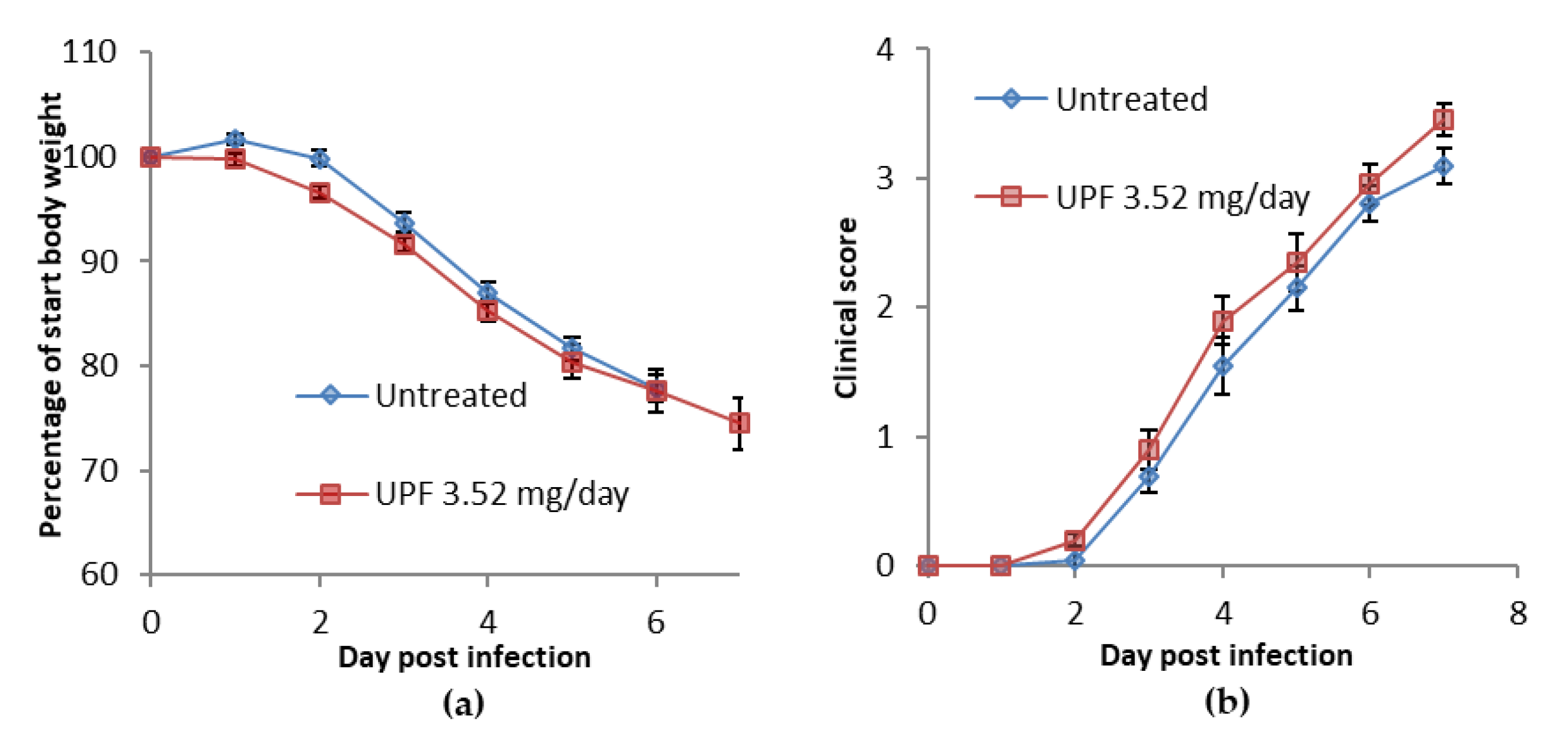
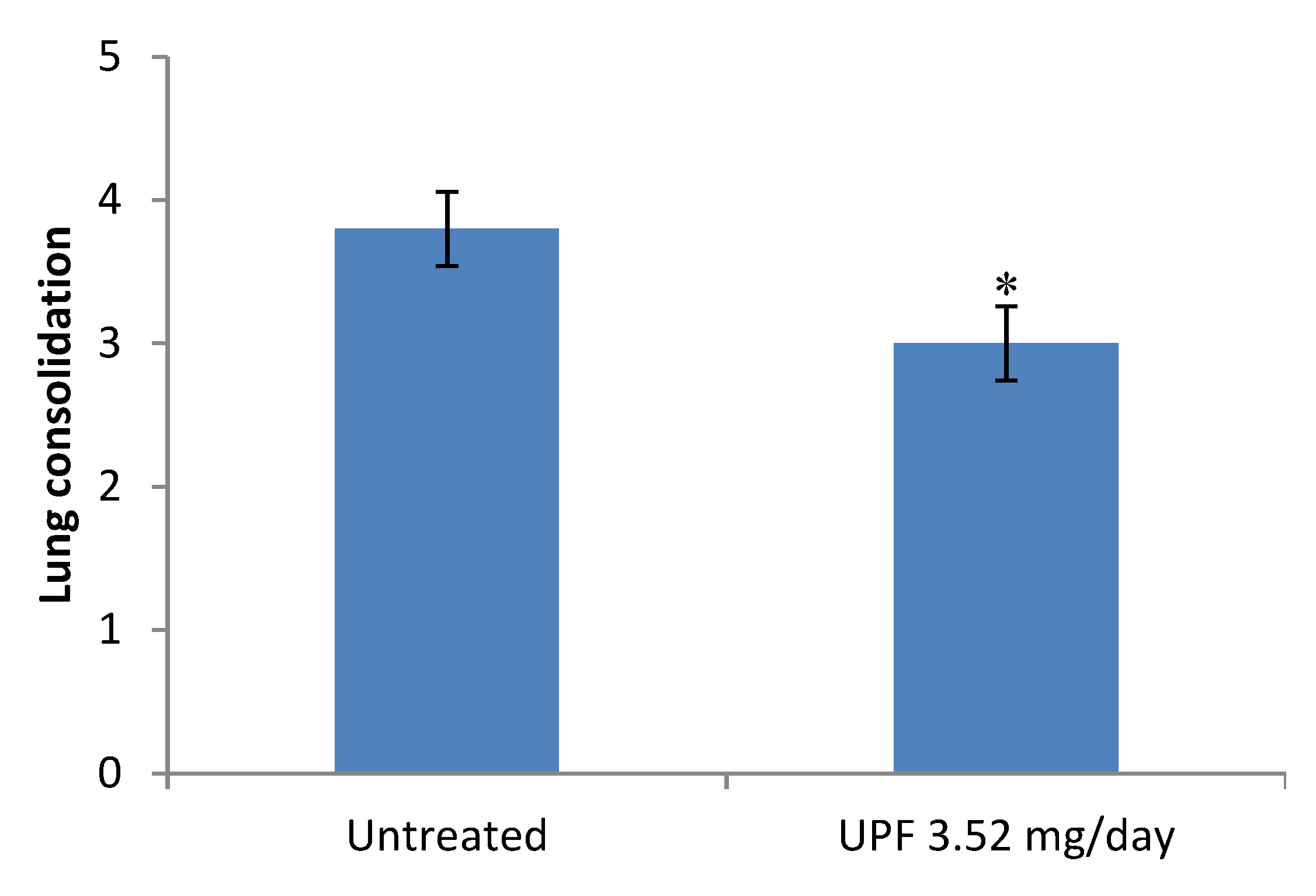
2.1. Treatment Model: Bodyweight, Clinical Disease Symptoms, and Lung Consolidation Scores
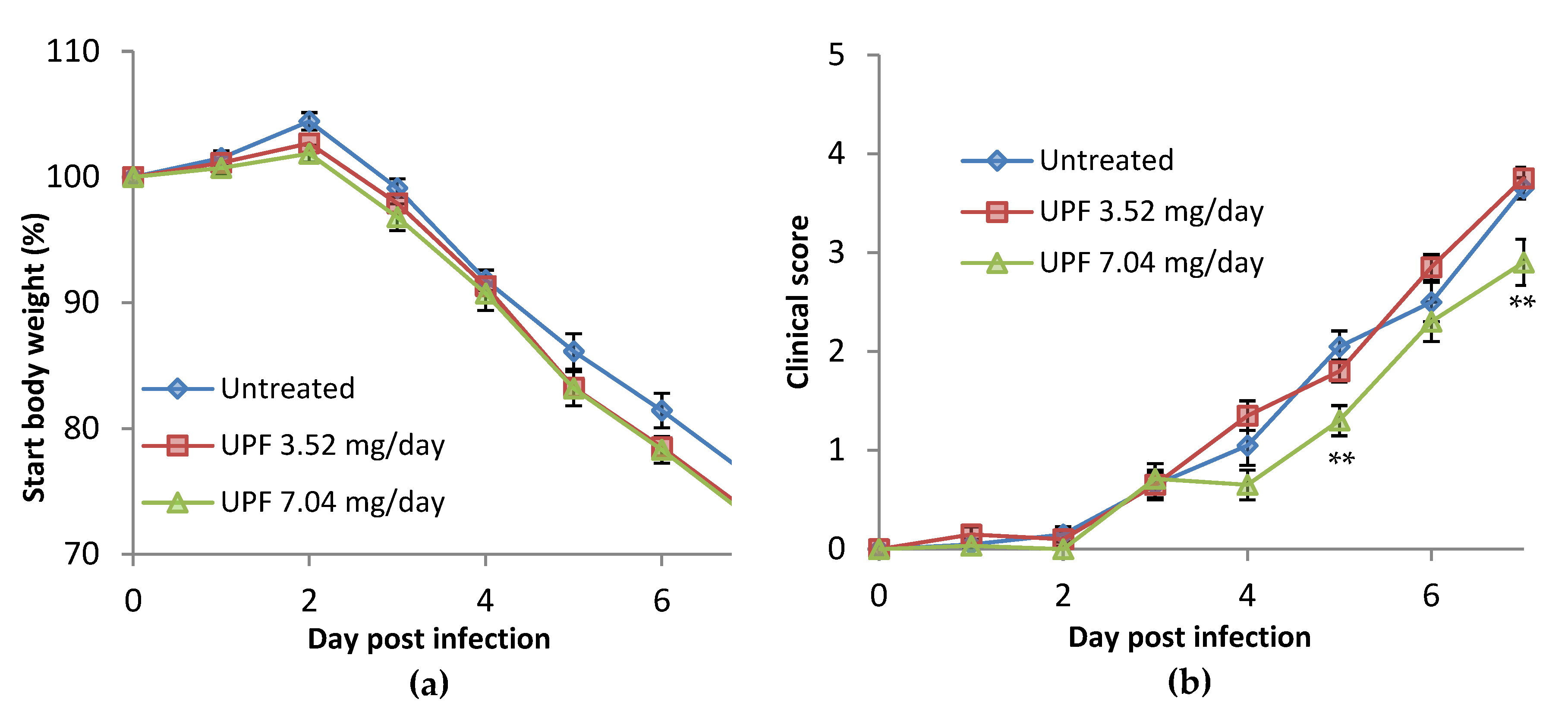
2.2. Prevention Model: Bodyweight
2.3. Prevention Model: Clinical Observations
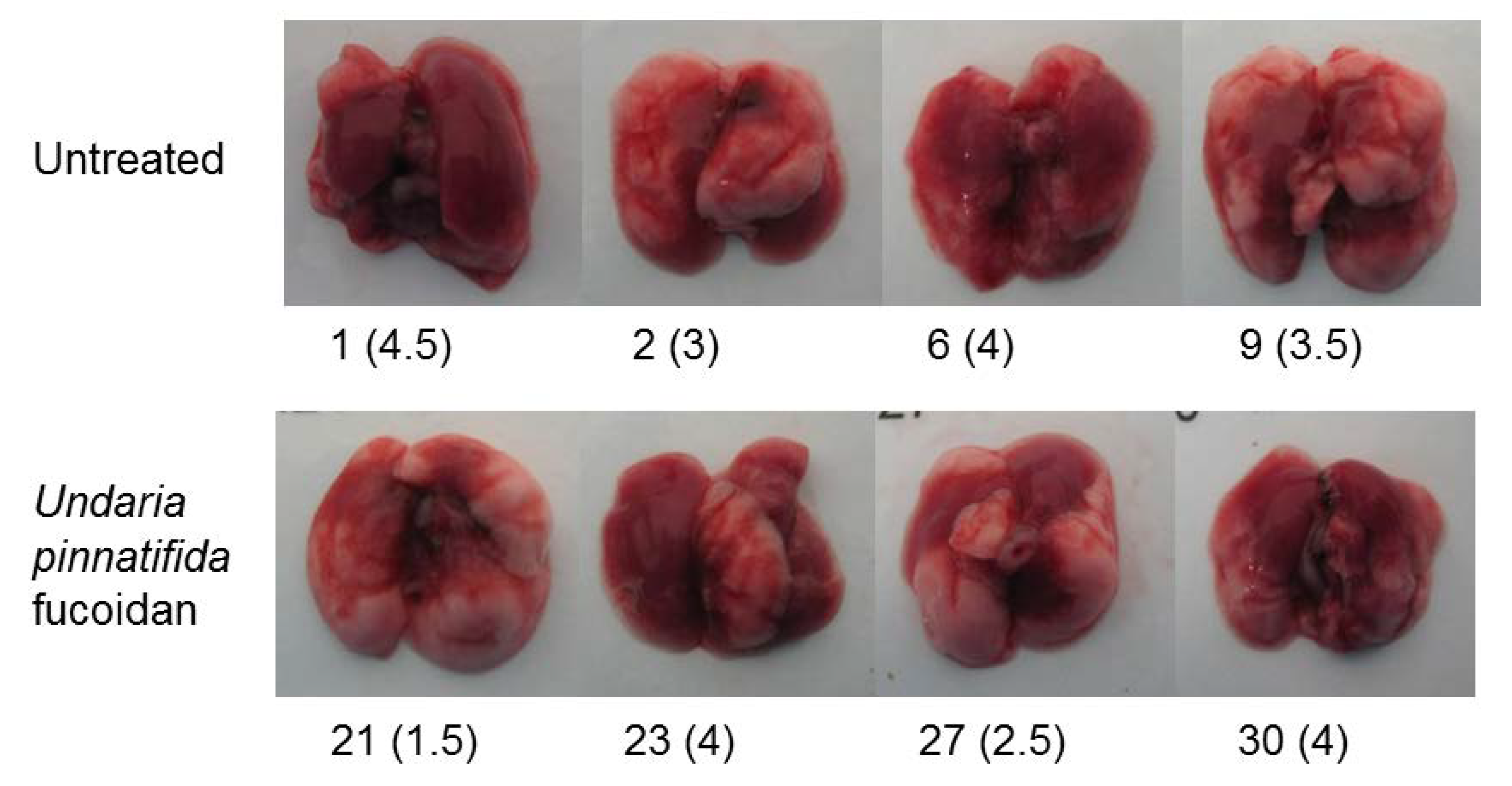
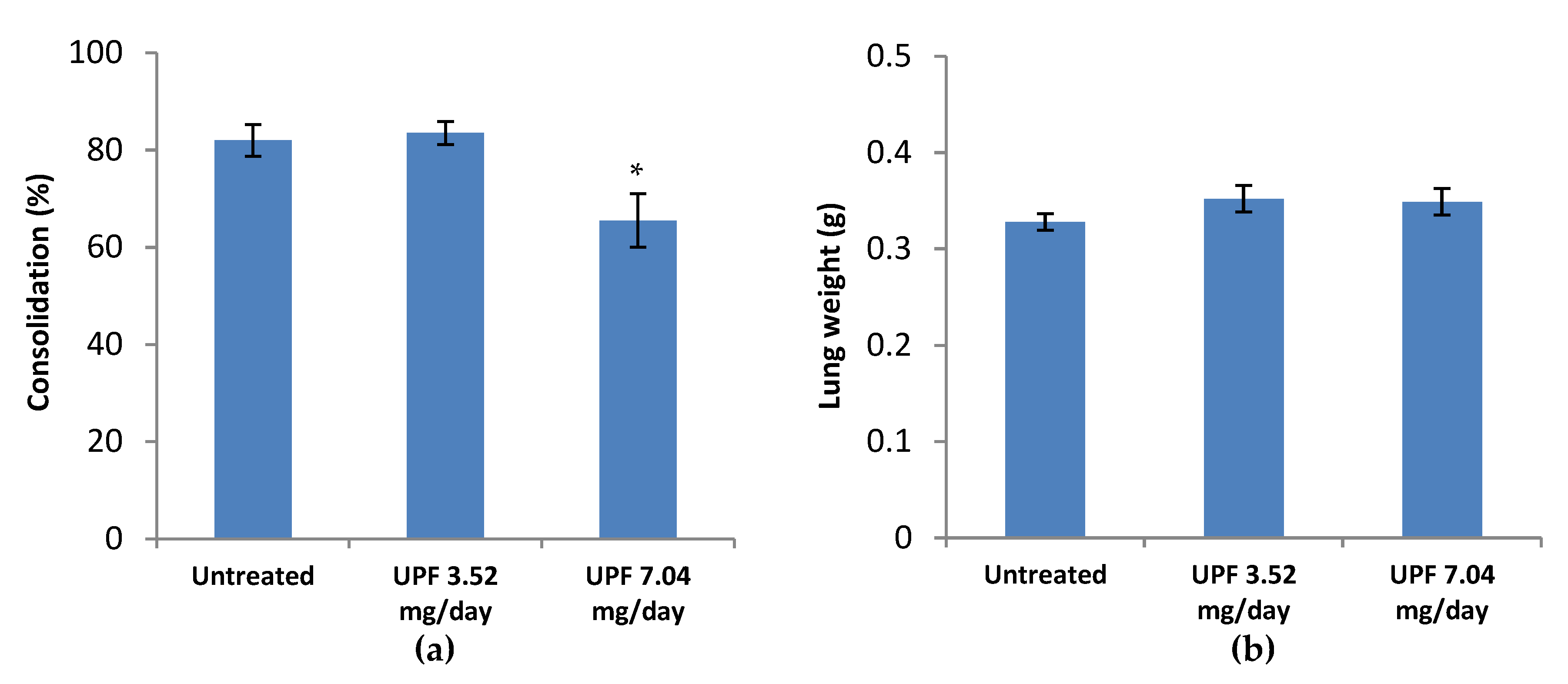
2.4. Prevention Model: Lung Consolidation
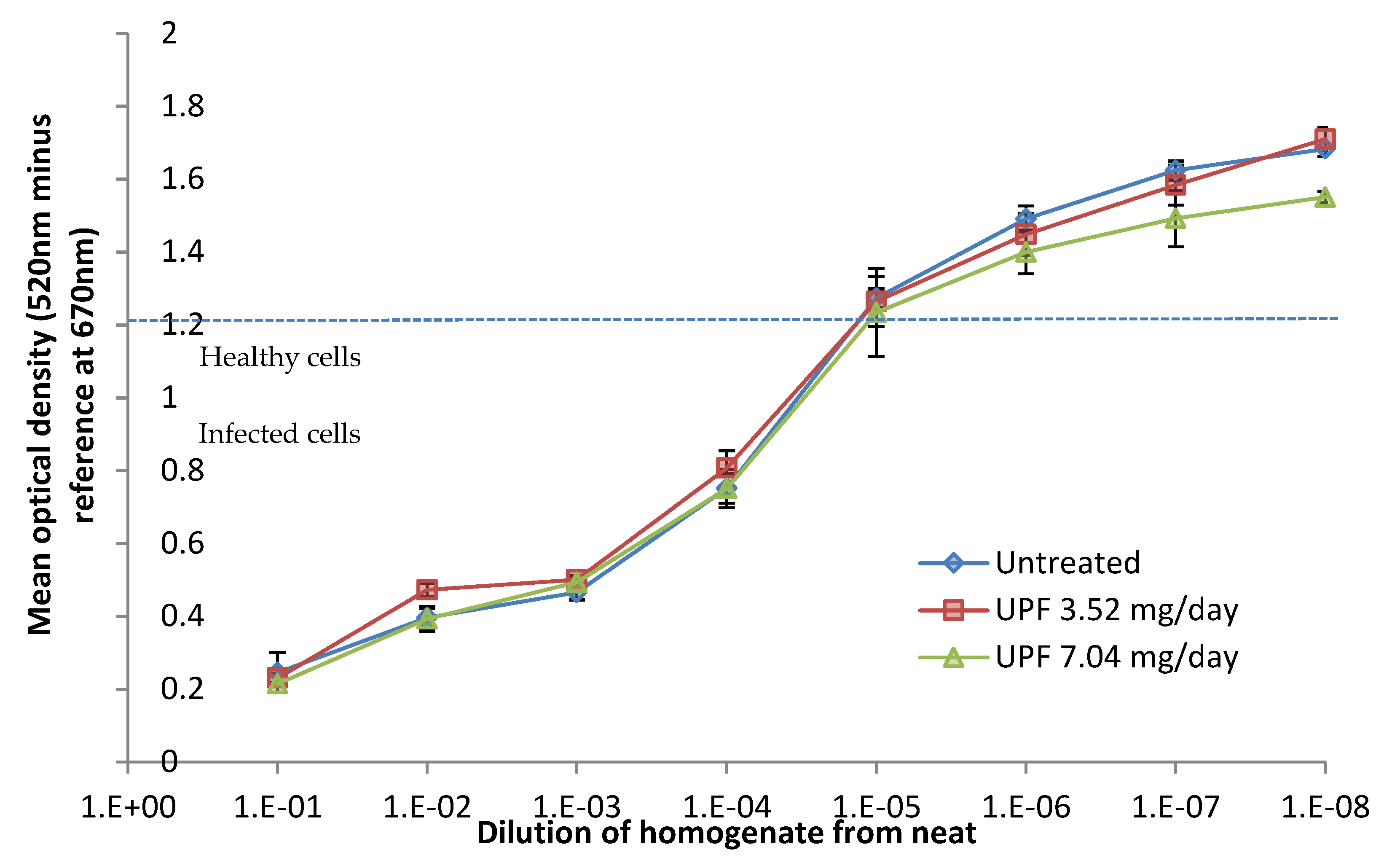
2.5. Prevention Model: Lung Titres
2.6. MTT Viability in the Treatment Model
3. Discussion
4. Experimental Section
4.1. Compounds and Reagents—Fucoidan from Undaria Pinnatifida
4.2. Delivery of Fucoidan
4.3. Assessment of Fucoidan from Undaria pinnatifida in a Treatment Model
4.4. Assessment of Fucoidan from Undaria Pinnatifida in a Prevention Model
4.5. Assessment of Clinical Symptoms
4.6. Lung Consolidation and Histopathology
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Baba, M.; Snoeck, R.; Pauwels, R.; de Clercq, E. Sulfated polysaccharides are potent and selective inhibitors of various enveloped viruses, including herpes simplex virus, cytomegalovirus, vesicular stomatitis virus, and human immunodeficiency virus. Antimicrob. Agents Chemother. 1988, 32, 1742–1745. [Google Scholar] [CrossRef] [Green Version]
