Sargassum fusiforme Polysaccharides Prevent High-Fat Diet-Induced Early Fasting Hypoglycemia and Regulate the Gut Microbiota Composition

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Preparation and Physicochemical Properties of SFPs
2.2. SFPs Significantly Alleviated HFD-Induced Early Fasting Hypoglycemia
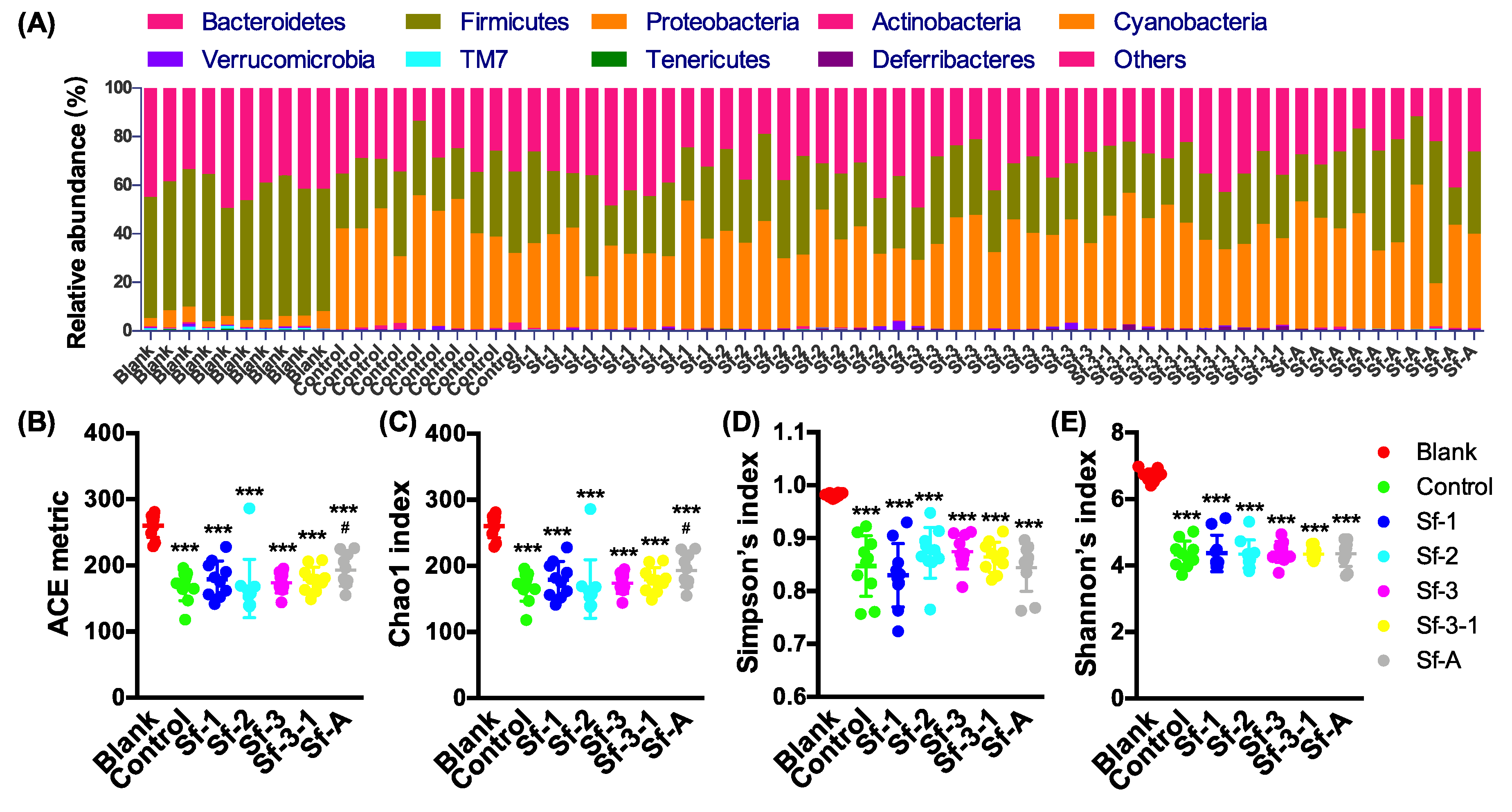
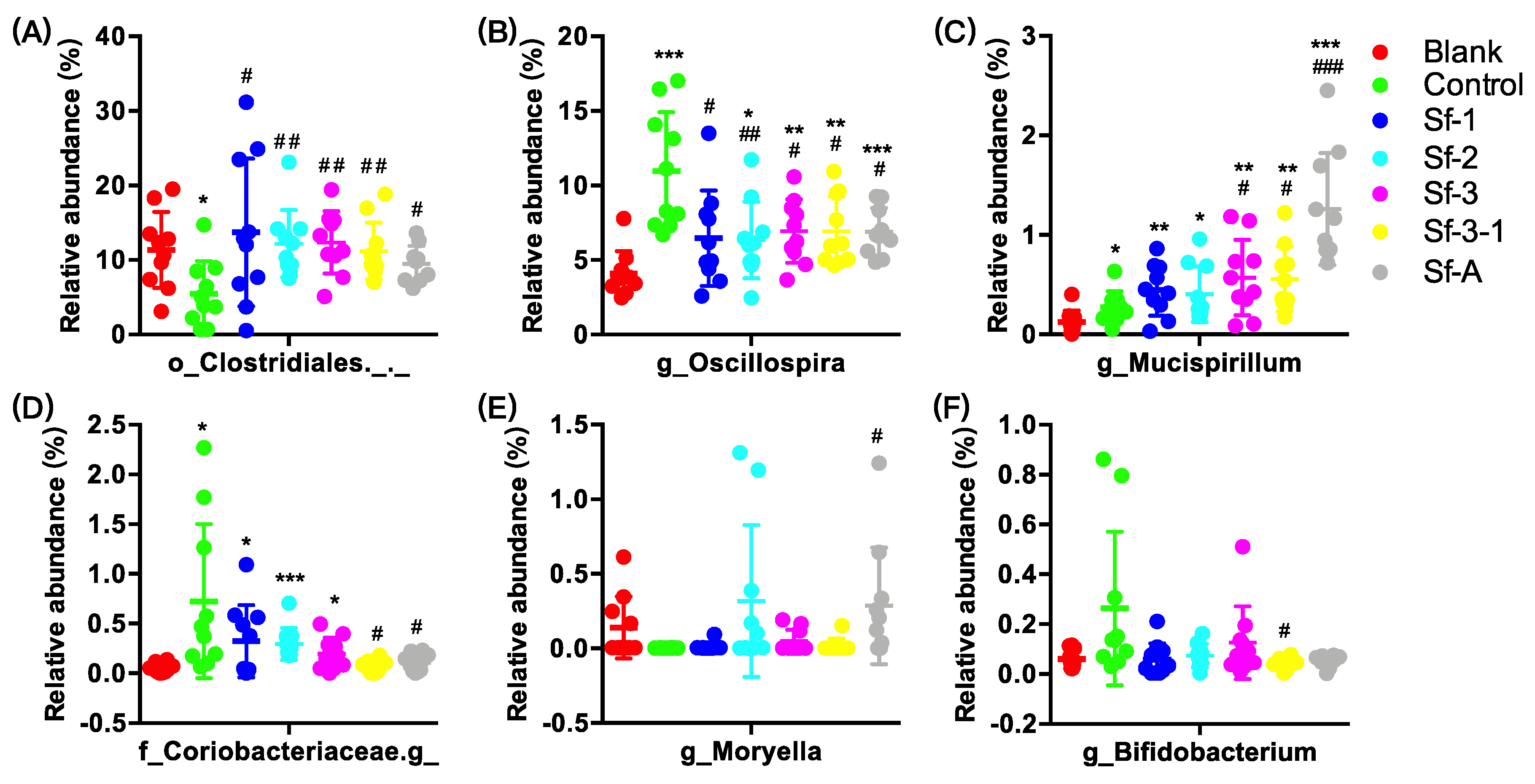
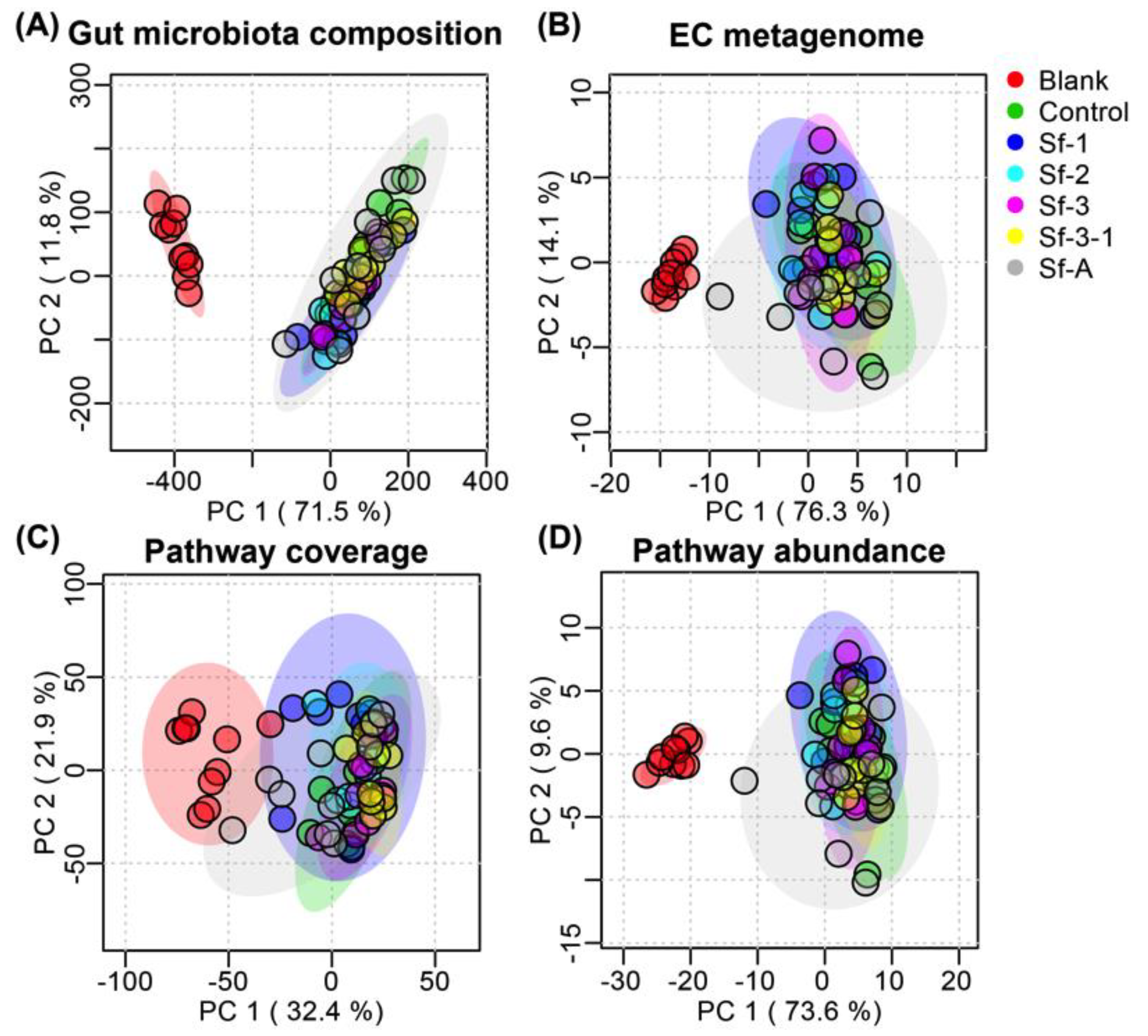
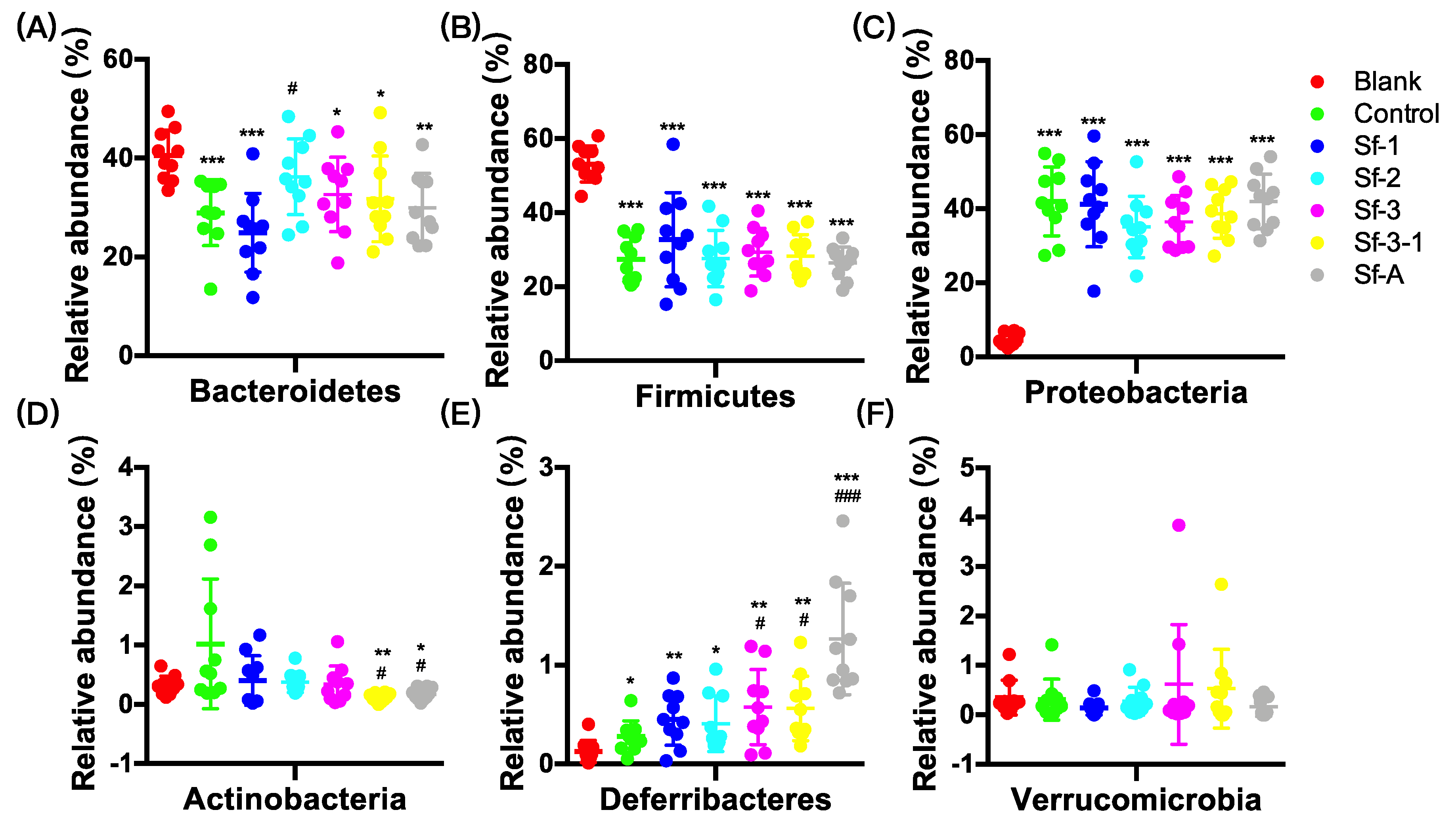
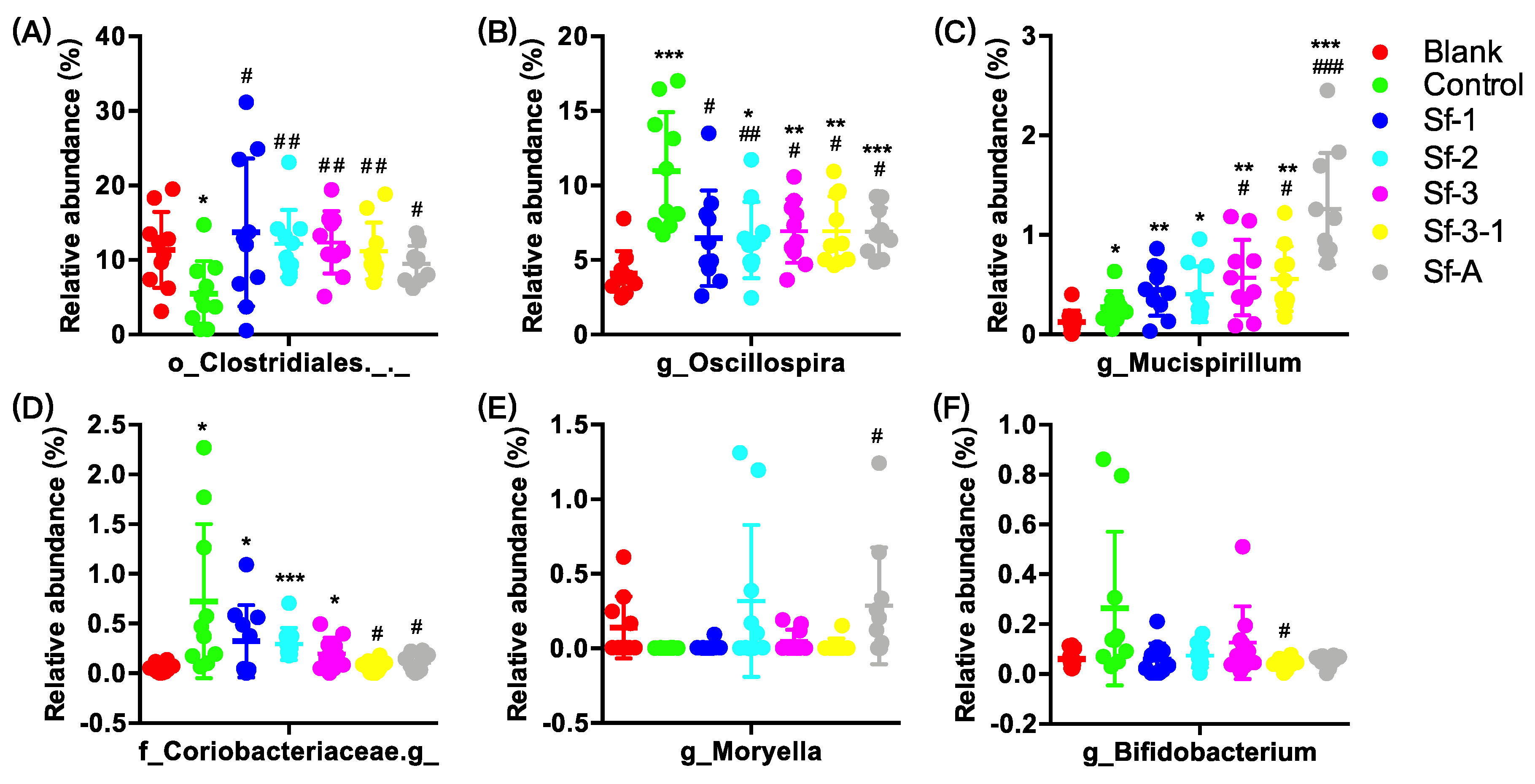
2.3. SFPs Regulated the Gut Microbiota Composition
2.4. SFPs Regulated the Metabolic Pathway Coverage of the Gut Microbiome
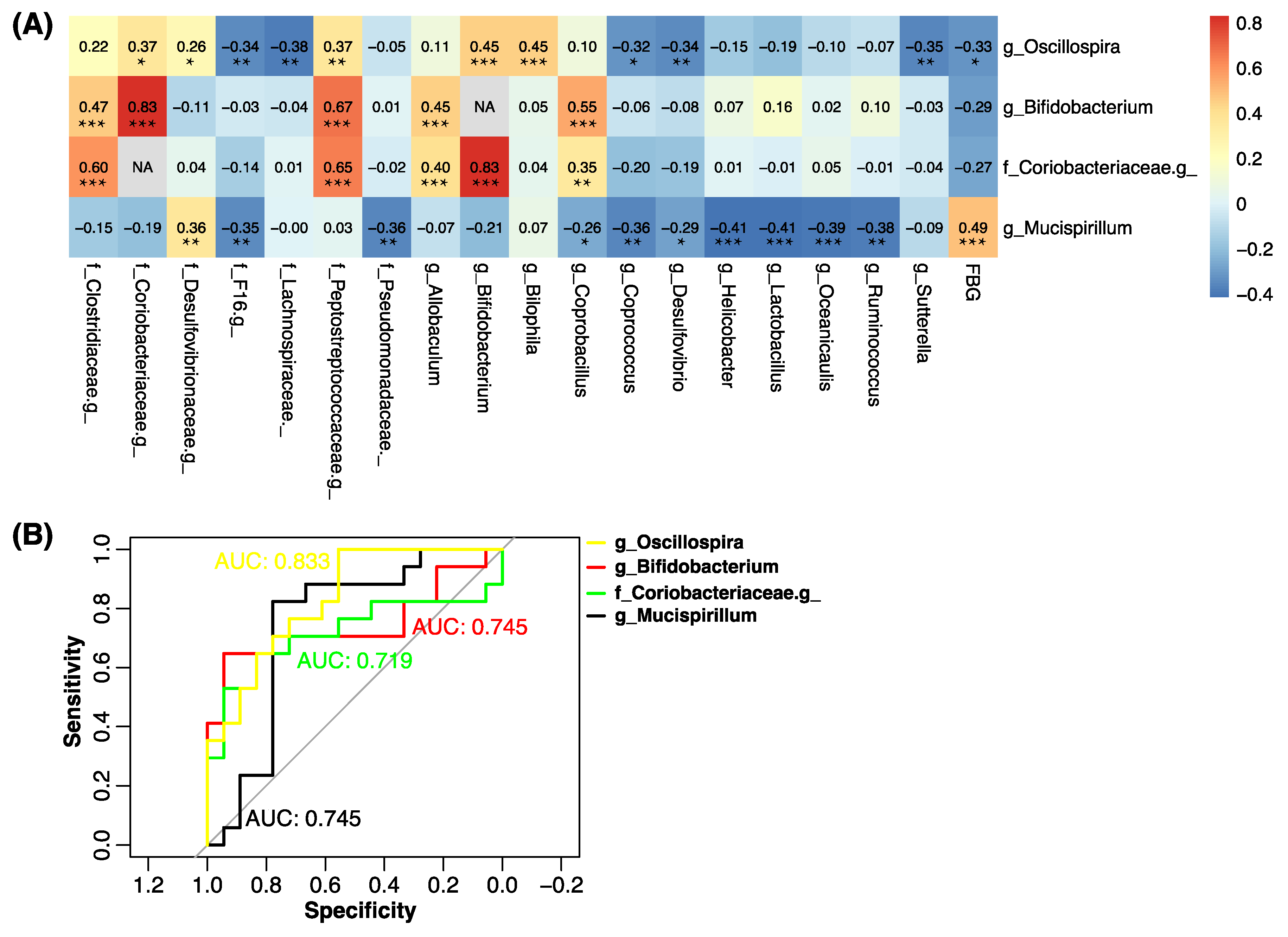
2.5. g_Mucispinllum and g_Oscillospira Were Associated with the Fasting Blood Glucose Level
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Preparation and Purification of SFPs
4.3. Chemical Analysis of SFPs
4.4. Animal Study
4.5. DNA Extraction and High Throughput Sequencing
4.6. Bioinformatics Analysis
4.7. Statistical Analysis
4.8. Accession Number
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Feng, F.J.; Yin, Z.H.; Wang, P.Y.; Zhang, Q.J.; Kang, W.Y. alpha-Glucosidase inhibitory compounds in malus Halliana koehne tea. Curr. Top. Nutraceutical Res. 2019, 17, 236–241. [Google Scholar]
