Effects of a Newly Developed Enzyme-Assisted Extraction Method on the Biological Activities of Fucoidans in Ocular Cells


 and
and
Abstract
:1. Introduction
2. Results
2.1. Chemical Characterization of the Fucoidans
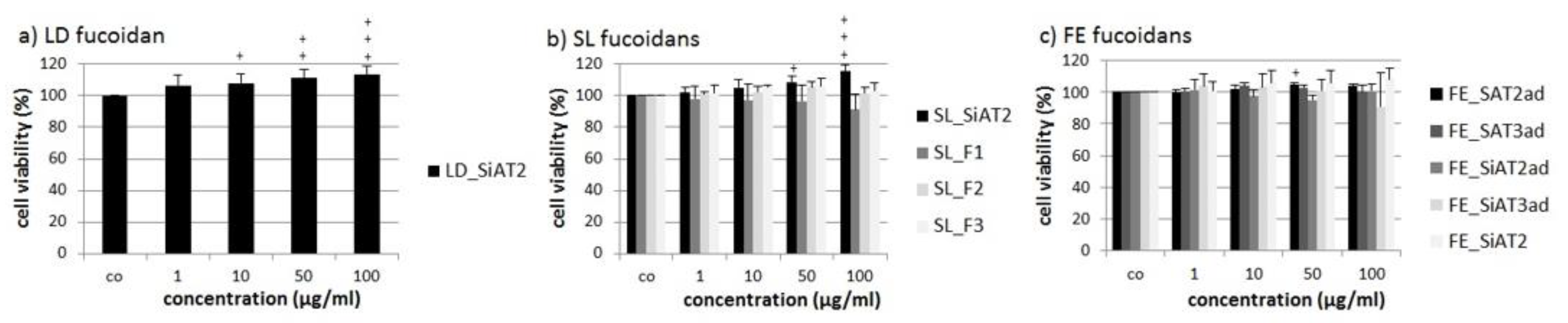
2.2. Effects on Cell Viability
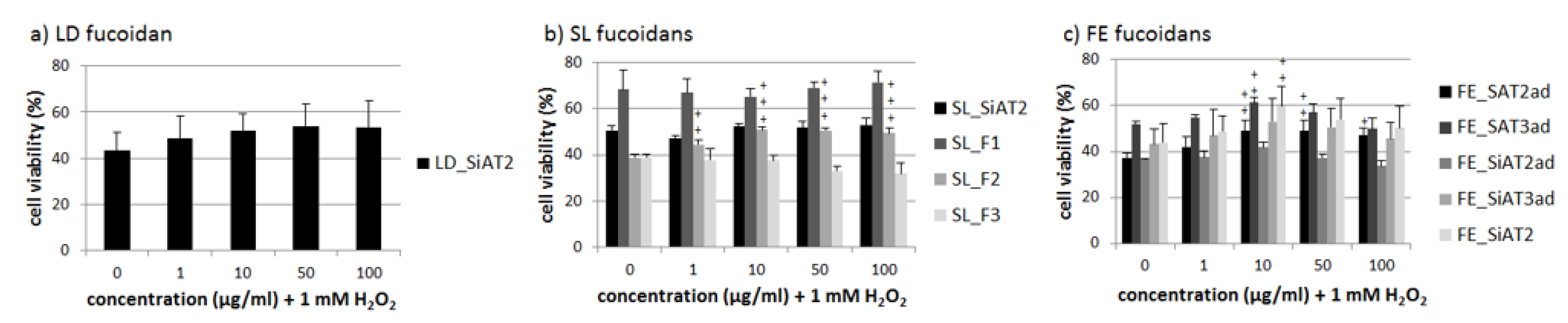
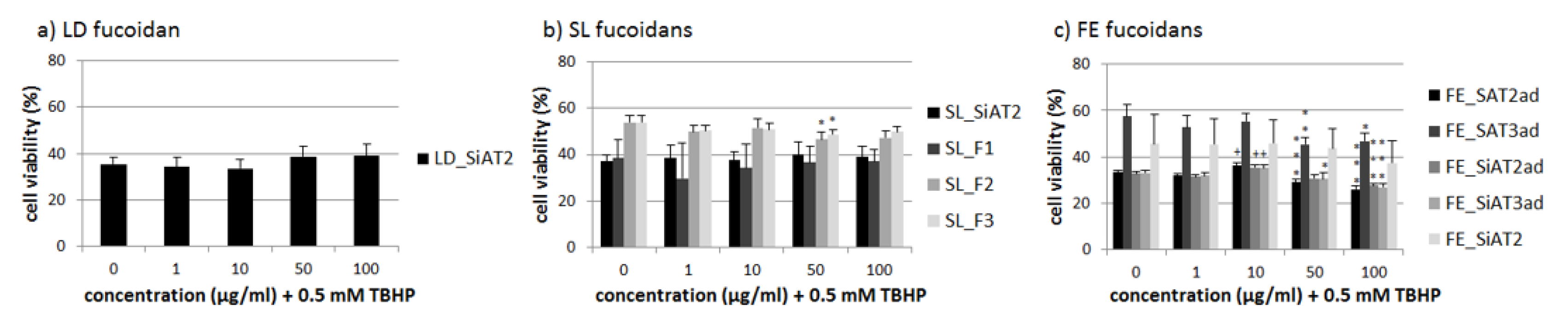
2.3. Effects on Oxidative Stress Protection
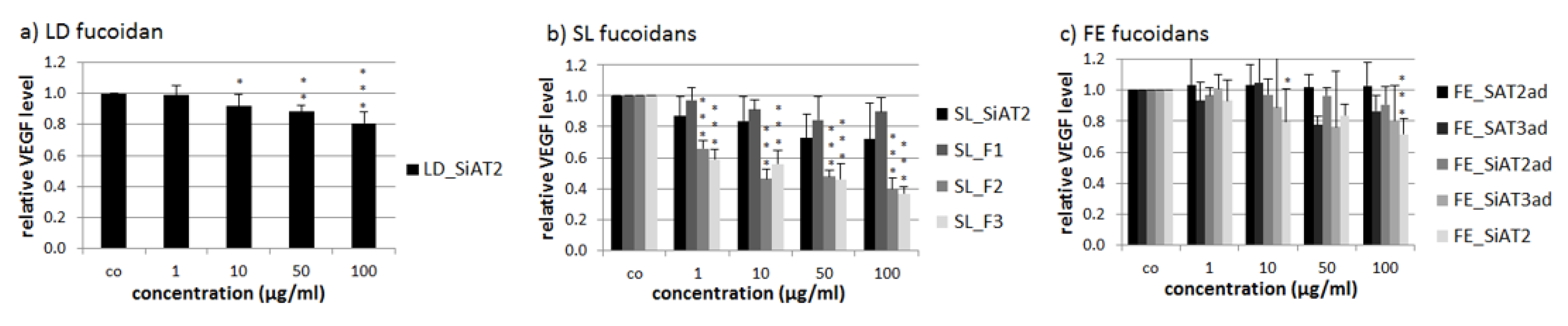
2.4. VEGF Secretion of ARPE-19
3. Discussion
3.1. Integration in Previous Studies
3.2. Slightly Increased Cell Viability in OMM-1 and ARPE-19 Relation to Uronic Acids, Molecular Weight and Concentration
3.3. Effects on Oxidative Stress Protection-Heterogeneous Results, Dependency on Alginates and Galactose
3.4. Effects on VEGF-Acid Precipitation Lowers and Higher Molecular Weight Improves VEGF Inhibition
3.5. Comparison of Cellic®CTec2 and 3, Alginate Lyases as Well as Precipitation Technique
3.6. Different Fucoidan Structures between Algal Species Lead to the Described Biological Effects
3.7. Conclusive Words
4. Material and Methods
4.1. Cell Culture
4.2. Used Fucoidans, Extraction and Purification Process
4.2.1. Fucoidan Origin
4.2.2. Alginate Lyase Expression and Purification
4.2.3. Enzyme Assisted Extraction of Brown Seaweed Polysaccharides
4.2.4. SL Fucoidan Fractionation by Anion-Exchange Chromatography
4.3. Chemical Composition and Size Distribution Analysis
4.4. Oxidative Stress
4.4.1. OMM-1
4.4.2. ARPE-19
4.5. Methyl Thiazolyl Tetrazolium (MTT)-Assay
4.6. MTS-Assay
4.7. VEGF ELISA
4.8. Statistics
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Fuc | Mannitol | Ara/Rham | Gal | Glc | Xyl | Man | GuluA | GluA | ManA | Total | Sulfate (SO42−) mol% |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LD_SiAT2 | 3.9 ± 0.1 | 1.2 ± 0.1 | 0.1 ± 0.0 | 14 ± 0.2 | 2.3 ± 0.1 | 1.5 ± 0.1 | 1.2 ± 0.1 | 12.4 ± 1.4 | 1.2 ± 0.1 | 74.8 ± 1.5 | 100 | 9.3 ± 2.4 |
| SL_SiAT2 | 12.3 ± 0.8 | 0.2 ± 0.0 | 0.2 ± 0.0 | 1.3 ± 1.1 | 2.6 ± 0.2 | 0.9 ± 0.0 | 0.8 ± 0.0 | 32.2 ± 1.2 | 1.5 ± 0.2 | 48.1 ± 0.7 | 100 | 14.4 ± 0.6 |
| FE_SiAT2 | 15.5 ± 0.9 | 0.0 ± 0.0 | 0.3 ± 0.0 | 2.5 ± 0.0 | 2.1 ± 0.0 | 3.6 ± 0.1 | 1.8 ± 0.2 | 18.7 ± 0.8 | 6.5 ± 5.5 | 48.8 ± 3.5 | 100 | 20.2 ± 1.5 |
| FE_SiAT2ad | 36.1 ± 3.1 | 0.1 ± 0.1 | 0.6 ± 0.6 | 6.6 ± 1.9 | 5.7 ± 1.5 | 10.2 ± 3.2 | 1.8 ± 1.5 | 7.1 ± 0.7 | 2.0 ± 0.4 | 30.0 ± 2.0 | 100 | 30.1 ± 0.6 |