- Makarenkova, I.D.; Deriabin, P.G.; L’Vov D, K.; Zviagintseva, T.N.; Besednova, N.N. Antiviral activity of sulfated polysaccharide from the brown algae Laminaria japonica against avian influenza A (H5N1) virus infection in the cultured cells. Voprosy Virusologii 2010, 55, 41–45. [Google Scholar] [PubMed]
- Taoda, N.; Shinji, E.; Nishii, K.; Nishioka, S.; Yonezawa, Y.; Uematsu, J.; Hattori, E.; Yamamoto, H.; Kawano, M.; Tsurudome, M.; et al. Fucoidan inhibits parainfluenza virus type 2 infection to LLCMK2 cells. Biomed. Res. 2008, 29, 331–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, T.; Hayashi, K.; Kanekiyo, K.; Ohta, Y.; Lee, J.-B. Promising antiviral Glyco-molecules from an edible alga. In Combating the Threat of Pandemic Infl uenza: Drug Discovery Approaches; Torrence, P.F., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2007; pp. 166–182. [Google Scholar]
- Synytsya, A.; Bleha, R.; Synytsya, A.; Pohl, R.; Hayashi, K.; Yoshinaga, K.; Nakano, T.; Hayashi, T. Mekabu fucoidan: Structural complexity and defensive effects against avian influenza A viruses. Carbohydr. Polym. 2014, 111, 633–644. [Google Scholar] [CrossRef] [PubMed]
- Leibbrandt, A.; Meier, C.; Konig-Schuster, M.; Weinmullner, R.; Kalthoff, D.; Pflugfelder, B.; Graf, P.; Frank-Gehrke, B.; Beer, M.; Fazekas, T.; et al. Iota-carrageenan is a potent inhibitor of influenza A virus infection. PLoS ONE 2010, 5, e14320. [Google Scholar] [CrossRef] [PubMed]
- Morokutti-Kurz, M.; Konig-Schuster, M.; Koller, C.; Graf, C.; Graf, P.; Kirchoff, N.; Reutterer, B.; Seifert, J.M.; Unger, H.; Grassauer, A.; et al. The Intranasal Application of Zanamivir and Carrageenan Is Synergistically Active against Influenza A Virus in the Murine Model. PLoS ONE 2015, 10, e0128794. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Yim, J.H.; Kim, S.Y.; Kim, H.S.; Lee, W.G.; Kim, S.J.; Kang, P.S.; Lee, C.K. In vitro inhibition of influenza a virus infection by marine microalga-derived sulfated polysaccharide p-KG03. Antivir. Res. 2012, 93, 253–259. [Google Scholar] [CrossRef]
- Gerber, P.; Dutcher, J.; Adams, E.; Sherman, J. Protective effect of seaweed extract for chicken embryos infected with influenza B or mumps virus. Proc. Soc. Exptl. Biol. Med. 1958, 99, 590–593. [Google Scholar] [CrossRef]