- Chen, M.J.; Yan, X.; Chen, Y.Q.; Zhao, C. Phytochemicals for non-insulin diabetes mellitus: A minireview on plant-derived compounds hypoglycemic activity. J. Food Nutr. Sci. 2017, 5, 23–27. [Google Scholar] [CrossRef] [Green Version]
- Fan, S.J.; Guo, L.; Zhang, Y.; Sun, Q.H.; Yang, B.C.; Huang, C. Okra polysaccharide improves metabolic disorders in high-fat diet-induced obese C57BL/6 mice. Mol. Nutr. Food Res. 2013, 57, 2075–2078. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.W.; Qu, Y.H.; Zhang, H.; Xue, Z.J.; Liu, T.; Yang, L.L.; Sun, L.; Zhou, Y.F.; Fan, Y.Y. Hypoglycemic effects of polysaccharides from Gomphidiaceae rutilus fruiting bodies and their mechanisms. Food Funct. 2020, 11, 424–434. [Google Scholar] [CrossRef]
- Burchfield, J.G.; Kebede, M.A.; Meoli, C.C.; Stockli, J.; Whitworth, P.T.; Wright, A.L.; Hoffman, N.J.; Minard, A.Y.; Ma, X.; Krycer, J.R.; et al. High dietary fat and sucrose result in an extensive and time-dependent deterioration in health of multiple physiological systems in mice. J. Biol. Chem. 2018, 293, 5731–5745. [Google Scholar] [CrossRef] [Green Version]
- Hafizur, R.M.; Raza, S.A.; Chishti, S.; Shaukat, S.; Ahmed, A.A. ‘Humanized’ rat model of pre-diabetes by high fat diet-feeding to weaning wistar rats. Integr. Obes. Diabetes 2015, 1, 44–48. [Google Scholar]