| FE_SiAT3ad | 35.9 ± 1.2 | 0.1 ± 0.0 | 0.4 ± 0.3 | 2.8 ± 0.7 | 2.2 ± 0.1 | 4.1 ± 0.3 | 2.0 ± 0.1 | 10. 0 ± 0.2 | 2.2 ± 0.0 | 40.4 ± 0.5 | 100 | 29.4 ± 1.7 |
| FE_SAT2ad | 52.2 ± 1.9 | 0.0 ± 0.0 | 1.2 ± 0.2 | 4.8 ± 0.5 | 2.7 ± 0.5 | 9.3 ± 1.1 | 4.7 ± 0.7 | 8.8 ± 1.2 | 3.9 ± 0.3 | 12.5 ± 0.8 | 100 | 31.7 ± 2.0 |
| FE_SAT3ad | 48.3 ± 0.6 | 0.1 ± 0.1 | 0.7 ± 0.6 | 5.0 ± 0.7 | 2.0 ± 0.1 | 8.3 ± 0.5 | 4.0 ± 0.7 | 11.6 ± 0.1 | 3.7 ± 0.2 | 16.4 ± 0.3 | 100 | 29.9 ± 1.4 |
| SL_F1 * | 5.4 ± 1.2 | 0.0 ± 0.0 | 0.1 ± 0.0 | 0.5 ± 0.0 | 0.4 ± 0.0 | 0.8 ± 0.1 | 0.8 ± 0.1 | 8.5 ± 4.7 | 1.1 ± 0.1 | 82.4 ± 4.3 | 100 | 6.6 ± 3.6 |
| SL_F2 * | 64.7 ± 0.3 | 0.1 ± 0.0 | 0.3 ± 0.0 | 12.2 ± 0.1 | 0.6 ± 0.1 | 4.8 ± 0.0 | 3.5 ± 0.2 | 0.0 ± 0.0 | 6.9 ± 0.3 | 6.9 ± 0.1 | 100 | 35.6 ± 2.5 |
| SL_F3 * | 63.3 ± 0.7 | 0.0 ± 0.0 | 0.3 ± 0.0 | 26.9 ± 0.3 | 0.4 ± 0.1 | 3.4 ± 0.2 | 2.1 ± 0.1 | 0.0 ± 0.0 | 2.8 ± 0.2 | 0.8 ± 0.1 | 100 | 46.4 ± 3.5 |
References
- Wong, W.L.; Su, X.; Li, X.; Cheung, C.M.G.; Klein, R.; Cheng, C.-Y.; Wong, T.Y. Global prevalence of age-related macular degeneration and disease burden projection for 2020 and 2040: A systematic review and meta-analysis. Lancet Glob. Health 2014, 2, e106–e116. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.W. Age-related macular degeneration revisited--piecing the puzzle: The LXIX Edward Jackson memorial lecture. Am. J. Ophthalmol. 2013, 155, 1–35.e13. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Patel, M.; Chan, C.-C. Molecular pathology of age-related macular degeneration. Prog. Retin. Eye Res. 2009, 28, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klettner, A. Oxidative stress induced cellular signaling in RPE cells. Front. Biosci. 2012, 4, 392–411. [Google Scholar] [CrossRef]
- Hageman, G.S.; Anderson, D.H.; Johnson, L.V.; Hancox, L.S.; Taiber, A.J.; Hardisty, L.I.; Hageman, J.L.; Stockman, H.A.; Borchardt, J.D.; Gehrs, K.M.; et al. A common haplotype in the complement regulatory gene factor H (HF1/CFH) predisposes individuals to age-related macular degeneration. Proc. Natl. Acad. Sci. USA 2005, 102, 7227–7232. [Google Scholar] [CrossRef] [Green Version]
- McHarg, S.; Clark, S.J.; Day, A.J.; Bishop, P.N. Age-related macular degeneration and the role of the complement system. Mol. Immunol. 2015, 67, 43–50. [Google Scholar] [CrossRef]