- Koenighofer, M.; Lion, T.; Bodenteich, A.; Prieschl-Grassauer, E.; Grassauer, A.; Unger, H.; Mueller, C.A.; Fazekas, T. Carrageenan nasal spray in virus confirmed common cold: Individual patient data analysis of two randomized controlled trials. Multidiscip. Respir. Med. 2014, 9, 57. [Google Scholar] [CrossRef] [Green Version]
- Thompson, K.D.; Dragar, C. Antiviral activity of Undaria pinnatifida against herpes simplex virus. Phytother. Res. 2004, 18, 551–555. [Google Scholar] [CrossRef]
- Hemmingson, J.; Falshaw, R.; Furneaux, R.; Thompson, K. Structure and Antiviral Activity of the Galactofucan Sulfates Extracted from Undaria Pinnatifida (Phaeophyta). J. Appl. Phycol. 2006, 18, 185–193. [Google Scholar] [CrossRef]
- Hayashi, K.; Nakano, T.; Hashimoto, M.; Kanekiyo, K.; Hayashi, T. Defensive effects of a fucoidan from brown alga Undaria pinnatifida against herpes simplex virus infection. Int. Immunopharmacol. 2008, 8, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-B.; Hayashi, K.; Hashimoto, M.; Nakano, T.; Hayashi, T. Novel antiviral fucoidan from sporophyll of Undaria pinnatifida (Mekabu). Chem. Pharm. Bull. 2004, 52, 1091–1094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krylova, N.V.; Ermakova, S.P.; Lavrov, V.F.; Leneva, I.A.; Kompanets, G.G.; Iunikhina, O.V.; Nosik, M.N.; Ebralidze, L.K.; Falynskova, I.N.; Silchenko, A.S.; et al. The Comparative Analysis of Antiviral Activity of Native and Modified Fucoidans from Brown Algae Fucus evanescens In Vitro and In Vivo. Mar. Drugs 2020, 18, 224. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Wu, J.; Zhang, X.; Hao, C.; Zhao, X.; Jiao, G.; Shan, X.; Tai, W.; Yu, G. Inhibition of Influenza A Virus Infection by Fucoidan Targeting Viral Neuraminidase and Cellular EGFR Pathway. Sci. Rep. 2017, 7, 40760. [Google Scholar] [CrossRef]
- Negishi, H.; Mori, M.; Mori, H.; Yamori, Y. Supplementation of elderly Japanese men and women with fucoidan from seaweed increases immune responses to seasonal influenza vaccination. J. Nutr. 2013, 143, 1794–1798. [Google Scholar] [CrossRef]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [Green Version]