- Liao, H.W.; Saver, J.; Yeh, H.C.; Chen, C.H.S.; Wu, Y.L.; Lee, M.; Ovbiagele, B. Low fasting glucose and future risks of major adverse outcomes in people without baseline diabetes or cardiovascular disease: A systematic review and meta-analysis. BMJ Open 2019, 9, 11. [Google Scholar] [CrossRef]
- He, X.Y.; He, J.J.; Zheng, N.N.; Wang, S.C.; Li, H.K. Study on the anti-obesity effect and modulation of gut microbiota by Astragalus polysaccharides in mice. World Chin. Med. 2016, 11, 2379–2384, 2388. [Google Scholar]
- Chen, P.C.; Zhang, Y.; Wu, M.J. How Sargassum fusiforme Polysaccharides Promote Health Condition. EC Nutr. 2018, 13, 63–64. [Google Scholar]
- Sun, G.W.; Lian, T.; Yang, B.; Gu, Y.; Li, X.J. Ameliorative effect of Sargassum fusiforme polysaccharides on oxidative stress and inflammation in ethanol-induced gastric ulcer. Pharm. Mag. 2019, 15, 244–252. [Google Scholar]
- Chen, P.; Yang, S.; Hu, C.; Zhao, Z.; Liu, J.; Cheng, Y.; Wang, S.; Chen, Q.; Yu, P.; Zhang, X.; et al. Sargassum fusiforme polysaccharide rejuvenates the small intestine in mice through altering its physiology and gut microbiota composition. Curr. Mol. Med. 2017, 17, 350–358. [Google Scholar] [PubMed]
- Cheng, Y.; Sibusiso, L.; Hou, L.F.; Jiang, H.J.; Chen, P.C.; Zhang, X.; Wu, M.; Tong, H. Sargassum fusiforme fucoidan modifies the gut microbiota during alleviation of streptozotocin-induced hyperglycemia in mice. Int. J. Biol. Macromol. 2019, 131, 1162–1170. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhang, X.X.; Tang, Y.X.; Mao, J.L. Composition, isolation, purification and biological activities of Sargassum fusiforme polysaccharides: A review. Carbohydr. Polym. 2020, 228. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Q.W.; Wei, B.; Wang, S.J.; Ke, S.Z.; Chen, J.W.; Zhang, H.W.; Wang, H. The antioxidant activity of polysaccharides derived from marine organisms: An overview. Mar. Drugs 2019, 17, 674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, S.Z.; Wei, B.; Qiu, W.H.; Zhou, T.S.; Wang, S.J.; Chen, J.W.; Chen, J.; Zhang, H.W.; Jin, W.H.; Wang, H. Structural characterization and α-glucosidase inhibitory and antioxidant activities of fucoidans extracted from Saccharina japonica. Chem. Biodivers. 2020. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, M.; Maison, N.; Achenbach, P.; Assfalg, R.; Braig, S.; Bocker, D.; Dunstheimer, D.; Ermer, U.; Gavazzeni, A.; Gerstl, E.M.; et al. Fasting hypoglycemia is associated with disease progression in presymptomatic early stage type 1 diabetes. Pediatric Diabetes 2018, 19, 1238–1242. [Google Scholar] [CrossRef]
- Giacco, R.; Parillo, M.; Rivellese, A.A.; Lasorella, G.; Giacco, A.; D’Episcopo, L.; Riccardi, G. Long-term dietary treatment with increased amounts at fiber-rich low-glycemic index natural foods improves blood glucose control and reduces the number of hypoglycemic events in type 1 diabetic patients. Diabetes Care 2000, 23, 1461–1466. [Google Scholar] [CrossRef] [Green Version]