- Schmidt-Erfurth, U.; Chong, V.; Loewenstein, A.; Larsen, M.; Souied, E.; Schlingemann, R.; Eldem, B.; Monés, J.; Richard, G.; Bandello, F. Guidelines for the management of neovascular age-related macular degeneration by the European Society of Retina Specialists (EURETINA). Br. J. Ophthalmol. 2014, 98, 1144–1167. [Google Scholar] [CrossRef]
- Deniaud-Bouët, E.; Kervarec, N.; Michel, G.; Tonon, T.; Kloareg, B.; Hervé, C. Chemical and enzymatic fractionation of cell walls from Fucales: Insights into the structure of the extracellular matrix of brown algae. Ann. Bot. 2014, 114, 1203–1216. [Google Scholar] [CrossRef] [Green Version]
- Catarino, M.D.; Silva, A.M.S.; Cardoso, S.M. Phycochemical Constituents and Biological Activities of Fucus spp. Mar. Drugs 2018, 16, 249. [Google Scholar] [CrossRef] [Green Version]
- Ale, M.T.; Meyer, A.S. Fucoidans from brown seaweeds: An update on structures, extraction techniques and use of enzymes as tools for structural elucidation. RSC Adv. 2013, 3, 8131–8141. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Lu, F.; Wei, X.; Zhao, R. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klettner, A. Fucoidan as a Potential Therapeutic for Major Blinding Diseases--A Hypothesis. Mar. Drugs 2016, 14, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dithmer, M.; Fuchs, S.; Shi, Y.; Schmidt, H.; Richert, E.; Roider, J.; Klettner, A. Fucoidan reduces secretion and expression of vascular endothelial growth factor in the retinal pigment epithelium and reduces angiogenesis in vitro. PLoS ONE 2014, 9, e89150. [Google Scholar] [CrossRef] [PubMed]
- Ale, M.T.; Mikkelsen, J.D.; Meyer, A.S. Important determinants for fucoidan bioactivity: A critical review of structure-function relations and extraction methods for fucose-containing sulfated polysaccharides from brown seaweeds. Mar. Drugs 2011, 9, 2106–2130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, T.T.; Mikkelsen, M.D.; Ha, V.T.N.; Dieu, T.V.T.; Rhein-Knudsen, N.; Holck, J.; Rasin, A.B.; Cao, H.T.T.; Van, T.T.T.; Meyer, A.S. Enzyme assisted fucoidan extraction from the brown macroalgae. Mar. Drugs 2020, 18, 168. [Google Scholar]
- Bittkau, K.S.; Dörschmann, P.; Blümel, M.; Tasdemir, D.; Roider, J.; Klettner, A.; Alban, S. Comparison of the Effects of Fucoidans on the Cell Viability of Tumor and Non-Tumor Cell Lines. Mar. Drugs 2019, 17, 441. [Google Scholar] [CrossRef] [Green Version]