- Openshaw, P.J. Crossing barriers: Infections of the lung and the gut. Mucosal Immunol. 2009, 2, 100–102. [Google Scholar] [CrossRef] [Green Version]
- Shu, Y.; Li, C.K.; Li, Z.; Gao, R.; Liang, Q.; Zhang, Y.; Dong, L.; Zhou, J.; Dong, J.; Wang, D.; et al. Avian influenza A(H5N1) viruses can directly infect and replicate in human gut tissues. J. Infect. Dis. 2010, 201, 1173–1177. [Google Scholar] [CrossRef]
- Fitton, J. Therapies from fucoidan; multifunctional marine polymers. Mar. Drugs 2011, 9, 1731–1760. [Google Scholar] [CrossRef]
- Pozharitskaya, O.N.; Shikov, A.N.; Faustova, N.M.; Obluchinskaya, E.D.; Kosman, V.M.; Vuorela, H.; Makarov, V.G. Pharmacokinetic and Tissue Distribution of Fucoidan from Fucus vesiculosus after Oral Administration to Rats. Mar. Drugs 2018, 16, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokita, Y.; Hirayama, M.; Nakajima, K.; Tamaki, K.; Iha, M.; Nagamine, T. Detection of Fucoidan in Urine after Oral Intake of Traditional Japanese Seaweed, Okinawa mozuku (Cladosiphon okamuranus Tokida). J. Nutr. Sci. Vitaminol. 2017, 63, 419–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitton, J.H.; Dell’Acqua, G.; Gardiner, V.-A.; Karpiniec, S.S.; Stringer, D.N.; Davis, E. Topical benefits of two fucoidan-rich extracts from marine macroalgae. Cosmetics 2015, 2, 66–81. [Google Scholar] [CrossRef] [Green Version]
