12-Deacetyl-12-epi-Scalaradial, a Scalarane Sesterterpenoid from a Marine Sponge Hippospongia sp., Induces HeLa Cells Apoptosis via MAPK/ERK Pathway and Modulates Nuclear Receptor Nur77

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. 12-Deacetyl-12-epi-scalaradial Inhibits HeLa Cell Proliferation
2.2. 12-Deacetyl-12-epi-scalaradial Induces HeLa Cells Apoptosis
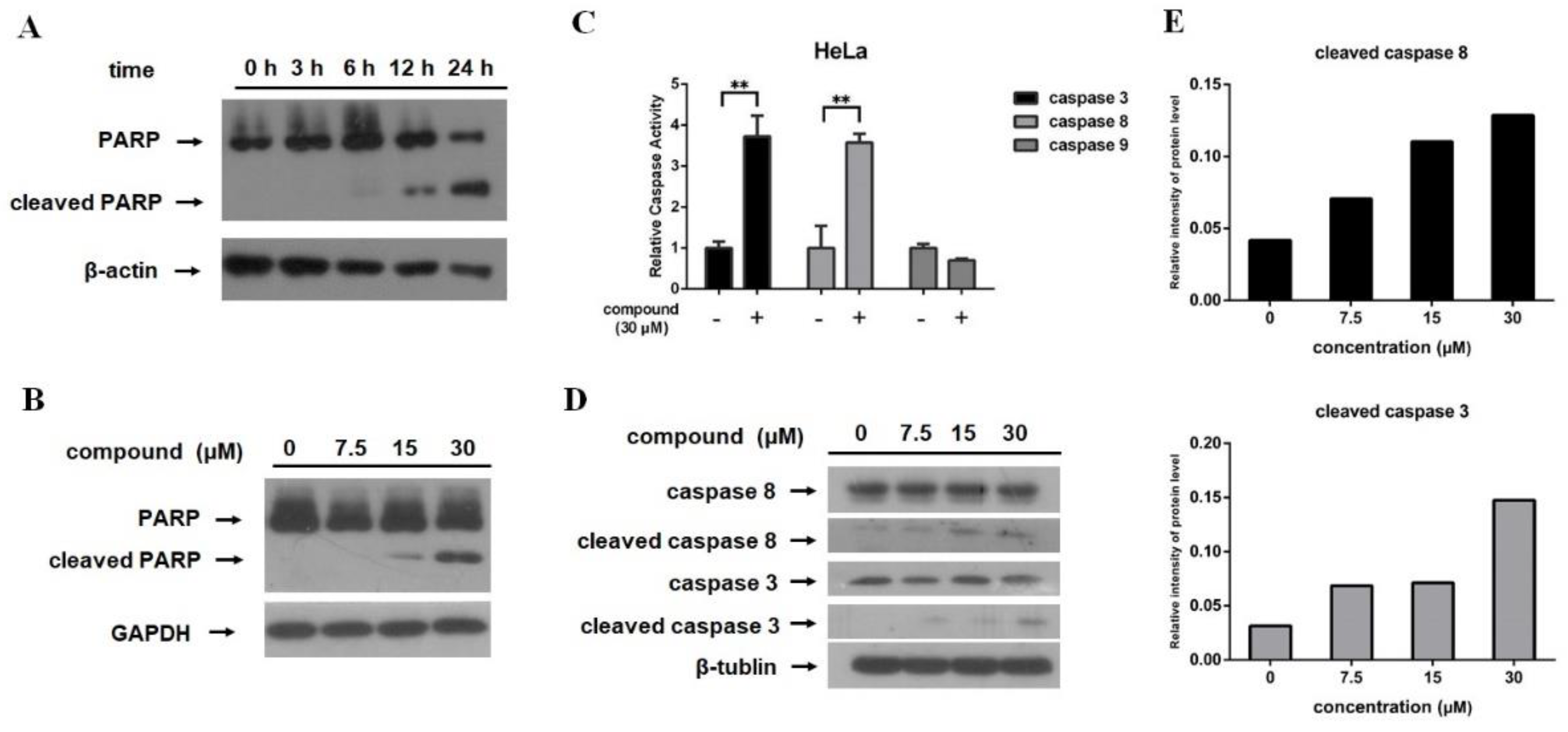
2.3. 12-Deacetyl-12-epi-scalaradial Induces PARP Cleavage and Activates Caspase Pathway in HeLa Cells
2.4. 12-Deacetyl-12-epi-scalaradial Suppresses MAPK/ERK Pathway
2.5. 12-Deacetyl-12-epi-scalaradial Modulates Trans-Activation Activity and Phosphorylation of Nur77, and Interacts with Nur77-LBD
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Isolation of Natural Products
4.3. Cell Cultures
4.4. Cytotoxic Activity
4.5. Flow Cytometry Analysis
4.6. Western Blot Analysis
4.7. Caspase Activation Activity
4.8. Dual-Luciferase Reporter Assay
4.9. Nur77-LBD Protein Purified
4.10. Fluorescence Quench Analysis
4.11. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Small, W.; Bacon, M.A.; Bajaj, A.; Chuang, L.T.; Fisher, B.J.; Harkenrider, M.M.; Jhingran, A.; Kitchener, H.C.; Mileshkin, L.R.; Viswanathan, A.N.; et al. Cervical cancer: A global health crisis. Cancer 2017, 123, 2404–2412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahrami, A.; Hasanzadeh, M.; Hassanian, S.M.; ShahidSales, S.; Ghayour-Mobarhan, M.; Ferns, G.A.; Avan, A. The potential value of the PI3K/Akt/mTOR signaling pathway for assessing prognosis in cervical cancer and as a target for therapy. J. Cell. Biochem. 2017, 118, 4163–4169. [Google Scholar] [CrossRef] [PubMed]
- Mohan, H.M.; Aherne, C.M.; Rogers, A.C.; Baird, A.W.; Winter, D.C.; Murphy, E.P. Molecular pathways: The role of NR4A orphan nuclear receptors in cancer. Clin. Cancer Res. 2012, 18, 3223–3228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moll, U.M.; Marchenko, N.; Zhang, X. p53 and Nur77/TR3—transcription factors that directly target mitochondria for cell death induction. Oncogene 2006, 25, 4725–4743. [Google Scholar] [CrossRef] [Green Version]
- Zhan, Y.; Du, X.; Chen, H.; Liu, J.; Zhao, B.; Huang, D.; Li, G.; Xu, Q.; Zhang, M.; Weimer, B.C.; et al. Cytosporone B is an agonist for nuclear orphan receptor Nur77. Nat. Chem. Biol. 2008, 4, 548–556. [Google Scholar] [CrossRef]
- Kolluri, S.K.; Bruey-Sedano, N.; Cao, X.; Lin, B.; Lin, F.; Han, Y.-H.; Dawson, M.I.; Zhang, X.-K. Mitogenic effect of orphan receptor TR3 and its Regulation by MEKK1 in lung cancer cells. Mol. Cell. Biol. 2003, 23, 8651–8667. [Google Scholar] [CrossRef] [Green Version]