- Soare, A.; Khazrai, Y.M.; Fontana, L.; Del Toro, R.; Lazzaro, M.C.; Di Rosa, C.; Buldo, A.; Fioriti, E.; Maddaloni, E.; Angeletti, S.; et al. Treatment of reactive hypoglycemia with the macrobiotic Ma-pi 2 diet as assessed by continuous glucose monitoring: The MAHYP randomized crossover trial. Metab. Clin. Exp. 2017, 69, 148–156. [Google Scholar] [CrossRef]
- Zhang, Z.W.; Xu, H.Y.; Zhao, H.; Geng, Y.; Ren, Y.L.; Guo, L.; Shi, J.; Xu, Z. Edgeworthia gardneri (Wall.) Meisn. water extract improves diabetes and modulates gut microbiota. J. Ethnopharmacol. 2019, 239. [Google Scholar] [CrossRef]
- Kuang, Y.S.; Lu, J.H.; Li, S.H.; Li, J.H.; Yuan, M.Y.; He, J.R.; Chen, N.N.; Xiao, W.Q.; Shen, S.Y.; Qiu, L.; et al. Connections between human gut microbiome and gestational diabetes mellitus. Gigascience 2017, 6. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Shen, D.Q.; Fang, Z.W.; Jie, Z.Y.; Qiu, X.M.; Zhang, C.F.; Chen, Y.; Ji, L. Human gut microbiota changes reveal the progression of glucose intolerance. PLoS ONE 2013, 8, e71108. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Zhao, X.; Guo, W.Y.; Gao, Y.; Yang, S.; Li, Z.; Wang, G.X. The relationship between frequently used glucose-lowering agents and gut microbiota in type 2 diabetes mellitus. J. Diabetes Res. 2018, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Wu, W.; Tang, H.; Wei, B.; Wang, H.; Sun, J.; Zhang, W.; Zhong, W. Structure analysis and anti-tumor and anti-angiogenic activities of sulfated galactofucan extracted from Sargassum thunbergii. Mar. Drugs 2019, 17, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, W.; Zhang, W.; Wang, J.; Ren, S.; Song, N.; Duan, D.; Zhang, Q. Characterization of laminaran and a highly sulfated polysaccharide from Sargassum fusiforme. Carbohydr. Polym. 2014, 385, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.M.; Pomeranz, Y. Effect of borate on colorimetric determinations of carbohydrates by the phenol-sulfuric acid method. Anal. Biochem. 1968, 24, 128–131. [Google Scholar] [CrossRef]
- Blumenkrantz, N.; Asboe-Hansen, G. New method for quantitative determination of uronic acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Dodgson, K.S.; Price, R.G. A note on determination of ester sulphate content of sulphated polysaccharides. Biochem. J. 1962, 84, 106–110. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.J.; Zhang, Q.B.; Wang, J.; Shi, X.L.; Zhang, Z.S. Analysis of the monosaccharide composition of fucoidan by precolumn derivation HPLC. Chin. J. Oceanol. Limn. 2009, 27, 578–582. [Google Scholar] [CrossRef]