- Morvai-Vitányi, M.; Molnár-Perl, I.; Knausz, D.; Sass, P. Simultaneous GC derivatization and quantification of acids and sugars. Chromatographia 1993, 36, 204–206. [Google Scholar] [CrossRef]
- Filisetti-Cozzi, T.M.C.C.; Carpita, N.C. Measurement of uronic acids without interference from neutral sugars. Anal. Biochem. 1991, 197, 157–162. [Google Scholar] [CrossRef]
- Dodgson, K.S.; Price, R.G. A note on the determination of the ester sulphate content of sulphated polysaccharides. Biochem. J. 1962, 84, 106–110. [Google Scholar] [CrossRef] [Green Version]
- Dogson, K.S. Determination of inorganic sulphate in studies on the enzymic and non-enzymic hydrolysis of carbohydrate and other sulphate esters. Biochem. J. 1961, 78, 312–319. [Google Scholar]
- Schouten, M.; Sluijs, K.F.; Gerlitz, B.; Grinnell, B.W.; Roelofs, J.J.; Levi, M.M.; van’t Veer, C.; van der Poll, T. Activated protein C ameliorates coagulopathy but does not influence outcome in lethal H1N1 influenza: A controlled laboratory study. Crit. Care 2010, 14, R65. [Google Scholar] [CrossRef] [Green Version]