- Li, H. Cytochrome c release and apoptosis induced by mitochondrial targeting of nuclear orphan receptor TR3. Science 2000, 289, 1159–1164. [Google Scholar] [CrossRef] [Green Version]
- Thigpen, T. The role of chemotherapy in the management of carcinoma of the cervix. Cancer J. 2003, 9, 425–432. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhao, W.; Xie, G.; Huang, M.; Hu, M.; Jiang, X.; Zeng, D.; Liu, J.; Zhou, H.; Chen, H.; et al. Induction of Nur77-dependent apoptotic pathway by a coumarin derivative through activation of JNK and p38 MAPK. Carcinogenesis 2014, 35, 2660–2669. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.H.; Park, J.-S.; Moon, B.; Kim, M.C.; Kim, J.-K.; Lee, S.; Suh, H.; Kim, N.D.; Kim, J.-M.; Park, Y.C.; et al. Orphan nuclear receptor Nur77 translocates to mitochondria in the early phase of apoptosis induced by synthetic chenodeoxycholic acid derivatives in human stomach cancer cell line SNU-1. Ann. N. Y. Acad. Sci. 2003, 1010, 171–177. [Google Scholar] [CrossRef]
- Tarhouni-Jabberi, S.; Zakraoui, O.; Ioannou, E.; Riahi-Chebbi, I.; Haoues, M.; Roussis, V.; Kharrat, R.; Essafi-Benkhadir, K. Mertensene, a halogenated monoterpene, induces G2/M cell cycle arrest and caspase dependent apoptosis of human colon adenocarcinoma HT29 cell line through the modulation of ERK-1/-2, AKT and NF-κB signaling. Mar. Drugs 2017, 15, 221. [Google Scholar] [CrossRef] [Green Version]
- Molinski, T.F.; Dalisay, D.S.; Lievens, S.L.; Saludes, J.P. Drug development from marine natural products. Nat. Rev. Drug Discov. 2009, 8, 69–85. [Google Scholar] [CrossRef]
- Roy, M.C.; Tanaka, J.; de Voogd, N.; Higa, T. New scalarane class sesterterpenes from an indonesian sponge, phyllospongia sp. J. Nat. Prod. 2002, 65, 1838–1842. [Google Scholar] [CrossRef]
- Chang, Y.-C.; Tseng, S.-W.; Liu, L.-L.; Chou, Y.; Ho, Y.-S.; Lu, M.-C.; Su, J.-H. Cytotoxic sesterterpenoids from a sponge hippospongia sp. Mar. Drugs 2012, 10, 987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hahn, D.; Won, D.H.; Mun, B.; Kim, H.; Han, C.; Wang, W.; Chun, T.; Park, S.; Yoon, D.; Choi, H.; et al. Cytotoxic scalarane sesterterpenes from a Korean marine sponge psammocinia sp. Bioorg. Med. Chem. Lett. 2013, 23, 2336–2339. [Google Scholar] [CrossRef] [PubMed]
- Renner, M.K.; Jensen, P.R.; Fenical, W. ChemInform abstract: Mangicols: Structures and biosynthesis of a new class of sesterterpene polyols from a marine fungus of the genus fusarium. ChemInform 2000, 31. [Google Scholar] [CrossRef]