- Dörschmann, P.; Kopplin, G.; Roider, J.; Klettner, A. Effects of Sulfated Fucans from Laminaria hyperborea Regarding VEGF Secretion, Cell Viability, and Oxidative Stress and Correlation with Molecular Weight. Mar. Drugs 2019, 17, 548. [Google Scholar] [CrossRef] [Green Version]
- Rohwer, K.; Neupane, S.; Bittkau, K.S.; Galarza Pérez, M.; Dörschmann, P.; Roider, J.; Alban, S.; Klettner, A. Effects of Crude Fucus distichus Subspecies evanescens Fucoidan Extract on Retinal Pigment Epithelium Cells-Implications for Use in Age-Related Macular Degeneration. Mar. Drugs 2019, 17, 538. [Google Scholar] [CrossRef] [Green Version]
- Dörschmann, P.; Bittkau, K.S.; Neupane, S.; Roider, J.; Alban, S.; Klettner, A. Effects of fucoidans from five different brown algae on oxidative stress and VEGF interference in ocular cells. Mar. Drugs 2019, 17, 258. [Google Scholar] [CrossRef] [Green Version]
- Schneider, T.; Ehrig, K.; Liewert, I.; Alban, S. Interference with the CXCL12/CXCR4 axis as potential antitumor strategy: Superiority of a sulfated galactofucan from the brown alga Saccharina latissima and fucoidan over heparins. Glycobiology 2015, 25, 812–824. [Google Scholar] [CrossRef] [Green Version]
- Bittkau, K.S.; Neupane, S.; Alban, S. Initial evaluation of six different brown algae species as source for crude bioactive fucoidans. Algal Res. 2020, 45, 101759. [Google Scholar] [CrossRef]
- Manns, D.; Deutschle, A.L.; Saake, B.; Meyer, A.S. Methodology for quantitative determination of the carbohydrate composition of brown seaweeds (Laminariaceae). RSC Adv. 2014, 4, 25736–25746. [Google Scholar] [CrossRef] [Green Version]
- Manns, D.; Andersen, S.K.; Saake, B.; Meyer, A.S. Brown seaweed processing: Enzymatic saccharification of Laminaria digitata requires no pre-treatment. J. Appl. Phycol. 2016, 28, 1287–1294. [Google Scholar] [CrossRef] [Green Version]
- Manns, D.; Nyffenegger, C.; Saake, B.; Meyer, A.S. Impact of different alginate lyases on combined cellulase–lyase saccharification of brown seaweed. RSC Adv. 2016, 6, 45392–45401. [Google Scholar] [CrossRef] [Green Version]
- Alboofetileh, M.; Rezaei, M.; Tabarsa, M. Enzyme-assisted extraction of Nizamuddinia zanardinii for the recovery of sulfated polysaccharides with anticancer and immune-enhancing activities. J. Appl. Phycol. 2019, 31, 1391–1402. [Google Scholar] [CrossRef]