- Longhi, M.P.; Williams, A.; Wise, M.; Morgan, B.P.; Gallimore, A. CD59a deficiency exacerbates influenza-induced lung inflammation through complement-dependent and -independent mechanisms. Eur. J. Immunol. 2007, 37, 1266–1274. [Google Scholar] [CrossRef] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Richards, C.; Williams, N.A.; Fitton, J.H.; Stringer, D.N.; Karpiniec, S.S.; Park, A.Y. Oral Fucoidan Attenuates Lung Pathology and Clinical Signs in a Severe Influenza A Mouse Model. Mar. Drugs 2020, 18, 246. https://doi.org/10.3390/md18050246
Richards C, Williams NA, Fitton JH, Stringer DN, Karpiniec SS, Park AY. Oral Fucoidan Attenuates Lung Pathology and Clinical Signs in a Severe Influenza A Mouse Model. Marine Drugs. 2020; 18(5):246. https://doi.org/10.3390/md18050246
Chicago/Turabian StyleRichards, Claire, Neil A. Williams, J. Helen Fitton, Damien N. Stringer, Samuel S. Karpiniec, and Ah Young Park. 2020. "Oral Fucoidan Attenuates Lung Pathology and Clinical Signs in a Severe Influenza A Mouse Model" Marine Drugs 18, no. 5: 246. https://doi.org/10.3390/md18050246