- Cassiano, C.; Esposito, R.; Tosco, A.; Zampella, A.; D’Auria, M.V.; Riccio, R.; Casapullo, A.; Monti, M.C. Heteronemin, a marine sponge terpenoid, targets TDP-43, a key factor in several neurodegenerative disorders. Chem. Commun. 2014, 50, 406–408. [Google Scholar] [CrossRef]
- Aoki, S.; Higuchi, K.; Isozumi, N.; Matsui, K.; Miyamoto, Y.; Itoh, N.; Tanaka, K.; Kobayashi, M. Differentiation in chronic myelogenous leukemia cell K562 by spongean sesterterpene. Biochem. Biophys. Res. Commun. 2001, 282, 426–431. [Google Scholar] [CrossRef]
- Elhady, S.; El-Halawany, A.; Alahdal, A.; Hassanean, H.; Ahmed, S. A new bioactive metabolite isolated from the red sea marine sponge hyrtios erectus. Molecules 2016, 21, 82. [Google Scholar] [CrossRef] [Green Version]
- Danial, N.N.; Korsmeyer, S.J. Cell death: Critical control points. Cell 2004, 16, 205–219. [Google Scholar] [CrossRef] [Green Version]
- Fesik, S.W. Promoting apoptosis as a strategy for cancer drug discovery. Nat. Rev. Cancer. 2005, 5, 876–885. [Google Scholar] [CrossRef] [PubMed]
- Duriez, P.J.; Shah, G.M. Cleavage of poly (ADP-ribose) polymerase: A sensitive parameter to study cell death. Biochem. Cell Biol. 1997, 75, 337–349. [Google Scholar] [CrossRef] [PubMed]
- Oliver, F.J.; de la Rubia, G.; Rolli, V.; Ruiz-Ruiz, M.C.; de Murcia, G.; Murcia, J.M. Importance of Poly (ADP-ribose) polymerase and its cleavage in apoptosis. J. Biol. Chem. 1998, 273, 33533–33539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scovassi, C.S.A.I. Poly (adp-ribose) polymerase-1 cleavage during apoptosis. Apoptosis Int. J. Program. Cell Death. 2002, 7, 321. [Google Scholar] [CrossRef]
- Porter, A.G.; Jänicke, R.U. Emerging roles of caspase-3 in apoptosis. Cell Death Differ. 1999, 6, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Weinlich, R.; Oberst, A.; Beere, H.M.; Green, D.R. Necroptosis in development, inflammation and disease. Nat. Rev. Mol. Cell Biol. 2017, 18, 127–136. [Google Scholar] [CrossRef]
- Seger, R.; Wexler, S. The MAPK signaling cascades. Faseb J. 1995, 9, 726–735. [Google Scholar] [CrossRef]
- Fang, J.Y.; Richardson, B.C. The MAPK signalling pathways and colorectal cancer. Lancet Oncol. 2005, 6, 322–327. [Google Scholar] [CrossRef]
- Fresno, V.; Casado, E.; de Castro, J.; Cejas, P.; Belda, C.; Gonzalez, M. PI3K/Akt signalling pathway and cancer. Cancer Treat. Rev. 2004, 30, 193–204. [Google Scholar] [CrossRef]