- Karlsson, M.; Kurz, T. Attenuation of iron-binding proteins in ARPE-19 cells reduces their resistance to oxidative stress. Acta Ophthalmol. 2016, 94, 556–564. [Google Scholar] [CrossRef]
- Mak, W.; Wang, S.K.; Liu, T.; Hamid, N.; Li, Y.; Lu, J.; White, W.L. Anti-Proliferation Potential and Content of Fucoidan Extracted from Sporophyll of New Zealand Undaria pinnatifida. Front. Nutr. 2014, 1, 9. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhao, H.; Wang, Q.; Liang, H.; Jiang, X. Fucoidan Protects Arpe-19 Cells from Oxidative Stress via Normalization of Reactive Oxygen Species Generation Through the Ca²⁺-Dependent Erk Signaling Pathway. Mol. Med. Rep. 2015, 11, 3746–3752. [Google Scholar] [CrossRef] [Green Version]
- Ehrig, K.; Alban, S. Sulfated galactofucan from the brown alga Saccharina latissima--variability of yield, structural composition and bioactivity. Mar. Drugs 2014, 13, 76–101. [Google Scholar] [CrossRef] [Green Version]
- Bo-Htay, C.; Palee, S.; Apaijai, N.; Chattipakorn, S.C.; Chattipakorn, N. Effects of d-galactose-induced ageing on the heart and its potential interventions. J. Cell. Mol. Med. 2018, 22, 1392–1410. [Google Scholar] [CrossRef] [Green Version]
- Van Weelden, G.; Bobiński, M.; Okła, K.; van Weelden, W.J.; Romano, A.; Pijnenborg, J.M.A. Fucoidan Structure and Activity in Relation to Anti-Cancer Mechanisms. Mar. Drugs. 2019, 17, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilan, M.I.; Grachev, A.A.; Ustuzhanina, N.E.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. Structure of a fucoidan from the brown seaweed Fucus evanescens C.Ag. Carbohydr. Res. 2002, 337, 719–730. [Google Scholar] [CrossRef]
- Zvyagintseva, T.N.; Shevchenko, N.M.; Chizhov, A.O.; Krupnova, T.N.; Sundukova, E.V.; Isakov, V.V. Water-soluble polysaccharides of some far-eastern brown seaweeds. Distribution, structure, and their dependence on the developmental conditions. J. Exp. Mar. Biol. Ecol. 2003, 294, 1–13. [Google Scholar] [CrossRef]
- Menshova, R.V.; Shevchenko, N.M.; Imbs, T.I.; Zvyagintseva, T.N.; Malyarenko, O.S.; Zaporoshets, T.S.; Besednova, N.N.; Ermakova, S.P. Fucoidans from Brown Alga Fucus evanescens: Structure and Biological Activity. Front. Mar. Sci. 2016, 3, 7. [Google Scholar] [CrossRef] [Green Version]