- Jiang, S.; Du, P.G.; An, L.P.; Yuan, G.X.; Sun, Z.W. Anti-diabetic effect of Coptis Chinensis polysaccharide in high-fat diet with STZ-induced diabetic mice. Int. J. Biol. Macromol. 2013, 55, 118–122. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Caporaso, J.G. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Langille, M.G. PICRUSt2: An improved and extensible approach for metagenome inference. BioRxiv 2019, 672295. [Google Scholar] [CrossRef] [Green Version]
- Yan, Z.X.; Gao, X.J.; Li, T.; Wei, B.; Wang, P.P.; Yang, Y.; Yan, R. Fecal microbiota transplantation in experimental ulcerative colitis reveals associated gut microbial and host metabolic reprogramming. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [Green Version]
- Chong, J.; Wishart, D.S.; Xia, J. Using metaboanalyst 4.0 for comprehensive and integrative metabolomics data analysis. Curr. Protoc. Bioinform. 2019, 68. [Google Scholar] [CrossRef]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Total Sugar Content | Sulfate Content | Protein Content | Uronic Acid Content | Average Molecular Weight | Neutral Monosaccharide Composition (Molar Ratio) | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Man | Gal | Glc | GlcA | Fuc | ||||||
| Sf-1 | 80.30 ± 1.03 | - | 1.00 ± 0.43 | 0.17 ± 0.25 | 698.3/8.9 | - | - | 1 | - | - |
| Sf-2 | 63.73 ± 4.56 | 11.12 ± 0.68 | 1.03 ± 0.40 | 7.71 ± 1.56 | 95.5/9.5 | 0.26 | 0.23 | 0.21 | 0.30 | 1 |