- Wang, A.; Rud, J.; Olson, C.M.; Anguita, J.; Osborne, B.A. Phosphorylation of Nur77 by the MEK-ERK-RSK cascade induces mitochondrial translocation and apoptosis in T Cells. J. Immunol. 2009, 183, 3268–3277. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Nie, Y.; Li, Y.; Wan, Y.-J.Y. ERK1/2 deactivation enhances cytoplasmic Nur77 expression level and improves the apoptotic effect of fenretinide in human liver cancer cells. Biochem. Pharmacol. 2011, 81, 910–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, M.; Luo, Q.; Alitongbieke, G.; Chong, S.; Xu, C.; Xie, L.; Chen, X.; Zhang, D.; Zhou, Y.; Wang, Z.; et al. Celastrol-Induced Nur77 interaction with TRAF2 alleviates inflammation by promoting mitochondrial ubiquitination and autophagy. Mol. Cell. 2017, 66, 141–153. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zeng, H.; Zhang, L.; Zhan, Y.; Chen, Y.; Wang, Y.; Wang, J.; Xiang, S.; Liu, W.; Wang, W.; et al. A unique pharmacophore for activation of the nuclear orphan receptor Nur77 in vivo and in vitro. Cancer Res. 2010, 70, 3628–3637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, D.; Zhou, M.; Lin, T.; Chen, J.; Wang, G.; Huang, Y.; Jiang, X.; Tian, W.; Chen, H. Cytotoxic components from hypericum elodeoides targeting RXRα and inducing HeLa cell apoptosis through Caspase-8 activation and PARP cleavage. J. Nat. Prod. 2019, 82, 1072–1080. [Google Scholar] [CrossRef] [PubMed]
- Lakshmi, S.P.; Reddy, A.T.; Banno, A.; Reddy, R.C. Molecular, chemical, and structural characterization of prostaglandin A2 as a novel agonist for Nur77. Biochem. J. 2019, 496, 2757–2767. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.-J.; Qiu, Y.-Q.; Yao, X.-J.; Chen, H.-F.; Dai, Y.; Zhang, X.-K.; Yao, X.-S. Dioxasampsones A and B, two polycyclic polyprenylated acylphloroglucinols with unusual epoxy-ring-fused skeleton from Hypericum sampsonii. Org. Lett. 2014, 16, 6346–6349. [Google Scholar] [CrossRef]
- Tian, W.-J.; Yu, Y.; Yao, X.-J.; Chen, H.-F.; Dai, Y.; Zhang, X.-K.; Yao, X.-S. Norsampsones A–D, four new decarbonyl polycyclic polyprenylated acylphloroglucinols from Hypericum sampsonii. Org. Lett. 2014, 16, 3448–3451. [Google Scholar] [CrossRef]
- Qiu, D.; Zhou, M.; Chen, J.; Wang, G.; Lin, T.; Huang, Y.; Yu, F.; Ding, R.; Sun, C.; Tian, W.; et al. Hyperelodiones A-C, monoterpenoid polyprenylated acylphoroglucinols from Hypericum elodeoides, induce cancer cells apoptosis by targeting RXRα. Phytochemistry 2020, 170, 112216. [Google Scholar] [CrossRef]
- Wu, H.; Zeng, W.; Chen, L.; Yu, B.; Guo, Y.; Chen, G.; Liang, Z. Integrated multi-spectroscopic and molecular docking techniques to probe the interaction mechanism between maltase and 1-deoxynojirimycin, an α-glucosidase inhibitor. Int. J. Biol. Macromol. 2018, 114, 1194–1202. [Google Scholar] [CrossRef]
- Bozoğlan, B.K.; Tunç, S.; Duman, O. Investigation of neohesperidin dihydrochalcone binding to human serum albumin by spectroscopic methods. J. Lumin. 2014, 155, 198–204. [Google Scholar] [CrossRef]