- Bilan, M.I.; Grachev, A.A.; Shashkov, A.S.; Kelly, M.; Sanderson, C.J.; Nifantiev, N.E.; Usov, A.I. Further studies on the composition and structure of a fucoidan preparation from the brown alga Saccharina latissima. Carbohydr. Res. 2010, 345, 2038–2047. [Google Scholar] [CrossRef] [PubMed]
- Luyten, G.P.; Naus, N.C.; Mooy, C.M.; Hagemeijer, A.; Kan-Mitchell, J.; van Drunen, E.; Vuzevski, V.; de Jong, P.T.; Luider, T.M. Establishment and characterization of primary and metastatic uveal melanoma cell lines. Int. J. Cancer 1996, 66, 380–387. [Google Scholar] [CrossRef]
- Dunn, K.C.; Aotaki-Keen, A.E.; Putkey, F.R.; Hjelmeland, L.M. ARPE-19, a human retinal pigment epithelial cell line with differentiated properties. Exp. Eye Res. 1996, 62, 155–169. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Dithmer, M.; Kirsch, A.-M.; Richert, E.; Fuchs, S.; Wang, F.; Schmidt, H.; Coupland, S.E.; Roider, J.; Klettner, A. Fucoidan Does Not Exert Anti-Tumorigenic Effects on Uveal Melanoma Cell Lines. Mar. Drugs. 2017, 15, 193. [Google Scholar] [CrossRef]
- Riss, T.L.; Moravec, R.A.; Niles, A.L.; Duellman, S.; Benink, H.A.; Worzella, T.J.; Minor, L. Cell Viability Assays. In Assay Guidance Manual [Internet]; Sittampalam, G.S., Coussens, N.P., Brimacombe, K., Grossman, A., Arkin, M., Auld, D., Austin, C., Baell, J., Bejcek, B., Caaveiro, J.M.M., et al., Eds.; Eli Lilly & Company and the National Center for Advancing Translational Sciences: Indianapolis, IN, USA, 2016. [Google Scholar]





| Fucoidan Code | Extraction Method | Algal Species |
|---|---|---|
| LD_SiAT2 | SigmALY_CTECH2_crude | Laminaria digitata |
| SL_SiAT2 | SigmAly_CTECH2_crude | Saccharina latissima |
| FE_SiAT2 | SigmAly_CTECH2_crude | Fucus distichus subsp. evanescens |
| FE_SiAT2ad | SigmAly_CTECH2_acid_dialysis | Fucus distichus subsp. evanescens |
| FE_SiAT3ad | SigmAly_CTECH3_acid_dialysis | Fucus distichus subsp. evanescens |
| FE_SAT2ad | Saly_CTECH2_acid_dialysis | Fucus distichus subsp. evanescens |
| FE_SAT3ad | Saly_CTECH3_acid_dialysis | Fucus distichus subsp. evanescens |
| SL_F1 | SALy_CTECH2_CaCl2_IEX_filtering_Fraction 1 * | Saccharina latissima |
| SL_F2 | SALy_CTECH2_CaCl2_IEX_filtering_Fraction 2 * | Saccharina latissima |
| SL_F3 | SALy_CTECH2_CaCl2_IEX_filtering_Fraction 3 * | Saccharina latissima |