| Sf-3 | 69.23 ± 2.51 | 35.08 ± 1.15 | 2.35 ± 0.27 | 2.34 ± 1.09 | 229.5 | - | 0.24 | - | - | 1 |
| Sf-3-1 | 51.95 ± 0.33 | 30.52 ± 1.58 | 0.96 ± 0.05 | 2.37 ± 0.66 | 10.0 | 0.09 | 0.33 | - | 0.08 | 1 |
| Sf-A | 63.60 ± 12.71 | 22.95 ± 1.77 | 0.47 ± 0.16 | 2.89 ± 0.35 | 46.5/5.1 | 0.10 | 0.42 | 1.26 | 0.05 | 1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, B.; Zhong, Q.-W.; Ke, S.-Z.; Zhou, T.-S.; Xu, Q.-L.; Wang, S.-J.; Chen, J.-W.; Zhang, H.-W.; Jin, W.-H.; Wang, H. Sargassum fusiforme Polysaccharides Prevent High-Fat Diet-Induced Early Fasting Hypoglycemia and Regulate the Gut Microbiota Composition. Mar. Drugs 2020, 18, 444. https://doi.org/10.3390/md18090444
Wei B, Zhong Q-W, Ke S-Z, Zhou T-S, Xu Q-L, Wang S-J, Chen J-W, Zhang H-W, Jin W-H, Wang H. Sargassum fusiforme Polysaccharides Prevent High-Fat Diet-Induced Early Fasting Hypoglycemia and Regulate the Gut Microbiota Composition. Marine Drugs. 2020; 18(9):444. https://doi.org/10.3390/md18090444
Chicago/Turabian StyleWei, Bin, Qi-Wu Zhong, Song-Ze Ke, Tao-Shun Zhou, Qiao-Li Xu, Si-Jia Wang, Jian-Wei Chen, Hua-Wei Zhang, Wei-Hua Jin, and Hong Wang. 2020. "Sargassum fusiforme Polysaccharides Prevent High-Fat Diet-Induced Early Fasting Hypoglycemia and Regulate the Gut Microbiota Composition" Marine Drugs 18, no. 9: 444. https://doi.org/10.3390/md18090444