- Ajmal, M.R.; Abdelhameed, A.S.; Alam, P.; Khan, R.H. Interaction of new kinase inhibitors cabozantinib and tofacitinib with human serum alpha-1 acid glycoprotein. A comprehensive spectroscopic and molecular Docking approach, spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2016, 159, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Chen, Z.; Hu, C.; Yan, S.; Li, Z.; Lian, B.; Xu, Y.; Ding, R.; Zeng, Z.; Zhang, X.; et al. Celastrol binds to its target protein via specific noncovalent interactions and reversible covalent bonds. Chem. Commun. 2018, 54, 12871–12874. [Google Scholar] [CrossRef] [PubMed]
- von Schwarzenberg, K.; Vollmar, A.M. Targeting apoptosis pathways by natural compounds in cancer: Marine compounds as lead structures and chemical tools for cancer therapy. Cancer Lett. 2013, 332, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, M.; Cerella, C.; Eifes, S.; Chateauvieux, S.; Morceau, F.; Jaspars, M.; Dicato, M.; Diederich, M. Heteronemin, a spongean sesterterpene, inhibits TNFα-induced NF-κB activation through proteasome inhibition and induces apoptotic cell death. Biochem. Pharmacol. 2010, 79, 610–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zahedifard, M.; Faraj, F.L.; Paydar, M.; Looi, C.Y.; Hajrezaei, M.; Hasanpourghadi, M.; Kamalidehghan, B.; Majid, N.A.; Ali, H.M.; Abdulla, M.A. Synthesis, characterization and apoptotic activity of quinazolinone Schiff base derivatives toward MCF-7 cells via intrinsic and extrinsic apoptosis pathways. Sci. Rep. 2015, 5, 11544. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S. Caspase function in programmed cell death. Cell Death Differ. 2007, 14, 32–43. [Google Scholar] [CrossRef]
- Bryant, K.L.; Stalnecker, C.A.; Zeitouni, D.; Klomp, J.E.; Peng, S.; Tikunov, A.P.; Gunda, V.; Pierobon, M.; Waters, A.M.; George, S.D.; et al. Combination of ERK and autophagy inhibition as a treatment approach for pancreatic cancer. Nat. Med. 2019, 25, 628–640. [Google Scholar] [CrossRef]
- Wagner, E.F.; Nebreda, Á.R. Signal integration by JNK and p38 MAPK pathways in cancer development. Nat. Rev. Cancer. 2009, 9, 537–549. [Google Scholar] [CrossRef]
- Janku, F.; Yap, T.A.; Meric-Bernstam, F. Targeting the PI3K pathway in cancer: Are we making headway? Nat. Rev. Clin. Oncol. 2018, 15, 273–291. [Google Scholar] [CrossRef]
- Fritsch, M.; Günther, S.D.; Schwarzer, R.; Albert, M.-C.; Schorn, F.; Werthenbach, J.P.; Schiffmann, L.M.; Stair, N.; Stocks, H.; Seeger, J.M.; et al. Caspase-8 is the molecular switch for apoptosis, necroptosis and pyroptosis. Nature 2019, 575, 683–687. [Google Scholar] [CrossRef]