| Sample | Fuc | GuluA | ManA | GuluA + ManA | Mannitol | Ara/Rham | Gal | Glc | Xyl | Man | GluA | Total | Sulfate (SO42−), % |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LD_SiAT2 | 3.9 | 12.4 | 74.8 | 87.2 | 1.2 | 0.1 | 1.4 | 2.3 | 1.5 | 1.2 | 1.2 | 100.0 | 9.3 |
| SL_SiAT2 | 12.3 | 32.2 | 48.1 | 80.3 | 0.2 | 0.2 | 1.3 | 2.6 | 0.9 | 0.8 | 1.5 | 100.0 | 14.4 |
| FE_SiAT2 | 15.5 | 18.7 | 48.8 | 67.5 | 0.0 | 0.3 | 2.5 | 2.1 | 3.6 | 1.8 | 6.5 | 100.0 | 20.2 |
| FE_SiAT2ad | 36.1 | 7.1 | 30.0 | 37.1 | 0.1 | 0.6 | 6.6 | 5.7 | 10.2 | 1.8 | 2.0 | 100.0 | 30.1 |
| FE_SiAT3ad | 35.9 | 10.0 | 40.4 | 50.4 | 0.1 | 0.4 | 2.8 | 2.2 | 4.1 | 2.0 | 2.2 | 100.0 | 29.4 |
| FE_SAT2ad | 52.2 | 8.8 | 12.5 | 21.3 | 0.0 | 1.2 | 4.8 | 2.7 | 9.3 | 4.7 | 3.9 | 100.0 | 31.7 |
| FE_SAT3ad | 48.3 | 11.6 | 16.4 | 28.0 | 0.1 | 0.7 | 5.0 | 2.0 | 8.3 | 4.0 | 3.7 | 100.0 | 29.9 |
| SL_F1 * | 5.4 | 8.5 | 82.4 | 90.9 | 0.0 | 0.1 | 0.5 | 0.4 | 0.8 | 0.8 | 1.1 | 100.0 | 6.6 |
| SL_F2 * | 64.7 | 0.0 | 6.9 | 6.9 | 0.1 | 0.3 | 12.2 | 0.6 | 4.8 | 3.5 | 6.9 | 100.0 | 35.6 |
| SL_F3 * | 63.3 | 0.0 | 0.8 | 0.8 | 0.0 | 0.3 | 26.9 | 0.4 | 3.4 | 2.1 | 2.8 | 100.0 | 46.4 |
| Fucoidan Code | Fucoidan Size Calculated (kDa) | Fucoidan Size-Distribution Estimated (kDa) |
|---|---|---|
| LD_SiAT2 | 322 | 250–450 |
| SL_SiAT2 | 251 | 100–400 |
| FE_SiAT2 | 322 | 100–500 |
| FE_SiAT2ad | 366 | 200–500 |
| FE_SiAT3ad | 416 | 200–500 |
| FE_SAT2ad | 366 | 200–500 |
| FE_SAT3ad | 366 | 200–500 |
| SL_F1 * | Not determined | Not determined |
| SL_F2 * | >800 | 100–1000 |
| SL_F3 * | >800 | 100–1000 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dörschmann, P.; Mikkelsen, M.D.; Thi, T.N.; Roider, J.; Meyer, A.S.; Klettner, A. Effects of a Newly Developed Enzyme-Assisted Extraction Method on the Biological Activities of Fucoidans in Ocular Cells. Mar. Drugs 2020, 18, 282. https://doi.org/10.3390/md18060282
Dörschmann P, Mikkelsen MD, Thi TN, Roider J, Meyer AS, Klettner A. Effects of a Newly Developed Enzyme-Assisted Extraction Method on the Biological Activities of Fucoidans in Ocular Cells. Marine Drugs. 2020; 18(6):282. https://doi.org/10.3390/md18060282
Chicago/Turabian StyleDörschmann, Philipp, Maria Dalgaard Mikkelsen, Thuan Nguyen Thi, Johann Roider, Anne S. Meyer, and Alexa Klettner. 2020. "Effects of a Newly Developed Enzyme-Assisted Extraction Method on the Biological Activities of Fucoidans in Ocular Cells" Marine Drugs 18, no. 6: 282. https://doi.org/10.3390/md18060282