- Chintharlapalli, S.; Burghardt, R.; Papineni, S.; Ramaiah, S.; Yoon, K.; Safe, S. Activation of Nur77 by selected 1,1-Bis(3′-indolyl)-1-(p-substituted phenyl )methanes induces apoptosis through nuclear pathways. J. Biol. Chem. 2005, 280, 24903–24914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, K.; Lee, S.-O.; Cho, S.-D.; Kim, K.; Khan, S.; Safe, S. Activation of nuclear TR3 (NR4A1) by a diindolylmethane analog induces apoptosis and proapoptotic genes in pancreatic cancer cells and tumors. Carcinogenesis 2011, 32, 836–842. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-J.; Kim, J.-Y.; Lee, S.J.; Kim, H.-J.; Oh, C.J.; Choi, Y.-K.; Lee, H.-J.; Do, J.-Y.; Kim, S.-Y.; Kwon, T.-K.; et al. α-Lipoic acid prevents neointimal Hyperplasia via induction of p38 mitogen-activated protein Kinase/Nur77-Mediated apoptosis of vascular smooth muscle cells and accelerates postinjury reendothelialization. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 2164–2172. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.-H.; Cao, X.; Lin, B.; Lin, F.; Kolluri, S.K.; Stebbins, J.; Reed, J.C.; Dawson, M.I.; Zhang, X.-K. Regulation of Nur77 nuclear export by c-Jun N-terminal kinase and Akt. Oncogene 2006, 25, 2974–2986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.-W.; Cobb, L.J.; Paharkova-Vatchkova, V.; Liu, B.; Milbrandt, J.; Cohen, P. Contribution of the orphan nuclear receptor Nur77 to the apoptotic action of IGFBP-3. Carcinogenesis 2007, 28, 1653–1658. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, M.; Peng, B.-R.; Tian, W.; Su, J.-H.; Wang, G.; Lin, T.; Zeng, D.; Sheu, J.-H.; Chen, H. 12-Deacetyl-12-epi-Scalaradial, a Scalarane Sesterterpenoid from a Marine Sponge Hippospongia sp., Induces HeLa Cells Apoptosis via MAPK/ERK Pathway and Modulates Nuclear Receptor Nur77. Mar. Drugs 2020, 18, 375. https://doi.org/10.3390/md18070375
Zhou M, Peng B-R, Tian W, Su J-H, Wang G, Lin T, Zeng D, Sheu J-H, Chen H. 12-Deacetyl-12-epi-Scalaradial, a Scalarane Sesterterpenoid from a Marine Sponge Hippospongia sp., Induces HeLa Cells Apoptosis via MAPK/ERK Pathway and Modulates Nuclear Receptor Nur77. Marine Drugs. 2020; 18(7):375. https://doi.org/10.3390/md18070375
Chicago/Turabian StyleZhou, Mi, Bo-Rong Peng, Wenjing Tian, Jui-Hsin Su, Guanghui Wang, Ting Lin, Dequan Zeng, Jyh-Horng Sheu, and Haifeng Chen. 2020. "12-Deacetyl-12-epi-Scalaradial, a Scalarane Sesterterpenoid from a Marine Sponge Hippospongia sp., Induces HeLa Cells Apoptosis via MAPK/ERK Pathway and Modulates Nuclear Receptor Nur77" Marine Drugs 18, no. 7: 375. https://doi.org/10.3390/md18070375